




-
森林生物量是森林生态系统最基本的数量特征之一[1],不仅能反映森林结构特征和林木生产力,还在减小温室效应、稳定全球碳平衡与碳循环及固碳能力等方面具有不可替代的作用[2]。建立相应生物量模型以减少外业的工作量是目前较为认可的测定森林生物量的方法,不同的生物量模型对生物量的预估效果有极大影响,而模型间的不相容性也是亟待解决的问题。相关研究表明:通过联立估计建立的生物量相容性模型不仅能够解决独立模型之间的不相容问题,还具有参数稳定、预估精度较高的特点[3],对区域森林生物量的准确预估具有重要意义。闽楠Phoebe bournei为樟科Lauraceae楠属Phoebe的常绿大乔木,俗称楠木,是中国Ⅱ级重点保护植物。该树种树形优美,材质致密坚韧,纹理美观,削面光滑,不易反翘开裂,有隔音和驱蚊的效果,为高级家具、雕刻和建筑用材。此外,闽楠在景观设计、香料合成以及日常用品开发等方面都有重要的经济和生态价值。近些年对楠木木材的需求大增,导致各地盗伐现象频发[4],如今仅在福建、江西、浙江等海拔1 000 m以下的常绿阔叶林中有零星分布。为科学保护和培育珍贵楠木树种,国内不少学者对闽楠天然次生林开展研究,但主要集中在种群生态位[5]、种间联结与竞争[6]、种群结构与空间格局[7]以及自然更新[8]等方面,对次生林中闽楠生物量分配特征及相容性模型研究尚为缺乏。因此,本研究以江西省安福县闽楠天然次生林为研究对象,分析闽楠总生物量及各器官生物量分配特征,构建相容性生物量模型,旨在为准确预估研究区森林生物量提供理论参考。
-
安福县(27°04′~27°36′N,114°00′~114°47′E)位于江西省中部偏西。地形以山地、丘陵为主,三面环山;地势西北高,东南低;海拔300~1000 m。土壤类型主要为红壤、山地黄壤等。成土母岩类型主要为千枚岩、花岗岩、板岩等。属亚热带季风温润性气候区,年平均气温为17.7 ℃,最热月为7月,平均气温28.9 ℃,最冷月1月,平均气温5.9 ℃。年均降水量1 553.0 mm,平均降雨日166.0 d,降水明显集中在春季和初夏,平均日照时数1 649.0 h,山区日照偏少,年无霜期279.0 d,适宜林木生长。全县森林覆盖率达70.5%,被誉为“樟乡安福”,森林资源丰富。森林植被类型有阔叶林、针阔混交林、针叶林、竹林等,主要树种有马尾松Pinus massoniana,杉木Cunninghamia lanceolata,樟树Cinnamomum camphora,木荷Schima superba,枫香Liquidambar formosana,拟赤杨Alniphyllum fortunei和毛竹Phyllostachys edulis等[9]。灌木草本主要以檵木Loropetalum chinense,杜茎山Maesa japonica,铁芒萁Dicranopterias linearis和庐山楼梯草Elatostema stewardii等为主[6]。
-
通过森林资源二类调查数据及在当地林业部门获取的信息,在对安福县闽楠天然次生林分布地进行踏查的基础上,选择人为干扰程度轻且有代表性的地块设置标准地,面积为400 m2(20 m × 20 m)或600 m2(20 m × 30 m),共计16块标准地。记录标准地所有乔木树种、胸径、树高等因子,同时,选择闽楠对象木,对其生物量及周围的竞争木进行调查。
-
由于闽楠为Ⅱ级重点保护植物,对象木的采伐受到严格限制,因此对闽楠研究过程中未按常规要求采伐大量对象木。根据标准地每木检尺数据,选取1~2株与标准地林分平均胸径及平均树高相接近且生长正常、无病虫害的林木作为平均标准木用于生物量研究。总共选取了17株闽楠,其胸径为12.3~32.1 cm,树高为12.7~23.4 m。采用整株收获法,为准确获取树冠生物量,按树冠长度平均分为上层和下层,记录各层枝条数量,并在各层分别取1~2根与该层平均枝基径与平均枝长相近且生长正常的标准枝,17株解析木共获取35根标准枝,在标准枝基部测量直径,剖离枝条,测量枝条长度和宽幅,并分不同层次(上层、下层),不同枝级(一级枝、次级枝)和不同种类(枝、叶)称取鲜质量。将各层标准枝的树枝和叶各取500 g左右的样品,准确称取鲜质量并带回。称取树干解析后的所有圆盘和分段后的树干鲜质量。挖掘根系时尽可能将根系全部挖出,对根系称量前清除干净泥土等附着物。将所有的样品带回实验室烘干、称量,计算各类样品含水率进而计算单木各器官和全株干质量。
-
生物量模型中的自变量通常选择实际易测量且与各器官生物量相关性较强的林木因子,且不宜选择过多自变量。目前研究学者多采用胸径(D),树高(H)或D2H,DH等作为模型自变量。本研究参考相关文献[10]并通过各分项生物量与林木因子之间的相关关系分析,将D,H和D2H及反映树冠长度及其所占空间大小的因子冠幅(WC)和树冠体积(VC)作为待选自变量。由于林木竞争值(IC)对生物量分配的影响也相对较大,为提高模型的预估效果,也将IC作为模型的一个待选自变量。
-
选取与各组分生物量相关性显著的因子作为模型的自变量,采用非线性方程建立各组分生物量独立模型。为消除异方差性,将方程进行对数转换,利用最小二乘法估计回归系数[11]。
-
参照唐守正等[12]提出的思路,本研究以总生物量为基础,采用3级控制的方法对生物量进行相容性模型的建立。表达式详见参考文献[12]。生物量数据通常表现出异方差性,即误差的方差随观测值是变化的,因此需选择权函数消除异方差。相容性模型构建中,采用了对数转换的方法消除独立模型生物量的异方差,而由于相容性生物量模型方程式较复杂,采用对数转换比较复杂,因此本研究采用非线性加权法,根据独立模型的残差方差来确定权函数,即地上、地下、树冠、树枝、树叶和树干生物量独立拟合方程的方差建立的一元回归方程(W=1/Dx,其中D为样木胸径),方程参数是通过对各独立模型的方差进行拟合而得,在采用ForStat 2.1软件求解参数时,采取每一个方程两边乘以权重变量的方法进行处理[13]。
-
为分析天然林中闽楠所受周围林木的竞争压力,采用研究林木种群竞争关系及单木生长模型常用的与距离有关的竞争指数Hegyi指数[14]。惠刚盈等[15]研究表明:以参照数及其相邻最近4株树组成的最小空间结构单元更能恰当地进行林分空间结构分析。因此,在用邻近木法确定对象木周围的竞争木时,取离对象木最近的4株树来计算,测量竞争木胸径、树高、冠幅及其到相应对象木的距离。
-
模型构建中,需要有一套指标来进行检验与评价。常用的模型评价指标有决定系数(R2),均方根误差(ERMS),剩余标准差(ESE),赤池信息准则(CAI),总相对误差(ETR),平均系统误差(EMS),平均百分标准误差(EMPS)和预估精度(P),具体定义及表达式见文献[12, 16]。
-
运用SPSS 19.0的简单相关分析法分析各组分生物量与林木特征因子的相关关系,采用单因素方差分析法分析不同直径枝条或胸径下各组分生物量大小及其占比的差异。运用Forsat 2.1软件的非线性度量误差法建立相容性生物量模型。运用Excel进行数据处理分析。运用Origin 7.5进行相关图表绘制。
-
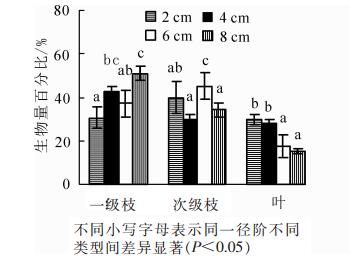
枝条生物量为0.343~16.474 kg·枝-1,其中一级枝生物量为0.087~9.308 kg·枝-1,次级枝生物量为0.110~5.299 kg·枝-1,叶生物量为0.096~2.373 kg·枝-1。由图 1可知:叶生物量占枝条生物量的比值随枝径阶的增加而不断减少,而一级枝和次级枝生物量占比随枝径阶增加呈波动变化,其中,径阶为4和8 cm时一级枝生物量占比最大,而径阶为2和6 cm时次级枝生物量占比最大。单因素方差分析表明:不同枝径阶的一级枝、次级枝和叶生物量分配比均差异显著(P<0.05)。

图 1 不同枝径阶枝、叶生物量分配比
Figure 1. Biomass allocation ratios of branch and leaf at different branch diameters levels
-
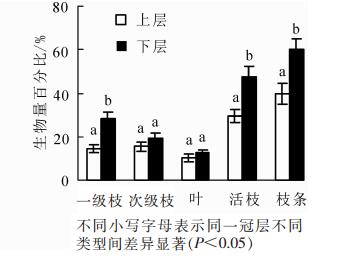
树冠生物量为23.40~185.71 kg·枝-1。由表 1可知:整体上来看,树冠一级枝和叶总生物量均随胸径的增加而不断增加,次级枝生物量则随胸径的增加呈先减后增的变化趋势;同一径阶各部分生物量表现为一级枝>次级枝>叶。不同径阶的一级枝、次级枝和叶生物量占整株生物量的比值差异显著(P<0.05),但进一步分析表明,当除去径阶为30 cm的各部分生物量,其余不同胸径间的一级枝生物量以及树冠总生物量之间均差异不显著(P>0.05)。树冠的垂直分配特征如图 2所示。下层一级枝和次级枝之和(活枝)明显大于上层。方差分析表明:位于树冠上层一级枝、次级枝及叶生物量之和(枝条)显著低于下层枝条生物量(P<0.05);位于树冠上层的活枝生物量显著低于下层生物量(P<0.05),其中一级枝生物量显著低于下层(P<0.05),而次级枝和叶生物量无显著差异(P>0.05)。
表 1 不同胸径阶树冠生物量分配特征
Table 1. Biomass allocation ratios of tree crown at different DBH levels
径阶/cm 生物量/(kg·株-1) 一级枝 次级枝 叶 总 14 15.75 ± 5.59 a 14.04 ± 3.80 ab 7.23 ± 3.74 a 33.51 ± 13.12 a 18 16.19 ± 3.20 a 11.49 ± 2.58 a 10.83 ± 2.50 a 38.51 ± 8.10 a 22 30.41 ± 6.53 a 31.60 ± 6.09 b 16.56 ± 1.68 ab 78.56 ± 13.86 a 26 39.13 ± 12.20 a 22.31 ± 1.40 ab 17.09 ± 2.05 ab 82.73 ± 11.73 a >30 79.57 ± 6.30 b 50.34 ± 2.36 c 25.36 ± 1.24 b 155.28 ± 7.40 b 说明:同列不同字母表示同一指标不同径阶间差异显著(P<0.05) 
图 2 不同冠层各组分生物量分配比
Figure 2. Biomass allocation ratio of each component at different crown levels
-
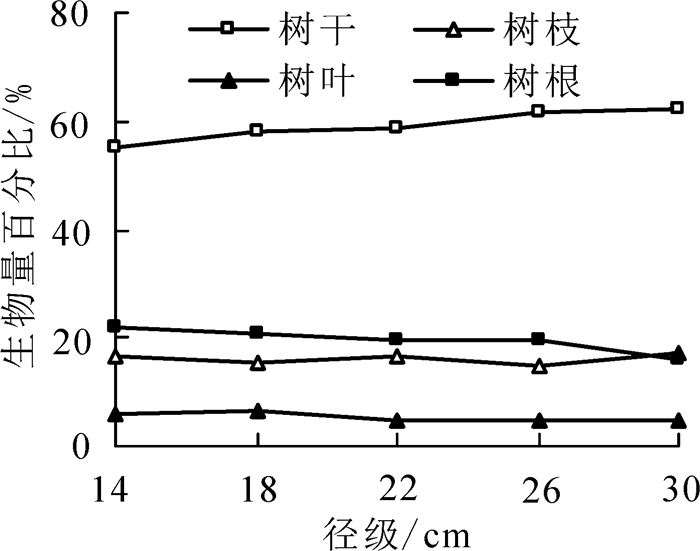
单木生物量为110.92~725.93 kg·株-1,各器官生物量占整株生物量的比值表现为:0.59(树干)>0.19(树根)>0.16(树枝)>0.06(树叶)。由表 2可知:各器官生物量均随胸径的增加而增加,不同胸径大小的各器官生物量差异显著(P<0.05)。由图 3可知:随着胸径的增大,树枝和树叶生物量占比表现为波动变化,树干生物量占比呈现逐渐增大的趋势,树根生物量占比则表现为下降趋势。不同胸径级的树根生物量占比差异显著(P<0.05),其余各器官生物量占比均差异不显著(P>0.05)。
表 2 不同胸径阶各器官生物量特征
Table 2. Biomass allocation of each tree organ at different DBH levels
径阶/cm 株数 生物量/(kg·株-1) 树干 树枝 树叶 地上部分 树根 全株 14 2 70.42 ± 10.82 a 21.01 ± 1.77 a 7.42 ± 1.05 a 98.84 ± 0.54 a 27.64 ± 11.55 a 126.49 ± 11.01 a 18 5 108.91 ± 15.73 ab 27.24 ± 2.25 a 11.68 ± 1.59 ab 147.83 ± 2.78 a 37.55 ± 15.80 ab 185.38 ± 18.31 a 22 4 161.24 ± 12.87 b 45.40 ± 3.20 b 12.59 ± 0.70 ab 219.22 ± 3.18 b 54.23 ± 13.47 b 273.45 ± 15.71 b 26 3 233.13 ± 14.08 c 54.83 ± 5.53 b 17.09 ± 2.05 b 305.05 ± 0.62 c 72.75 ± 7.78 c 377.80 ± 8.36 c >30 3 407.64 ± 24.18 d 131.44 ± 6.81 c 32.18 ± 4.39 c 571.26 ± 9.71 d 97.42 ± 16.09 d 668.68 ± 24.73 d 均值 191.33 ± 28.14 54.04 ± 9.31 15.97 ± 2.17 261.33 ± 74.21 57.09 ± 6.03 318.42 ± 44.32 说明:同列不同字母表示同一器官不同径阶间差异显著(P<0.05) 
图 3 不同径级各器官生物量分配比
Figure 3. Biomass allocation ratio of each tree organ at different DBH levels
-
由表 3可知:各器官生物量与林木特征因子之间存在显著相关,其中树干生物量与林木年龄、冠长呈显著相关(P<0.05),与树高极显著相关(P<0.01),树根生物量与树高显著相关(P<0.05),树干、树枝、树叶、树根生物量与胸径、冠幅和树冠体积均呈极显著相关(P<0.01)。树干生物量与胸径有较强相关性,树枝和树冠生物量则与冠幅和树冠体积相关性较大。
表 3 林木特征因子与各器官或各组分生物量之间的相关系数
Table 3. Correlation coefficient between tree characteristics and biomass of various organs or components
特征因子 树干 树枝 树叶 树根 树冠 地上部分 全株 年龄 0.499* 0.341 0.140 0.379 0.307 0.425 0.422 胸径 0.946** 0.886** 0.785** 0.930** 0.878** 0.947** 0.951** 树高 0.620** 0.404 0.326 0.582* 0.395 0.568* 0.574* 冠幅 0.761** 0.844** 0.689** 0.714** 0.825** 0.797** 0.791** 树冠体积 0.829** 0.862** 0.764** 0.767** 0.855** 0.855** 0.849** 冠长 0.494* 0.332 0.352 0.473 0.340 0.460 0.465 说明:*表示显著相关,**表示极显著相关 -
将选取的D,H,D2H,WC,VC和IC逐步代入到回归方程进行参数拟合,最后根据调整的决定系数(R2),ESE,CAI及模型复杂程度等选择较优的独立基础模型。由于各种因子组成的回归方程形式较多,本研究仅列出各组分拟合效果较好的前3个模型。较优模型F值检验结果均表现为极显著(P<0.001),独立基础模型最终形式、参数估计值及拟合优度见表 4。由表 4可知:树冠、树枝和树叶生物量模型R2较小,其余各项生物量模型的R2均达93.0%以上,其中树干生物量模型的R2达96.3%,树叶生物量模型最低,最优R2为71.9%。绝大部分基于D的独立模型R2都较高;模型在有D的基础上逐步加入D2H,IC,WC变量后,R2均有一定程度的提高。在模型中引入IC因子能在一定程度上提高各分项生物量的预估效果,而WC仅对树冠生物量预估有一定影响。除了自变量本身与生物量相关性从而影响模型R2大小外,自变量个数的增加也会导致模型R2增大,因此利用调整决定系数(RAdj2)值来评价模型的优劣。综合考虑RAdj2较大者、ESE较小者以及CAI较小者为最优模型,最终模型形式及评价指标见表 5。由表 5可知:模型对闽楠全株及各项生物量估计的ETR均在10%以内,EMS在5%以内。所有模型的EMPS在30%以内,其中树冠、树枝、树叶的EMPS较大。除树叶、树枝、树冠生物量预估精度较小以外,其余各项生物量预估精度均大于90%,其中模型对树干生物量预估精度达93%。综上,最终所选模型对生物量拟合效果相对较好,可作为相容性模型的基础模型。
表 4 总量(W)与各分量独立模型的参数估计值和拟合优度
Table 4. Measurement error and fitting optimization of independent models
项目 模型形式 拟合优度 R2 RAdj2 ESE CAI lnW=r0+r1 ln(D2H) 0.963 0.960 21.011 12.27 树干 lnW=r0+r1 lnD+r2 ln(D2H) 0.964 0.959 23.076 12.83 lnW=r0+r1 lnD+r2 ln(D2H)+r3 lnIC 0.965 0.957 22.147 13.56 lnW=r0+r1 lnD 0.830 0.818 17.532 -0.34 树枝 lnW=r0+r1 lnD+r4 lnWC 0.852 0.831 15.803 -0.37 lnW=r0+r1 lnD+r3 lnIC+r4WC 0.858 0.825 19.124 -0.14 lnW=r0+r1 lnD 0.595 0.568 9.248 -3.23 树叶 lnW=r0+r1 lnD+r3 lnIC 0.718 0.677 5.361 -3.49 lnW=r0+r1 lnD+r3 lnIC+r4WC 0.719 0.654 5.312 -3.20 lnW=r0+r1 lnD 0.807 0.795 20.124 -0.64 树冠 lnW=r0+r1 lnD+r3 lnIC 0.815 0.788 19.452 -0.14 lnW=r0+r1 lnD+r3 lnIC+r4WC 0.833 0.795 19.024 -0.72 地上部分 lnW=r0+r1 lnD 0.937 0.933 38.243 9.28 lnW=r0+r1 ln(D2H) 0.946 0.943 37.156 9.69 lnW=r0+r1 lnD+r2 ln(D2H) 0.954 0.948 37.425 9.18 lnW=r0+r1 lnD 0.871 0.863 9.246 7.41 树根 lnW=r0+r1 ln(D2H) 0.876 0.868 8.725 6.34 lnW=r0+r1 lnD+r2 ln(D2H)+r3 lnIC 0.885 0.859 8.829 6.29 lnW=r0+r1 lnD 0.935 0.931 42.576 9.73 全株 lnW=r0+r1 lnD+r2 ln(D2H) 0.952 0.945 41.086 9.57 lnW=r0+r1 lnD+r2 ln(D2H)+r3 lnIC 0.953 0.941 40.536 11.22 表 5 总量(W)与各分量最优独立模型形式与参数估计值
Table 5. Model form of optimal independent model
项目 模型形式 参数估计值 评价指标 a b c d e ETR EMS EMPS P 树干 lnW=r0+r1 ln(D2H) 5.366 0.936 1.203 0.653 9.429 93.1 树枝 lnW=r0+r1 lnD+r4 lnWC -2.639 1.751 0.560 5.106 3.069 21.866 80.8 树叶 lnW=r0+r1 lnD+r3 lnIC 1.253 0.488 -0.649 4.097 3.584 21.834 78.2 树冠 lnW=r0+r1 lnD+r3 lnIC+r4WC -0.368 1.143 -0.301 0.521 4.188 2.765 21.161 82.4 地上部分 lnW=r0+r1 lnD+r2 ln(D2H) 2.853 0.886 0.540 1.601 0.664 10.293 90.9 树根 lnW=r0+r1 lnD+r2 ln(D2H)+r3 lnIC 1.889 0.710 0.356 0.005 1.357 1.124 11.936 90.2 全株 lnW=r0+r1 lnD+r2 ln(D2H) 3.124 0.863 0.500 1.646 0.848 9.901 91.8 说明:各分量权重函数树干1/D2.329,树枝1/D2.203,树叶1/D1.252,树冠1/D2.996,地上部分1/D3.156,树根1/D2.007;各权函数F检验均显著(P<0.05) -
利用上述最优独立基础模型基于非线性度量误差法构建相容性联立方程组。一级控制:
$$ {{\tilde W}_2} = \frac{1}{{1 + \frac{{{a_3}}}{{{a_2}}}{D^{{b_3} - {b_2}}}{{\left( {{D^2}H} \right)}^{{c_5} - {c_2}}}{I_{\rm{C}}}^{{d_2}}}} \times {{\mathord{\buildrel{\lower3pt\hbox{$\scriptscriptstyle\frown$}} \over W} }_1}; $$ $$ {{\tilde W}_3} = \frac{1}{{1 + \frac{{{a_2}}}{{{a_3}}}{D^{{b_2} - {b_3}}}{{\left( {{D^2}H} \right)}^{{c_2} - {c_3}}}{I_{\rm{C}}}^{{d_2}}}} \times {{\mathord{\buildrel{\lower3pt\hbox{$\scriptscriptstyle\frown$}} \over W} }_1}。 $$ 其中:${\mathord{\buildrel{\lower3pt\hbox{$\scriptscriptstyle\frown$}} \over W} _1} $为总生物量估计值,$ \tilde{W}_{2}$和$\tilde{W}_{3} $为联合估计后的地上部分生物量和树根生物量估计值。令$ {a_{\rm{3}}}{\rm{ / }}{a_{\rm{2}}}{\rm{ = }}{r_0}, \quad {b_3} - {b_2} = {r_1}, \quad {c_3} - {c_2} = {r_2}, \quad {d_2} = {r_3};\quad {r_0}, \quad {r_1}, \quad {r_2}, \quad {r_3}$为联合估计参数,其初值为地上部分和树根生物量独立模型的参数估计值。二级控制:
$$ {{\tilde W}_4} = \frac{1}{{1 + \frac{{{a_7}}}{{{a_4}}}{D^{{b_7} - {b_4}}}{{\left( {{D^2}H} \right)}^{{c_7} - {c_4}}}{I_{\rm{C}}}^{{d_7} - {d_4}}{W_{\rm{C}}}^{{e_7} - {e_4}}}} \times {{\mathord{\buildrel{\lower3pt\hbox{$\scriptscriptstyle\frown$}} \over W} }_2}; $$ $$ {{\tilde W}_7} = \frac{1}{{1 + \frac{{{a_4}}}{{{a_7}}}{D^{{b_4} - {b_7}}}{{\left( {{D^2}H} \right)}^{{c_4} - {c_7}}}{I_{\rm{C}}}^{{d_4} - {d_7}}{W_{\rm{C}}}^{{e_4} - {e_7}}}} \times {{\mathord{\buildrel{\lower3pt\hbox{$\scriptscriptstyle\frown$}} \over W} }_2}。 $$ 其中:${{\mathord{\buildrel{\lower3pt\hbox{$\scriptscriptstyle\frown$}} \over W} }_2} $为地上部分生物量估计值,$\tilde{W}_{4} $和$ \tilde{W}_{7}$为联合估计后的树干和树冠生物量估计值。令$a_{7} / a_{4}=r_{0} $,$ {b_7} - {b_4} = {r_1}, \quad {c_7} - {c_4} = {r_2}, \quad {d_7} - {d_4} = {r_3}, \quad {e_7} - {e_4} = {r_4}。\quad {r_0}, \quad {r_1}, \quad {r_2}, \quad {r_3}$和r4为联合估计参数,其初值为树干和树冠生物量独立模型的参数估计值。三级控制:
$$ {{\tilde W}_5} = \frac{1}{{1 + \frac{{{a_6}}}{{{a_5}}}{D^{{b_6} - {b_5}}}{I_{\rm{C}}}^{{d_6}}{W_{\rm{C}}}^{ - {e_5}}}} \times {{\mathord{\buildrel{\lower3pt\hbox{$\scriptscriptstyle\frown$}} \over W} }_7}; $$ $$ {{\tilde W}_6} = \frac{1}{{1 + \frac{{{a_5}}}{{{a_6}}}{D^{{b_5} - {b_6}}}{I_{\rm{C}}}^{ - {d_6}}{W_{\rm{C}}}^{{e_5}}}} \times {{\mathord{\buildrel{\lower3pt\hbox{$\scriptscriptstyle\frown$}} \over W} }_7}。 $$ 其中:${{\mathord{\buildrel{\lower3pt\hbox{$\scriptscriptstyle\frown$}} \over W} }_7} $为树冠生物量估计值,$ \tilde{W}_{5}$和$ \tilde{W}_{6}$为联合估计后的树枝和树叶生物量的估计值。令$ a_{6} / a_{5}=r_{0}, b_{6}-$ $b_{5}=r_{1}, \quad d_{6}=r_{3}, \quad-e_{5}=r_{4 } 。\quad r_{0}, \quad r_{1}, \quad r_{2}, \quad r_{3} $和r4为联合估计参数,其初值为树枝和树叶生物量独立模型的参数估计值。进行模型参数估计值求解时要在方程两边同时乘以各分项的权重函数(表 5)。参数拟合结果及评价指标见表 6。由表 6可知,各项拟和的决定系数R2较高,除树枝、树叶生物量仅为85.2%和81.5%外,其他各项生物量的R2均大于90%。除树叶生物量外,其他各项生物量的ETR和EMS值均在±10%内,地上、树干和树根生物量预估精度P值也均达90%以上,树冠、树叶和树枝生物量P值相对较低,但也大于70%,模型具有一定的预估性。
表 6 相容性模型参数估计值、拟合优度和评价指标
Table 6. Measurement error, fitting optimization and evaluating indicators of compatible models
项目 参数估计值 拟合优度 评价指标 r0 r1 r2 r3 r4 R2 ESE ETR EMS EMPS P 地上 0.249 1 -0.044 3 -0.184 0 0.145 2 0.966 34.982 1.665 0.936 10.320 90.9 树根 0.910 9.190 1.556 0.813 12.328 90.2 树干 0.000 2 1.574 6 -1.114 3 -0.151 7 1.128 7 0.974 24.177 1.254 -0.377 9.499 91.6 树冠 0.917 15.802 2.888 5.336 21.521 85.1 树枝 1.726 7 1.161 0 0.229 1 -1.594 9 0.852 11.172 1.845 3.786 21.647 86.8 树叶 0.815 6.827 6.583 12.180 27.930 71.0 -
枝叶生长不仅受年龄、林分密度、气候环境等的影响[17-19],而且还受其在树冠垂直分布的影响,因为不同的空间分布会影响枝叶对光照的吸收和有机物质的积累[20]。卢军等[21]对帽儿山天然次生林主要阔叶树种叶量分布模拟研究表明:天然次生林阔叶树种的叶量在树冠上部所占比例很小,在树冠中部及中下部叶量所占比例较大,几乎集中了整个树冠60%~70%的叶量。本研究发现:树冠下层活枝生物量显著高于上层,而下层树叶生物量略大于上层,但表现并不显著。这可能是因为天然次生林中尽管树冠上层光照充足,但本身枝条较小,着生的叶量也因此较少,而树冠下层枝条虽大,但获得的光照较少,且易受周围林木的遮挡,导致着生的叶量也不是很多,从而上下层树叶生物量差异不显著。玉宝等[22]对兴安落叶松Larix gmelinii天然林单株树冠生物量分配研究表明:树枝生物量分配比高于树叶,并因其相对直径大小的不同而不同。本研究也得出相似的结论,且研究发现叶生物量占比随枝径的增大而逐渐减少,一级枝和次级枝生物量占比则呈波动变化。究其原因可能是树冠下层的枝条多为已经生长的直径较大的活枝,但位于树冠下层的枝条因受遮蔽等影响使得获取的光照资源较少,从而导致着生在枝条上的阔叶量相对减少,而光照的缺乏对一级枝和次级枝自身物质积累影响不大。活枝分配比呈波动变化的原因可能与环境因素或树种生态学特征有关[23],如林木一定条件下的自然整枝。
植物的个体生理生态学特征、林内环境、立地空间是影响植物器官生物量积累及分配差异的重要原因[24]。相关研究表明:通常植株地上部分生物量占植株总生物量的79.0%,而地下生物量约占21.0%[25],这与本研究得出的次生林中闽楠地下生物量占全株生物量的19.0%有差异。同时,董点等[26]研究表明:紫椴Tilia amurensis树根生物量占全株生物量的24.2%,刘坤等[10]发现银杏Gingo biloba根生物量占全株生物量的26.0%,造成这种差异的原因可能是因为树种间本身特性及立地环境的不同导致,如闽楠适合生长在土层深厚、肥沃、排水良好的土壤,多分布在峡谷、山洼和河流两岸的台地等,而这些地方往往光照不足,因此树体相对减少树根有机质积累,增加树干及树冠等有机质积累以获取更多的光照资源,加速生长。王宁等[27]对山杨Populus davidiana生物量分配研究表明:各器官生物量分配比从小到大依次为树叶、树枝、树根、树干,本研究结果与之相一致。植物生物量分配特征与胸径、年龄等因子密切相关,如余碧云等[28]对栓皮栎Quercus variabilis生物量研究表明:树干生物量分配比随胸径的增加呈先减小后逐渐增大的趋势,而枝叶生物量分配比则随胸径的增加呈先增后减再增的变化趋势。本研究表明:叶生物量占比随胸径增大呈先增后减,树干生物量占比表现为先减后增的波动变化,而树根生物量占比则逐渐减小。一方面可能是由于幼林时期植株相对较小,枝叶及根部具有充足的营养资源及生长空间进行物质积累,而随着幼苗的逐渐生长,生长空间不断减小,周围林木的遮蔽影响树冠的生长,树枝生物量减小;为获得林木上层充足资源,林木开始积累主干生物量而削弱对树根生物量的积累。另一方面,闽楠在一定阶段的自然整枝以及林内的环境变化也会影响枝叶生物量占比。这也表明,前期闽楠幼苗主要靠树根生长获取营养资源,随着径级增加,逐渐转为主干有机质的积累,这有利于形成大径材。
-
不少研究学者对林木生物量模型进行研究。如程堂仁等[29]为准确预估小陇山林区的森林生物量以及碳汇能力,通过标准木资料建立了锐齿槲栎Quercus aliena等8种树种的各器官生物量回归方程。魏海燕等[30]采用幂函数回归模型对甜槠Castanopsis eyrei单木生物量数据进行拟合,得到各器官生物量最优模型,除树枝、树叶生物量预估精度较差外,其余各器官生物量预估精度均达90%以上。本研究建立的相容性模型系统对闽楠全株、树根、地上部分、树冠、树干、树枝、树叶生物量的拟合精度分别为91.8%,90.2%,91.0%,85.1%,91.6%,86.8%和71.0%,除叶生物量外,其余各分项生物量预估精度值均大于85%,总相对误差均在3%以下,能较准确地预估研究区次生林中闽楠全株及各部分生物量。树冠、树枝和树叶的模型预估精度较差,可能是树叶本身受环境影响因素较大等原因造成的。树叶是光合作用的最重要场所,对周围光照条件敏感,因此异质性较大[31],另一方面也可能是次生林中阔叶树树枝易分叉,尽管在枝条数据获取时考虑到分叉的枝条,但在实际取样时还是会出现抽样误差,同时模型指标的选取及评价标准也可能是导致树叶等生物量预估精度较低。
相容性模型主要是协调各分量间的关系,使其预估值等于总体和相应的分项。相关研究对独立模型和相容性模型的生物量预估效果评价表明:这2种方法对总量和各分量的预估精度没有很大差异[11]。本研究也得出类似的结论。为更全面地分析林分因子对生物量的影响,本研究在构建生物量模型时引入除胸径、树高之外的冠幅和竞争因子,结果表明:引入与生物量相关性较强的因子能在一定程度上提高模型的拟合优度(R2)以及估测精度。由于研究区域立地条件差异不大,因此在构建生物量模型时未考虑随机效应和区域差异。同时采伐解析木受到限制,本研究生物量样本数据相对较少,特别是小径阶的样本,这将会影响所建立的模型精度,有待补充这部分数据并做深入研究。
Biomass allocation characteristics and a compatible model for Phoebe bournei in a natural secondary forest
-
摘要: 闽楠Phoebe bournei为中国特有树种,是国家Ⅱ级重点保护植物,具有重要的经济价值和生态价值。以江西安福县闽楠天然次生林为研究对象,通过标准地及生物量调查获取基础数据,分析闽楠生物量分配特征,并基于非线性度量误差法建立相容性生物量模型。结果表明:①不同枝径级(2,4,6,8 cm)的一级枝、次级枝、叶生物量占枝条生物量的比值均差异显著(P < 0.05),枝条生物量集中于树冠下层,且显著高于上层生物量(P < 0.05)。②各器官生物量占整株生物量的比值从大到小依次为树干、树根、树枝、树叶,不同胸径级(14,18,22,26,30 cm及以上)各器官生物量大小差异显著(P < 0.05),不同胸径级树根生物量占整株生物量的比值差异显著(P < 0.05),其余各器官生物量占比均差异不显著(P > 0.05)。③相容性生物量模型对全株、地上、根、树冠、干、枝和叶的拟合精度分别为91.8%,91.0%,90.2%,85.1%,91.6%,86.8%和71.0%,拟合效果较好。Abstract: As a unique species of China, Phoebe bournei, a protected second-level rare and vulnerable tree species, has had important ecological and economic value. Within the research area of Anfu County in Jiangxi Province, a natural secondary P. bournei forest was taken as the research object, and data was obtained by means of a typical sample investigation and whole tree harvesting. Biomass allocation patterns of total tree and each component (stem, root, branch, and leaf) of P. bournei were analyzed with branch diameter levels of 2, 4, 6, and 8 cm and with DBH levels of 14, 18, 22, 26, and 30 cm as well as above. Compatible biomass models were established using a nonlinear measurement error model. Results showed (1) Significant differences among the biomass allocation ratios of primary branch, secondary branch, and leaf at different branch diameter levels (P < 0.05). Live branch biomass mainly occurred in the lower canopy layers, and was significantly higher than the upper layer (P < 0.05). (2) Biomass allocation ratios of each organ were as follows: stem > root > branch > leaf. The biomass of organs at different DBH levels was significantly different (P < 0.05). The biomass allocation ratio of roots was significantly different (P < 0.05); whereas, the biomass allocation ratios of other organs were not significantly different (P > 0.05). (3) The precision values of the compatible biomass model based on the nonlinear measurement error model for biomass were whole plant (91.8%), aboveground (91.0%), root (90.2%), crown (85.1%), stem (91.6%), branch (86.8%), and leaf (71.0%). Overall, the model was a good fit.
-
Key words:
- forest ecology /
- Phoebe bournei /
- natural secondary forest /
- biomass allocation /
- compatible mode
-
图 1 不同枝径阶枝、叶生物量分配比
Figure 1 Biomass allocation ratios of branch and leaf at different branch diameters levels
图 2 不同冠层各组分生物量分配比
Figure 2 Biomass allocation ratio of each component at different crown levels
图 3 不同径级各器官生物量分配比
Figure 3 Biomass allocation ratio of each tree organ at different DBH levels
表 1 不同胸径阶树冠生物量分配特征
Table 1. Biomass allocation ratios of tree crown at different DBH levels
径阶/cm 生物量/(kg·株-1) 一级枝 次级枝 叶 总 14 15.75 ± 5.59 a 14.04 ± 3.80 ab 7.23 ± 3.74 a 33.51 ± 13.12 a 18 16.19 ± 3.20 a 11.49 ± 2.58 a 10.83 ± 2.50 a 38.51 ± 8.10 a 22 30.41 ± 6.53 a 31.60 ± 6.09 b 16.56 ± 1.68 ab 78.56 ± 13.86 a 26 39.13 ± 12.20 a 22.31 ± 1.40 ab 17.09 ± 2.05 ab 82.73 ± 11.73 a >30 79.57 ± 6.30 b 50.34 ± 2.36 c 25.36 ± 1.24 b 155.28 ± 7.40 b 说明:同列不同字母表示同一指标不同径阶间差异显著(P<0.05)  下载: 导出CSV
下载: 导出CSV
表 2 不同胸径阶各器官生物量特征
Table 2. Biomass allocation of each tree organ at different DBH levels
径阶/cm 株数 生物量/(kg·株-1) 树干 树枝 树叶 地上部分 树根 全株 14 2 70.42 ± 10.82 a 21.01 ± 1.77 a 7.42 ± 1.05 a 98.84 ± 0.54 a 27.64 ± 11.55 a 126.49 ± 11.01 a 18 5 108.91 ± 15.73 ab 27.24 ± 2.25 a 11.68 ± 1.59 ab 147.83 ± 2.78 a 37.55 ± 15.80 ab 185.38 ± 18.31 a 22 4 161.24 ± 12.87 b 45.40 ± 3.20 b 12.59 ± 0.70 ab 219.22 ± 3.18 b 54.23 ± 13.47 b 273.45 ± 15.71 b 26 3 233.13 ± 14.08 c 54.83 ± 5.53 b 17.09 ± 2.05 b 305.05 ± 0.62 c 72.75 ± 7.78 c 377.80 ± 8.36 c >30 3 407.64 ± 24.18 d 131.44 ± 6.81 c 32.18 ± 4.39 c 571.26 ± 9.71 d 97.42 ± 16.09 d 668.68 ± 24.73 d 均值 191.33 ± 28.14 54.04 ± 9.31 15.97 ± 2.17 261.33 ± 74.21 57.09 ± 6.03 318.42 ± 44.32 说明:同列不同字母表示同一器官不同径阶间差异显著(P<0.05)
下载: 导出CSV
表 3 林木特征因子与各器官或各组分生物量之间的相关系数
Table 3. Correlation coefficient between tree characteristics and biomass of various organs or components
特征因子 树干 树枝 树叶 树根 树冠 地上部分 全株 年龄 0.499* 0.341 0.140 0.379 0.307 0.425 0.422 胸径 0.946** 0.886** 0.785** 0.930** 0.878** 0.947** 0.951** 树高 0.620** 0.404 0.326 0.582* 0.395 0.568* 0.574* 冠幅 0.761** 0.844** 0.689** 0.714** 0.825** 0.797** 0.791** 树冠体积 0.829** 0.862** 0.764** 0.767** 0.855** 0.855** 0.849** 冠长 0.494* 0.332 0.352 0.473 0.340 0.460 0.465 说明:*表示显著相关,**表示极显著相关
下载: 导出CSV
表 4 总量(W)与各分量独立模型的参数估计值和拟合优度
Table 4. Measurement error and fitting optimization of independent models
项目 模型形式 拟合优度 R2 RAdj2 ESE CAI lnW=r0+r1 ln(D2H) 0.963 0.960 21.011 12.27 树干 lnW=r0+r1 lnD+r2 ln(D2H) 0.964 0.959 23.076 12.83 lnW=r0+r1 lnD+r2 ln(D2H)+r3 lnIC 0.965 0.957 22.147 13.56 lnW=r0+r1 lnD 0.830 0.818 17.532 -0.34 树枝 lnW=r0+r1 lnD+r4 lnWC 0.852 0.831 15.803 -0.37 lnW=r0+r1 lnD+r3 lnIC+r4WC 0.858 0.825 19.124 -0.14 lnW=r0+r1 lnD 0.595 0.568 9.248 -3.23 树叶 lnW=r0+r1 lnD+r3 lnIC 0.718 0.677 5.361 -3.49 lnW=r0+r1 lnD+r3 lnIC+r4WC 0.719 0.654 5.312 -3.20 lnW=r0+r1 lnD 0.807 0.795 20.124 -0.64 树冠 lnW=r0+r1 lnD+r3 lnIC 0.815 0.788 19.452 -0.14 lnW=r0+r1 lnD+r3 lnIC+r4WC 0.833 0.795 19.024 -0.72 地上部分 lnW=r0+r1 lnD 0.937 0.933 38.243 9.28 lnW=r0+r1 ln(D2H) 0.946 0.943 37.156 9.69 lnW=r0+r1 lnD+r2 ln(D2H) 0.954 0.948 37.425 9.18 lnW=r0+r1 lnD 0.871 0.863 9.246 7.41 树根 lnW=r0+r1 ln(D2H) 0.876 0.868 8.725 6.34 lnW=r0+r1 lnD+r2 ln(D2H)+r3 lnIC 0.885 0.859 8.829 6.29 lnW=r0+r1 lnD 0.935 0.931 42.576 9.73 全株 lnW=r0+r1 lnD+r2 ln(D2H) 0.952 0.945 41.086 9.57 lnW=r0+r1 lnD+r2 ln(D2H)+r3 lnIC 0.953 0.941 40.536 11.22
下载: 导出CSV
表 5 总量(W)与各分量最优独立模型形式与参数估计值
Table 5. Model form of optimal independent model
项目 模型形式 参数估计值 评价指标 a b c d e ETR EMS EMPS P 树干 lnW=r0+r1 ln(D2H) 5.366 0.936 1.203 0.653 9.429 93.1 树枝 lnW=r0+r1 lnD+r4 lnWC -2.639 1.751 0.560 5.106 3.069 21.866 80.8 树叶 lnW=r0+r1 lnD+r3 lnIC 1.253 0.488 -0.649 4.097 3.584 21.834 78.2 树冠 lnW=r0+r1 lnD+r3 lnIC+r4WC -0.368 1.143 -0.301 0.521 4.188 2.765 21.161 82.4 地上部分 lnW=r0+r1 lnD+r2 ln(D2H) 2.853 0.886 0.540 1.601 0.664 10.293 90.9 树根 lnW=r0+r1 lnD+r2 ln(D2H)+r3 lnIC 1.889 0.710 0.356 0.005 1.357 1.124 11.936 90.2 全株 lnW=r0+r1 lnD+r2 ln(D2H) 3.124 0.863 0.500 1.646 0.848 9.901 91.8 说明:各分量权重函数树干1/D2.329,树枝1/D2.203,树叶1/D1.252,树冠1/D2.996,地上部分1/D3.156,树根1/D2.007;各权函数F检验均显著(P<0.05)
下载: 导出CSV
表 6 相容性模型参数估计值、拟合优度和评价指标
Table 6. Measurement error, fitting optimization and evaluating indicators of compatible models
项目 参数估计值 拟合优度 评价指标 r0 r1 r2 r3 r4 R2 ESE ETR EMS EMPS P 地上 0.249 1 -0.044 3 -0.184 0 0.145 2 0.966 34.982 1.665 0.936 10.320 90.9 树根 0.910 9.190 1.556 0.813 12.328 90.2 树干 0.000 2 1.574 6 -1.114 3 -0.151 7 1.128 7 0.974 24.177 1.254 -0.377 9.499 91.6 树冠 0.917 15.802 2.888 5.336 21.521 85.1 树枝 1.726 7 1.161 0 0.229 1 -1.594 9 0.852 11.172 1.845 3.786 21.647 86.8 树叶 0.815 6.827 6.583 12.180 27.930 71.0
下载: 导出CSV
-
[1] 高志雄, 王新杰, 王廷蓉, 等.福建杉木单木生物量分配及相容性模型的应用[J].南京林业大学学报(自然科学版), 2015, 39(4):157-162. GAO Zhixiong, WANG Xinjie, WANG Tingrong, et al. Biomass allocation and application of compatible model of Cunninghamia lanceolata in Fujian area[J]. J Nanjing For Univ Nat Sci Ed, 2015, 39(4):157-162. [2] 肖生苓, 杨嘉龙.大兴安岭北部兴安落叶松天然林单木地上生物量[J].林业科学, 2014, 50(8):22-29. XIAO Shengling, YANG Jialong. Individual tree aboveground biomass of Larix gmelinii natural forest in the northern greater Khingan mountains[J]. Sci Silv Sin, 2014, 50(8):22-29. [3] 董利虎, 李凤日, 贾炜玮, 等.含度量误差的黑龙江省主要树种生物量相容性模型[J].应用生态学报, 2011, 22(10):2653-2661. DONG Lihu, LI Fengri, JIA Weiwei, et al. Compatible biomass models for main tree species with measurement error in Heilongjiang Province of Northeast China[J]. Chin J Appl Ecol, 2011, 22(10):2653-2661. [4] 丁鑫, 肖建华, 黄建峰, 等.珍贵木材树种楠木的野生资源调查[J].植物分类与资源学报, 2015, 37(5):629-639. DING Xin, XIAO Jianhua, HUANG Jianfeng, et al. Investigation on wild resource of valuable timber tree Phoebe zhennan(Lauraceae)[J]. Plant Diver Resour, 2015, 37(5):629-639. [5] 吴大荣.福建罗卜岩闽楠(Phoebe bournei)林中优势树种生态位研究[J].生态学报, 2001, 21(5):851-855. WU Darong. A study on the niche of dominant species in Phoebe bournei forests in Luoboyan nature reserve of Fujian[J]. Acta Ecol Sin, 2001, 21(5):851-855. [6] 游晓庆, 潘萍, 彭诗涛, 等.闽楠天然次生林树种间联结性分析[J].安徽农业大学学报, 2017, 44(4):630-635. YOU Xiaoqing, PAN Ping, PENG Shitao, et al. Interspecific association of Phoebe bournei natural secondary forest[J]. Acta Agric Univ Anhui, 2017, 44(4):630-635. [7] 吴大荣, 朱政德.福建省罗卜岩自然保护区闽楠种群结构和空间分布格局初步研究[J].林业科学, 2003, 39(1):23-30. WU Darong, ZHU Zhengde. Preliminary study on structure and spatial distribution pattern of Phoebe bournei in Luoboyan nature reserve in Fujian Province[J]. Sci Silv Sin, 2003, 39(1):23-30. [8] 李雪云, 潘萍, 臧颢, 等.闽楠天然次生林自然更新的影响因子研究[J].林业科学研究, 2017, 30(5):701-708. LI Xueyun, PAN Ping, ZANG Hao, et al. Study on factors affecting natural regeneration of natural secondary Phoebe bournei forest[J]. For Res, 2017, 30(5):701-708. [9] 游晓庆.闽楠天然次生林林分空间结构及种间关联性研究[D].南昌: 江西农业大学, 2016. YOU Xiaoqing. Study on Spatial Structure of Natural Secondary Phoebe bournei Forest and Its Interspecific Association[D]. Nanchang: Jiangxi Agricultural University, 2016. [10] 刘坤, 曹林, 汪贵斌, 等.银杏生物量分配格局及异速生长模型[J].北京林业大学学报, 2017, 39(4):12-20. LIU Kun, CAO Lin, WANG Guibin, et al. Biomass allocation patterns and allometric models of Ginkgo biloba[J]. J Beijing For Univ, 2017, 39(4):12-20. [11] 汪金松, 张春雨, 范秀华, 等.臭冷杉生物量分配格局及异速生长模型[J].生态学报, 2011, 31(14):3918-3927. WANG Jinsong, ZHANG Chunyu, FAN Xiuhua, et al. Biomass allocation patterns and allometric models of Abies nephrolepis Maxim[J]. Acta Ecol Sin, 2011, 31(14):3918-3927. [12] 唐守正, 张会儒, 胥辉.相容性生物量模型的建立及其估计方法研究[J].林业科学, 2000, 36(增刊1):19-27. TANG Shouzheng, ZHANG Huiru, XU Hui. Study on establish and estimate method of compatible biomass model[J]. Sci Silv Sin, 2000, 36(suppl 1):19-27. [13] 董利虎, 李凤日, 贾炜玮.东北林区天然白桦相容性生物量模型[J].林业科学, 2013, 49(7):75-85. DONG Lihu, LI Fengri, JIA Weiwei. Compatible tree biomass models for natural white birch (Betula platyphylla) in northeast China forest area[J]. Sci Silv Sin, 2013, 49(7):75-85. [14] 钱升平, 吕飞舟, 边更战, 等.基于简单竞争指标的青冈栎次生林林木竞争单元构建[J].林业资源管理, 2016(2):56-61. QIAN Shengping, LÜ Feizhou, BIAN Gengzhan, et al. Struture of tree competition unit of Cyclobalanopsis glauca sencodary forest based on simple competition index[J]. For Resour Manage, 2016(2):56-61. [15] 惠刚盈, 胡艳波.混交林树种空间隔离程度表达方式的研究[J].林业科学研究, 2001, 14(1):23-27. HUI Gangying, HU Yanbo. Measuring species spatial isolation in mixed forests[J]. For Res, 2001, 14(1):23-27. [16] 曾伟生, 唐守正.立木生物量方程的优度评价和精度分析[J].林业科学, 2011, 47(11):106-113. ZENG Weisheng, TANG Shouzheng. Goodness evaluation and precision analysis of tree biomass equations[J]. Sci Silv Sin, 2011, 47(11):106-113. [17] 钟泰林, 朱廷朴, 来水方.中华九龙桂枝条自然弯曲与气温的相关性[J].浙江林学院学报, 2006, 23(1):109-111. ZHONG Tailin, ZHU Tingpu, LAI Shuifang. Correlation between the natural curve of Osmanthus fragrans 'Jiulonggui' with air temperatures[J]. J Zhejiang For Coll, 2006, 23(1):109-111. [18] 涂洁, 刘琪璟.中亚热带红壤丘陵区湿地松枝条生长规律[J].浙江林学院学报, 2007, 24(2):162-167. TU Jie, LIU Qijing. Branch growth of Pinus elliottii in the hilly red-soil area of the mid-subtropical climatic zone[J]. J Zhejiang For Coll, 2007, 24(2):162-167. [19] 郑亚琼, 冯梅, 李志军.胡杨枝芽生长特征及其展叶物候特征[J].生态学报, 2015, 35(4):1198-1207. ZHENG Yaqiong, FENG Mei, LI Zhijun. Investigation of bud burst, shoot growth and leaf expansion in Populus euphratica of different ages[J]. Acta Ecol Sin, 2015, 35(4):1198-1207. [20] SCHMID I. The influence of soil type and interspecific competition on the fine root system of Norway spruce and European beech[J]. Basic Appl Ecol, 2002, 3(4):339-346. [21] 卢军, 李凤日, 张会儒, 等.帽儿山天然次生林主要阔叶树种叶量分布模拟[J].林业科学, 2011, 47(12):114-120. LU Jun, LI Fengri, ZHANG Huiru, et al. Simulation of foliage distribution for major broad-leaved species in secondary forest in Mao'er mountain[J]. Sci Silv Sin, 2011, 47(12):114-120. [22] 玉宝, 乌吉斯古楞, 王百田, 等.兴安落叶松天然林树冠生长特性分析[J].林业科学, 2010, 46(5):41-48. YU Bao, WU Jisiguleng, WANG Baitian, et al. Analysis of crown growth characteristics in natural Larix gmelinii forests[J]. Sci Silv Sin, 2010, 46(5):41-48. [23] BRIX H. Effects of thinning and nitrogen fertilization on branch and foliage production in Douglas-fir[J]. Can J For Res, 1981, 11(3):205-511. [24] LEWIS S L, TANNER E V J. Effects of above-and below-ground competition on growth and survival of rain forest tree seedlings[J]. Ecology, 2000, 81:2525-2538. [25] HOUGHTON R A, LAWRENCE K T, HACKLER J L, et al. The spatial distribution of forest biomass in the Brazilian Amazon:a comparison of estimates[J]. Global Change Biol, 2001, 7(7):731-746. [26] 董点, 林天喜, 唐景毅, 等.紫椴生物量分配格局及异速生长方程[J].北京林业大学学报, 2014, 36(4):54-63. DONG Dian, LIN Tianxi, TANG Jingyi, et al. Biomass allocation patterns and allometric models of Tilia amurensis[J]. J Beijing For Univ, 2014, 36(4):54-63. [27] 王宁, 王百田, 王瑞君, 等.晋西山杨和油松生物量分配格局及异速生长模型研究[J].水土保持通报, 2013, 33(2):151-155. WANG Ning, WANG Baitian, WANG Ruijun, et al. Biomass allocation patterns and allometric models of Populous davidiana and Pinus tabulaeformis Carr. in west Shanxi Province[J]. Bull Soil Water Conserv, 2013, 33(2):151-155. [28] 余碧云, 张文辉, 胡晓静, 等.栓皮栎地上部分构型及生物量分配[J].应用生态学报, 2015, 26(8):2265-2272. YU Biyun, ZHANG Wenhui, HU Xiaojing, et al. Aboveground architecture and biomass distribution of Quercus variabilis[J]. Chin J Appl Ecol, 2015, 26(8):2265-2272. [29] 程堂仁, 冯菁, 马钦彦, 等.甘肃小陇山锐齿栎林生物量及其碳库研究[J].北京林业大学学报, 2007, 29(增刊2):209-215. CHENG Tangren, FENG Jing, MA Qinyan, et al. Forest biomass and carbon storage of Quercus aliena var. acuteserrata stand-types in Xiaolong mountains, Gansu Province[J]. J Beijing For Univ, 2007, 29(suppl 2):209-215. [30] 魏海燕, 王军才, 刘济明, 等.雷山常绿阔叶林甜槠栲的单木生物量估测[J].贵州农业科学, 2015, 43(4):163-167, 172. WEI Haiyan, WANG Juncai, LIU Jiming, et al. Estimation on single-tree biomass of evergreen broad-leaved forest with Castanopsis eyrei in Leishan[J]. Guizhou Agric Sci, 2015, 43(4):163-167, 172. [31] FORRESTER D I, TACHAUER I H H, BARBEITO I. Generalized biomass and leaf area allometric equations for European tree species incorporating stand structure, tree age and climate[J]. For Ecol Manage, 2017, 396:160-175. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2019.04.017
 点击查看大图
点击查看大图
计量
- 文章访问数: 4076
- HTML全文浏览量: 898
- PDF下载量: 45
- 被引次数: 0
