




-
光系统Ⅱ(PSⅡ)是植物对温度胁迫最敏感的光合系统组分[1-3],利用非损伤、检测时间短、灵敏度高的快速叶绿素荧光诱导动力学技术[4-5],可以方便地研究温度与PSⅡ光化学活性之间的关系。低温胁迫会引起快速叶绿素荧光诱导动力学曲线(OJIP曲线)趋向平缓、O相升高、J相下降或上升、I相和P 相下降、K 点和L点出现[6-8]等变化,反映出PSⅡ有活性的反应中心数量减少[9],反应中心吸收和热耗散的能量增加[9-11],以及PSⅡ受体侧的电子传递速率降低等光合作用的“内在变化”[11-12]。低温胁迫还会使叶片叶绿素降解[12-13],导致光合作用的物质基础发生改变。目前多数研究采用离体试验材料、野外观测、设置单一温度处理等方式,温度与PSⅡ光化学活性的关系难以被准确描述。为获得不同胁迫程度下叶绿素荧光的综合性指标,当前研究主要利用隶属函数法[14-15]、主成分分析法[16-17]、TOPSIS综合评价法[18-19]和综合生物反应指数(integrated biomarker response,IBR)[20]计算综合指标。其中IBR评价法具有操作简单、实用性强和应用广泛的特点,但尚未用于计算低温胁迫下的叶绿素荧光综合指标。南极假山毛榉Nothofagus antarctica为壳斗科Fagaceae假山毛榉属Nothofagus落叶乔木或灌木,主要分布在南美洲南部的温带森林,耐寒能力极强[21],为探索低温环境下南极假山毛榉光合生理特性,本研究采用连续测定与控制试验的方法,观测低温胁迫下南极假山毛榉叶片相对叶绿素含量(relative chlorophyll content)、OJIP曲线特征、快速叶绿素荧光诱导动力学测量(JIP-test)参数等的变化,并将JIP-test参数代入IBR评价法,用以阐明低温胁迫对南极假山毛榉PSⅡ光化学活性的影响规律,并检验IBR评价法用于计算低温胁迫下的叶绿素荧光综合指标的可行性。
HTML
-
供试材料为2年生南极假山毛榉实生苗(山西省林业科学研究院基地培育),株高(50±5) cm。2018年9月初,选取长势良好,无病虫害,大小相近的南极假山毛榉5盆,置于人工气候箱中适应培养3 d。人工气候箱参数为:温度25 ℃;昼夜相隔时间12 h;光照强度500 μmol·m−2·s−1;相对湿度(60±3)%,根据表土干燥情况适量浇水。设置25、15、5、−5、−15和−25 ℃等6个测试温度,从25 ℃依次降低,降温速率为2 ℃·h−1,待植株在5 ℃的人工气候箱中测试完毕,转移到3 ℃预冷的冰柜中,继续以2 ℃·h−1的速率依次降到−5、−15和−25 ℃;各温度下处理时间为24 h。冰柜装有人工光源,光照强度500 μmol·m−2·s−1,采用大气采样仪(QC-1S,北京市科安劳保新技术公司)从冰柜门缝隙处向内缓慢送气。
各测试温度下处理完成后,在原处理环境下关闭光源,使植株暗适应20 min;立刻进行快速叶绿素荧光诱导动力学测量,随后立即测定相对叶绿素含量。同1株上的2项测量均在同1片健康、成熟的叶片上进行,测量位置相互错开。每个处理5盆,每盆作为1个独立重复实验,测定5片·盆−1。以25 ℃处理为对照(ck)。10:00测定各项指标。
-
采用叶绿素测定仪(SPAD-502Plus,KONICA MINOLTA,日本)测定各温度处理下植株的相对叶绿素含量(SPAD值)。
-
采用叶绿素荧光仪(Yaxin-116G,雅欣理仪公司)测定快速叶绿素荧光诱导动力学曲线(OJIP),测定前暗适应20 min,选取叶肉部分用3 000 μmol·m−2·s−1饱和蓝闪光照射1 s,以10 μs (300 μs之前)、100 μs (300 μs~3 ms)和1 ms (3 ms之后)的间隔记录荧光信号,并计算叶绿素荧光动力学参数。
-
叶片经过暗适应后再给予饱和蓝闪光照射,散发的荧光随时间变化的曲线,构成叶绿素荧光诱导动力学曲线。暗适应后出现最小荧光O相,此时PSⅡ处于“完全开放”状态;照射强光后,陆续出现J、I相,最终由于PSⅡ反应中心关闭,达到P相。将不同温度处理的OJIP曲线离差标准化。应用公式VO-P=(Ft−FO)/(FP−FO)计算FO与FP间相对可变荧光强度(VO-P),其中Ft表示t时的荧光强度,FO表示暗适应后的最小荧光强度,FP表示暗适应后的最大荧光强度。应用公式△VO-P=VO-P(处理)−VO-P(ck)计算FO与FP间相对可变荧光强度差值(△VO-P)。FO与FJ、FK间的相对可变荧光强度差值(△VO-J和△VO-K)的计算方式同VO-P和△VO-P。
-
暗适应后照光,O、L、K、J、I、P点分别表示最小荧光、0.15 ms荧光,0.30 ms荧光,2.00 ms荧光、30.00 ms荧光和最大荧光[22],其中0.15和0.30 ms时的相对荧光强度用VL和VK表示,PSⅡ单位面积反应中心数量(RC/CSO)、单位反应中心吸收的能量(ABS/RC)、单位反应中心以热能形式耗散的能量(DIO/RC)、单位反应中心捕获的用于还原初级醌受体(QA)的能量(TRO/RC)、单位反应中心用于将电子从QA−传递到质体醌(PQ)的能量(ETO/RC)、单位反应中心用于将电子从QA−传递到光系统Ⅰ(PSⅠ)的能量(REO/RC)、PSⅡ光化学初级最大效率(φPO,意义与暗适应下PSⅡ的最大量子产额Fv/Fm相同)、PSⅡ捕获的电子从QA−传递到PQ的效率(ψEO)、电子从PQH2传递到PSⅠ最终受体侧的效率(δRO)、叶片性能指数(PIABS)等参数计算参照文献[22]和[23]。
-
计算方法参照文献并略有改动:①计算各项荧光参数在所有处理下的平均值m和标准差s;②代入公式(1)对各处理下的各项荧光参数(X)标准化,得到Y;③对不同温度处理下的各项荧光参数进行回归拟合,如果某项荧光参数的拟合曲线随着温度降低而上升,则Z=Y,反之则Z=−Y;④依据公式(2)计算不同温度处理下单项荧光参数的得分Si,其中|Zm
in|是某温度处理下所有荧光参数Z值的最小值的绝对值;⑤以某温度处理下某项荧光参数的Si值作为辐射线的长度绘制星状图,各温度处理下的综合生物反应指数(IBR)由顺时针相邻荧光参数的辐射线围成的三角形面积Ai之和得到公式(3),公式(3)中的Ai依据公式(4)计算,公式(4)中的n为参与计算的荧光参数的个数。 -
将各处理所得的IBR、Fv/Fm、
PIABS、相对叶绿素含量进行相关性分析,依据Pearson参数和显著性,检验IBR评价法用于计算低温胁迫下的叶绿素荧光综合指标的可行性。采用SPSS 22进行数据统计分析,应用最小显著差法(LSD)进行多重比较,采用Origin 2018进行回归分析和绘图。
1.1. 试验材料与处理方法
1.2. 测定方法
1.2.1. 相对叶绿素含量测定
1.2.2. 快速叶绿素荧光诱导动力学测量(JIP-test)
1.3. 数据处理
1.3.1. OJIP曲线处理
1.3.2. 快速叶绿素荧光诱导动力学曲线参数计算
1.3.3. 综合生物反应指数(IBR)计算
1.3.4. 相关性分析和多重比较
-
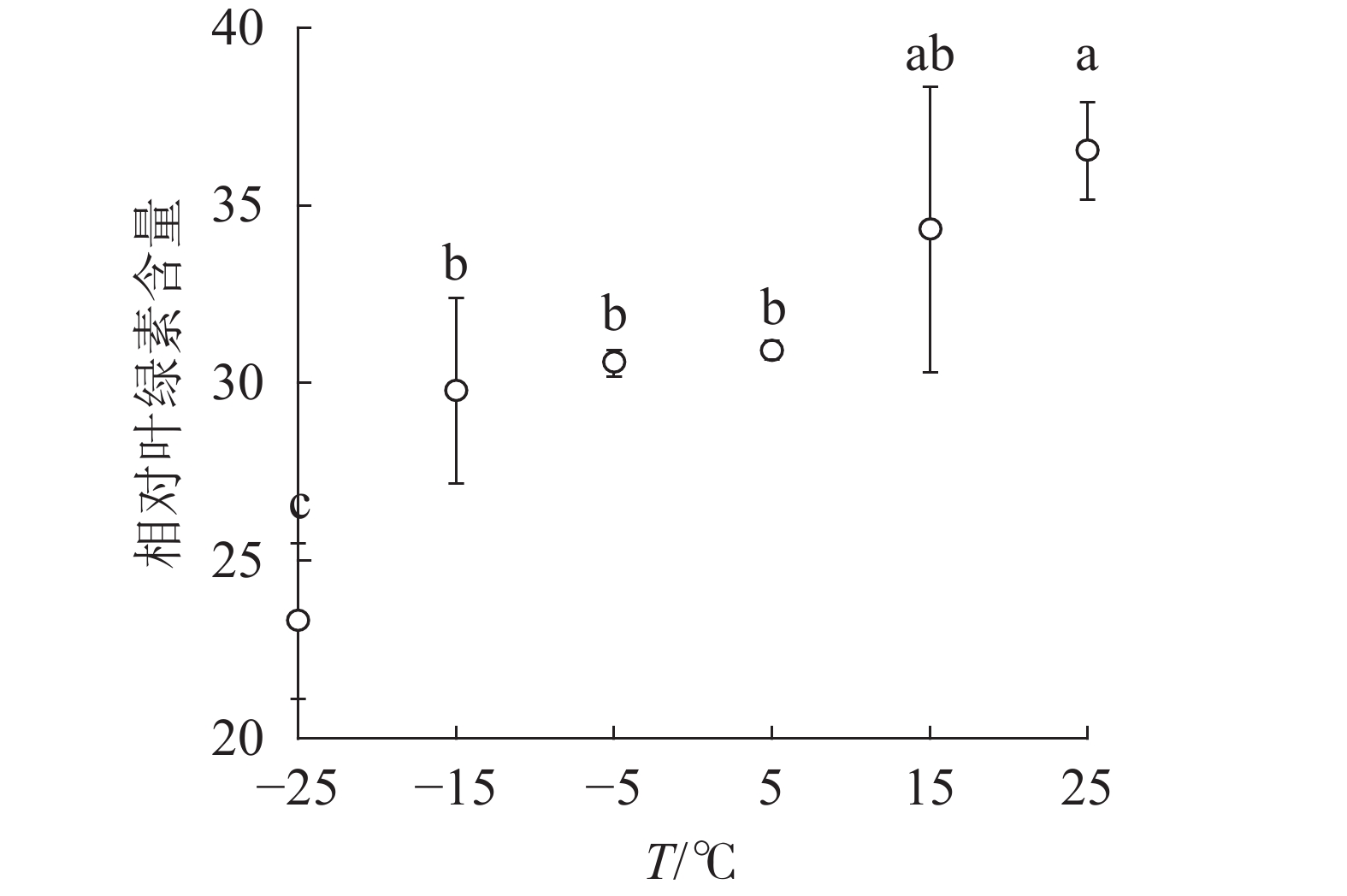
由图1可知:15和5 ℃时相对叶绿素含量无显著变化,−5 ℃时开始显著下降,−25 ℃时极显著降低,与对照相比降低了63.8%。表明≤−5 ℃的低温环境会导致南极假山毛榉叶片相对叶绿素含量显著降低。
Figure 1. SPAD value of leaves under different temperature treatments
-
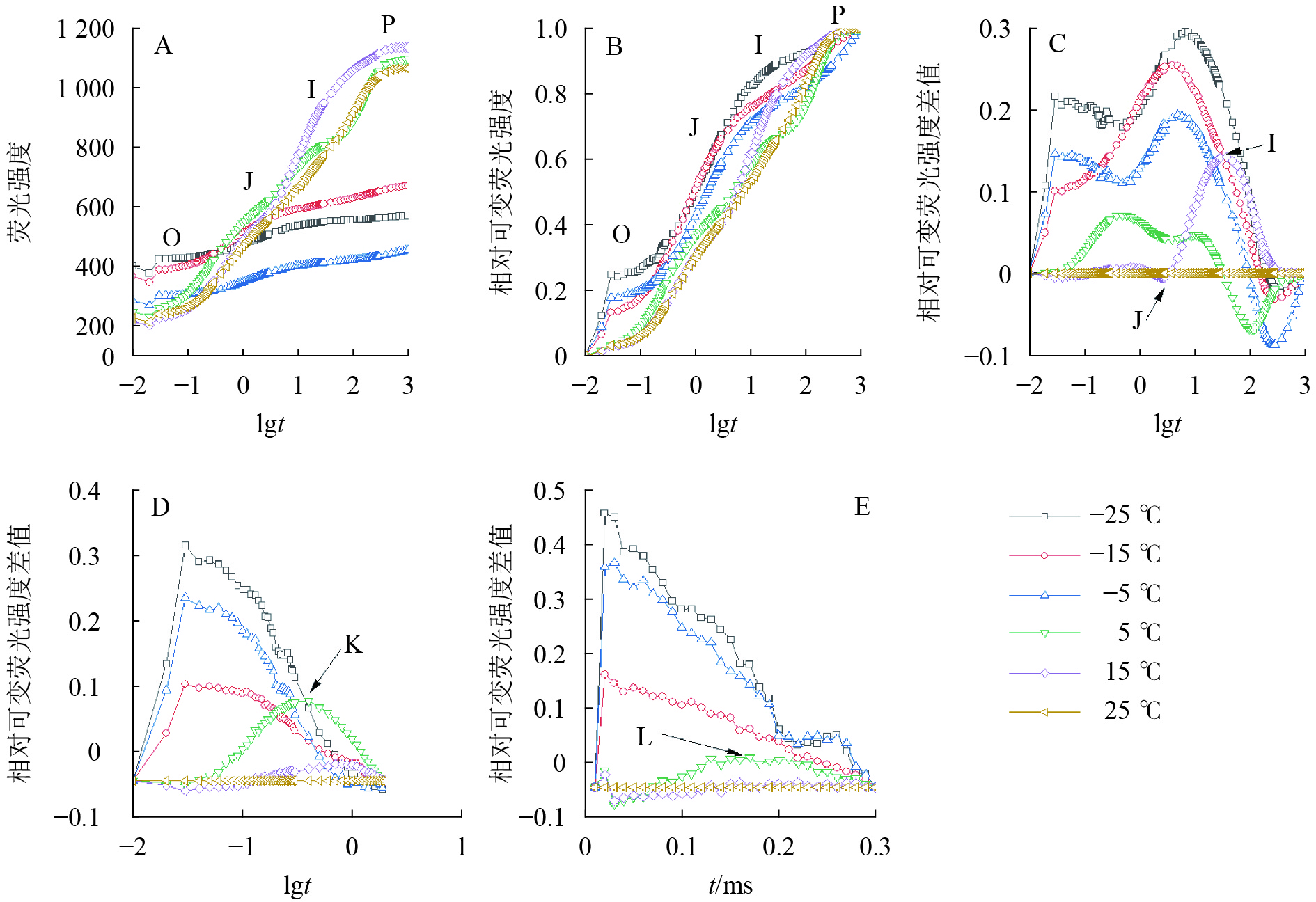
由图2可知:5、15和25 ℃处理下OJIP曲线出现O、J、I、P各相(图2A、B),表明这些温度下电子传递链运转正常;−5、−15和−25 ℃的OJIP曲线较为平缓(图2A),缺少J相和I相(图2B)。将标准化的各处理曲线与ck相减,可见J相和I相合并,并且在光照极短时间(约0.03 ms)出现1个较大峰(图2C),表明电子传递链严重受阻。OJIP曲线在5 ℃出现典型的K点(图2D)和L点(图2E),表明PSⅡ供体侧和PSⅡ反应中心受损。
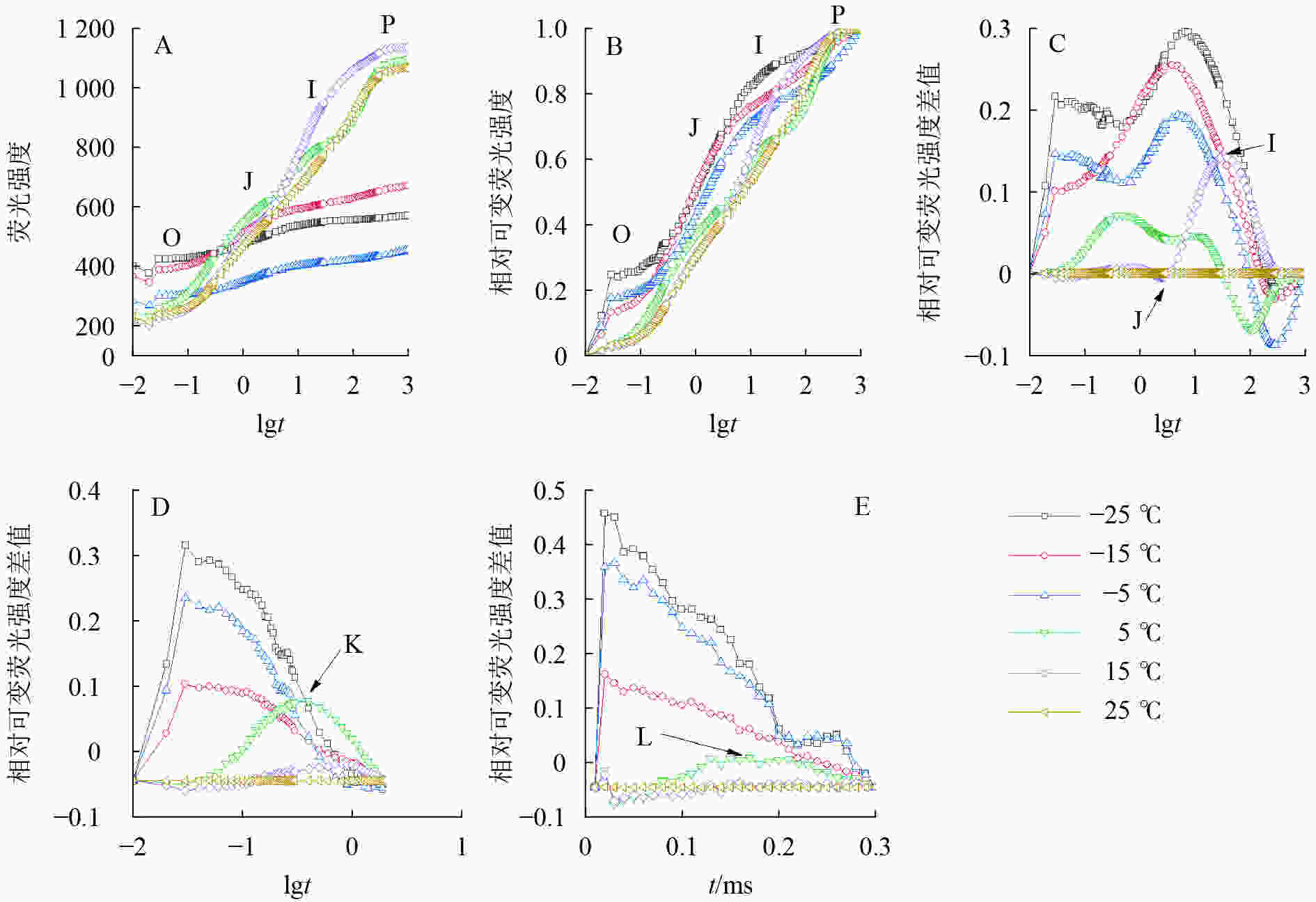
Figure 2. Chlorophyll fluorescence kinetics curve of leaves under different temperature treatments
-
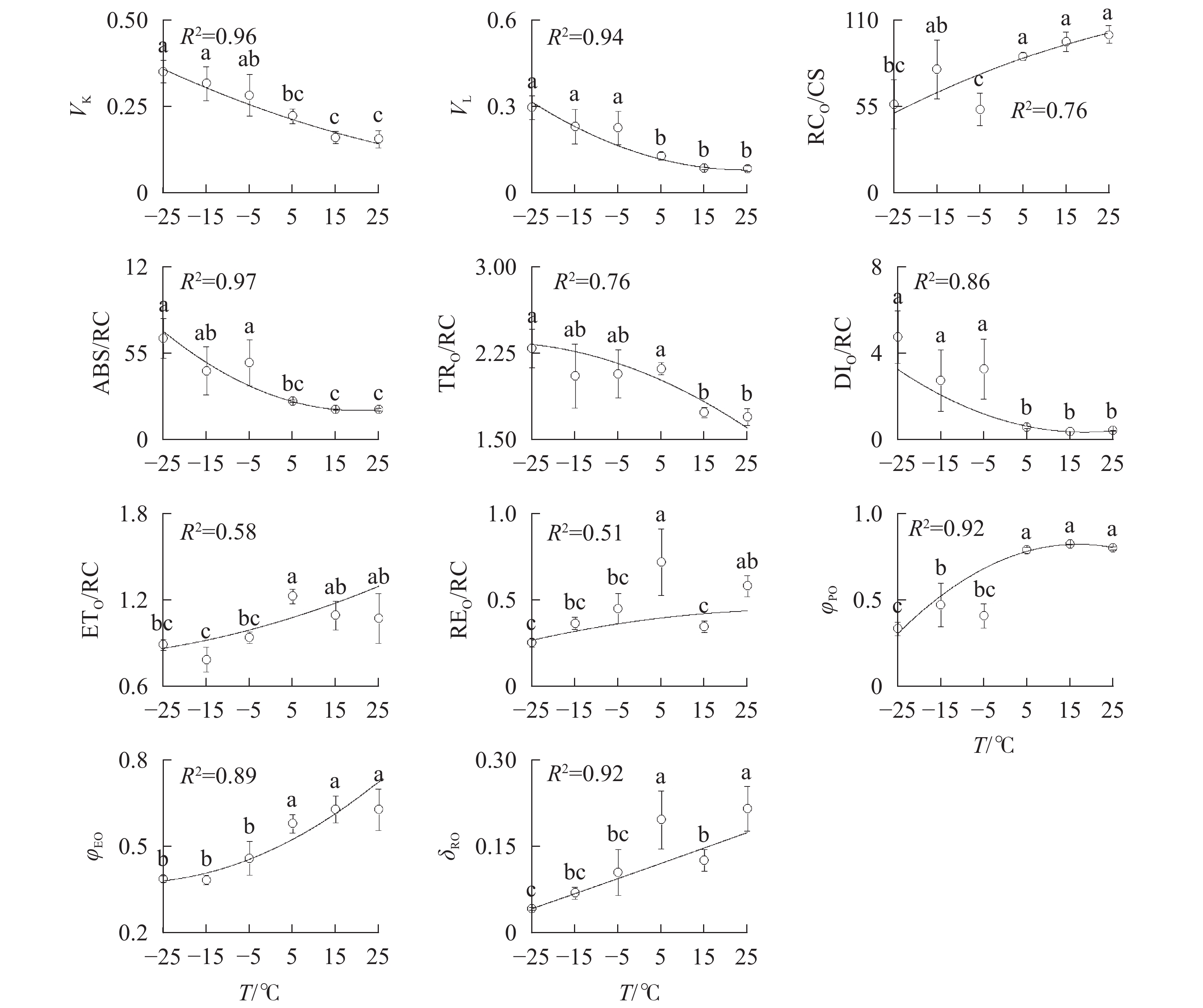
由图3A可知:VK
随着温度降低,−5、−15和−25 ℃时均与对照达到显著差异,表明≤−5 ℃的低温处理会造成PSⅡ供体侧损伤。 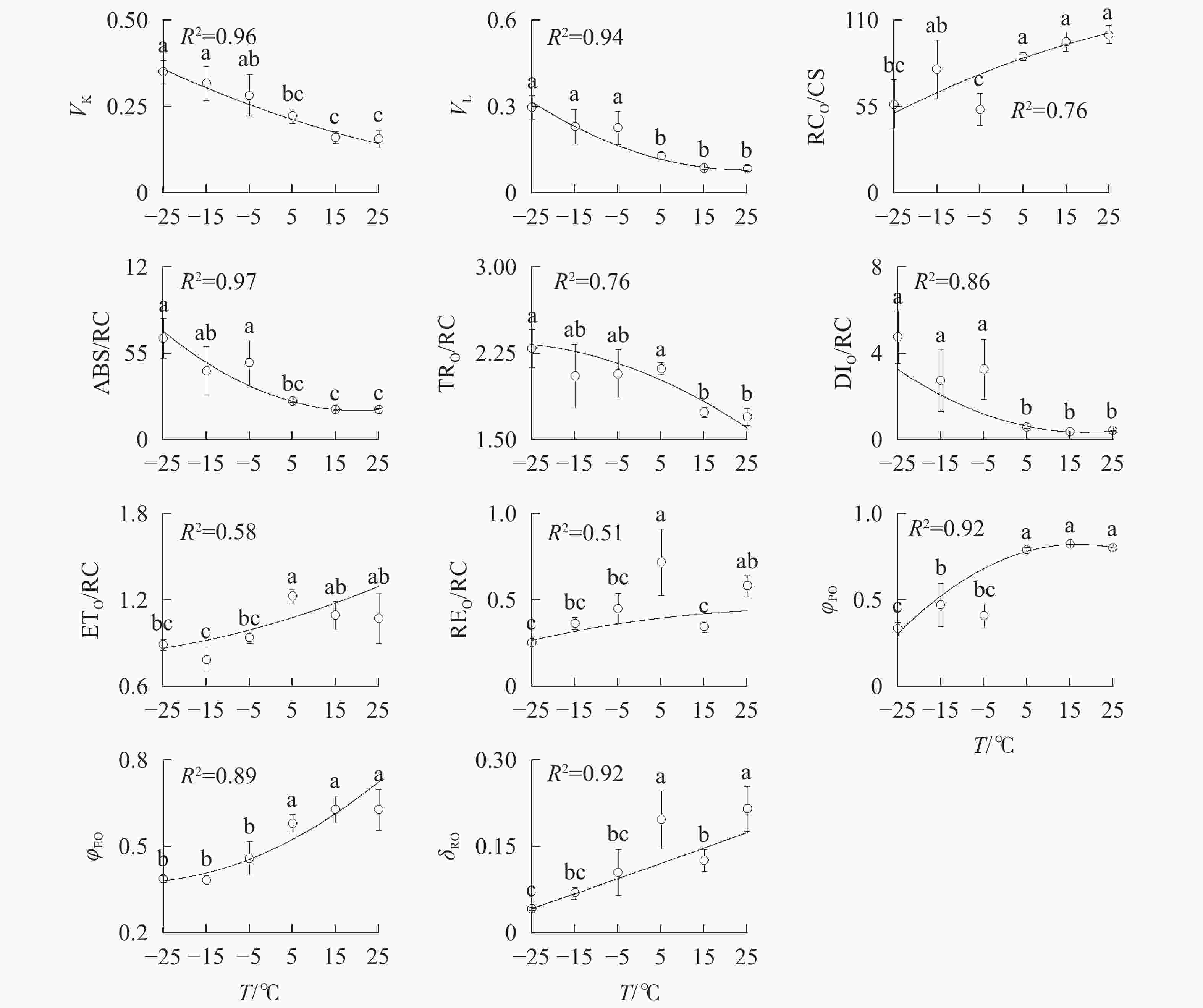
Figure 3. Parameters of chlorophyll a fluorescence transient under different temperature treatments
-
−5、−15、−25 ℃处理下VL
上升明显(图3B),表明≤−5 ℃的低温导致PSⅡ各单位之间的能量流通不畅。从−5 ℃到−25 ℃,RC/CSO随着温度降低逐渐减少(图3C),但−15 ℃的RC/CSO显著高于−5 ℃,表明−15 ℃时PSⅡ单位面积反应中心数量出现短暂回升。从−5 ℃开始,随着温度降低,ABS/RC(图3D)、TRO/RC(图3E)和DIO/RC(图3F)持续增加,−25 ℃时分别比对照增加331.0%、134.7%和1 112.7%,表明由于缺少可供还原的QA,PSⅡ单位反应中心集聚了大量能量。 -
与对照相比,ETO/RC(图3G)和REO/RC(图3H)在−15和15 ℃时短暂下降,REO/RC在−25 ℃时又显著下降,表明高于−15 ℃的温度对PSⅡ受体侧的能量传输影响较小。当温度从25 ℃降低到5 ℃时,φPO(图3I)和ψEO(图3J)与对照相比无显著差异,δRO(图3K)仅在15 ℃时出现短暂下降;当温度继续降低到−5 ℃时,3项参数均显著下降,表明≤−5 ℃的低温对PSⅡ的电子传递效率有显著负面影响。
-
随着温度降低,Fv/Fm与φPO的变化趋势相同(图4)。5 ℃时,
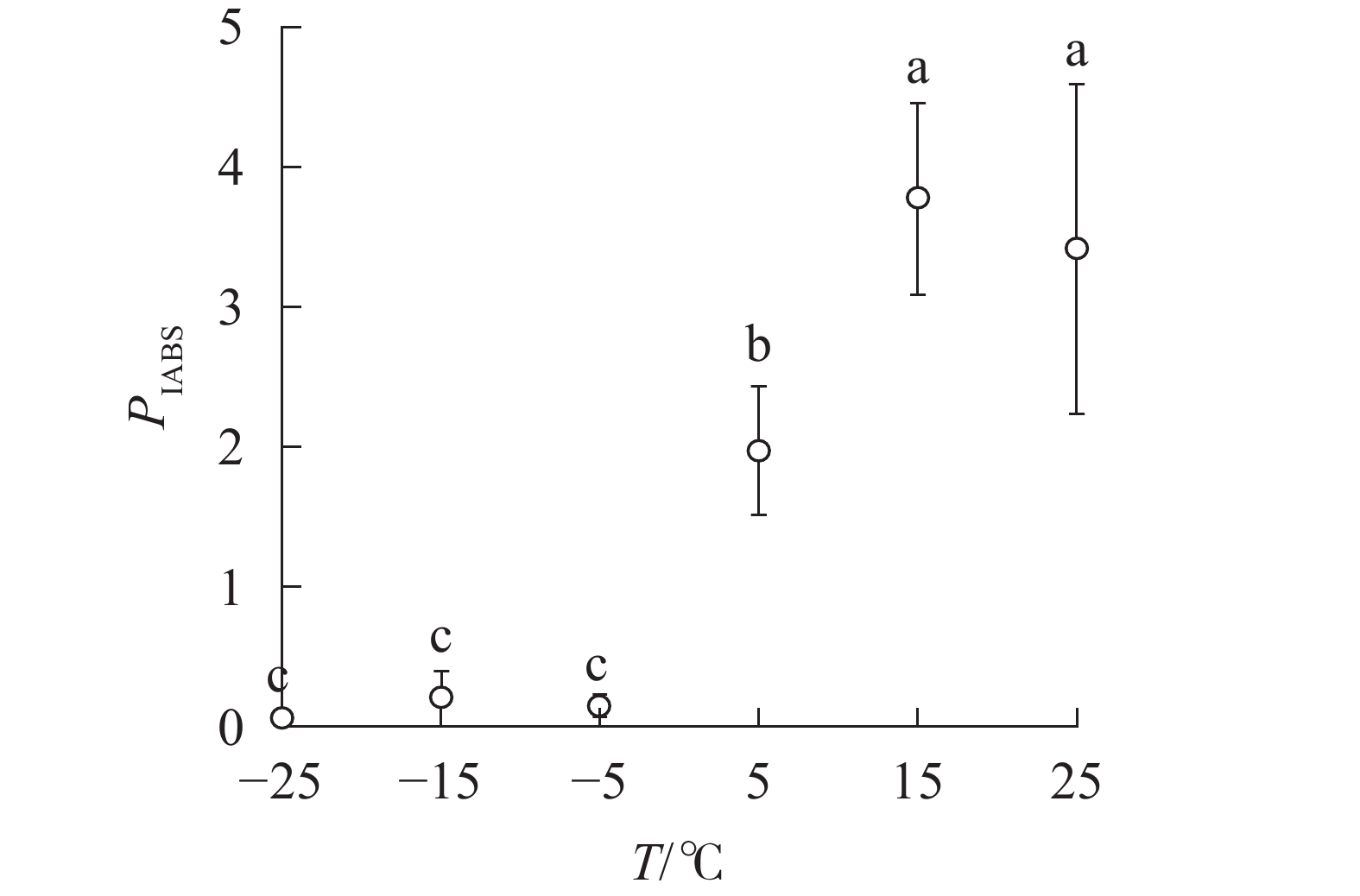
PIABS显著低于对照,−5、−15和−25 ℃时, PIABS接近于0,表明≥15 ℃的环境温度对南极假山毛榉叶片的光能吸收无显著影响,≤5 ℃时开始出现低温胁迫,≤−5 ℃会造成严重的冰冻胁迫(图5)。 
Figure 4. Fv/Fm of leaves under different temperature treatments
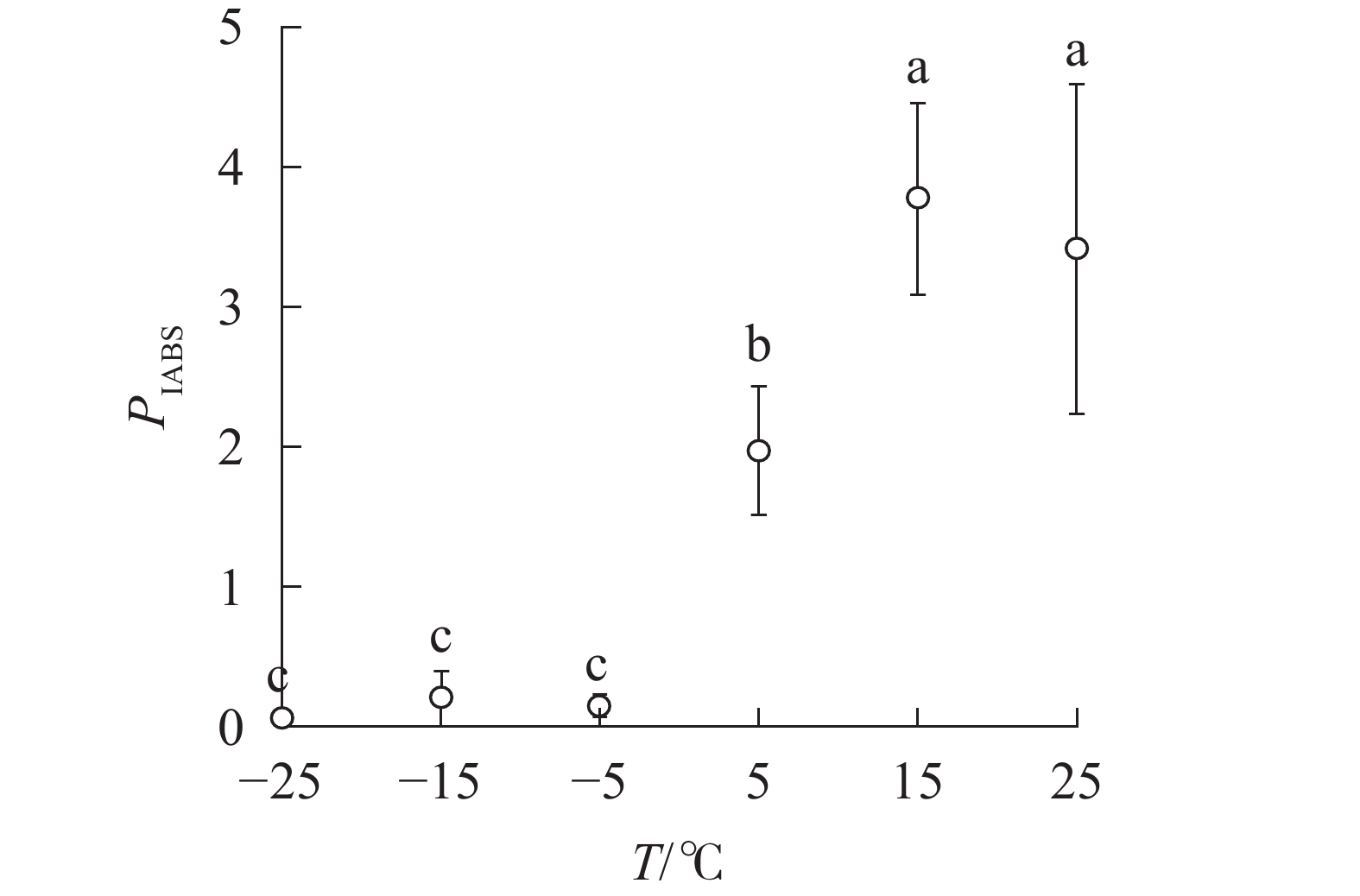
Figure 5.
PIABS of leaves under different temperature treatments -
随着温度从25 ℃降至−25 ℃,VK
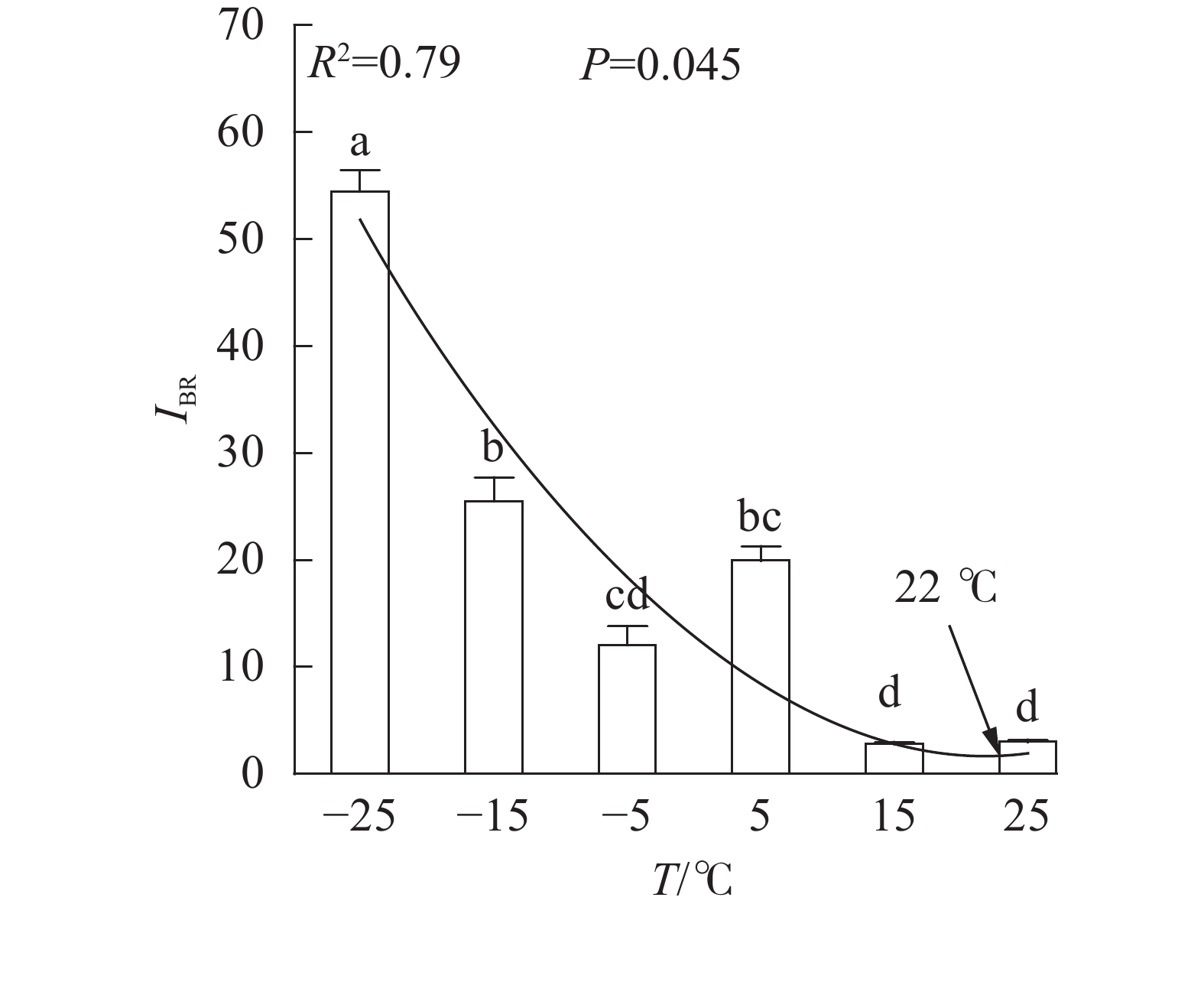
、VL、ABS/RC、DIO/RC、TRO/RC的拟合曲线持续上升,其余参数的拟合曲线持续下降(图3),因此对VL 、VK、ABS/RC、DIO/RC、TRO/RC的标准后数值取正值,其余参数取负值,绘制星状图(图6)并计算IBR(图7)。与对照相比,IBR在5、−5和−15 ℃时显著增加,−25 ℃极显著升高。对IBR作回归分析显示:22 ℃为IBR拐点,高于或低于此温度均导致IBR升高;表明IBR与温度胁迫程度呈正相关,南极假山毛榉叶片对光能的吸收利用效率最佳值在22 ℃,≤5 ℃的低温会对南极假山毛榉PSⅡ光化学活性造成胁迫。 
Figure 6. Star plots for biomarker responses under different temperature treatments
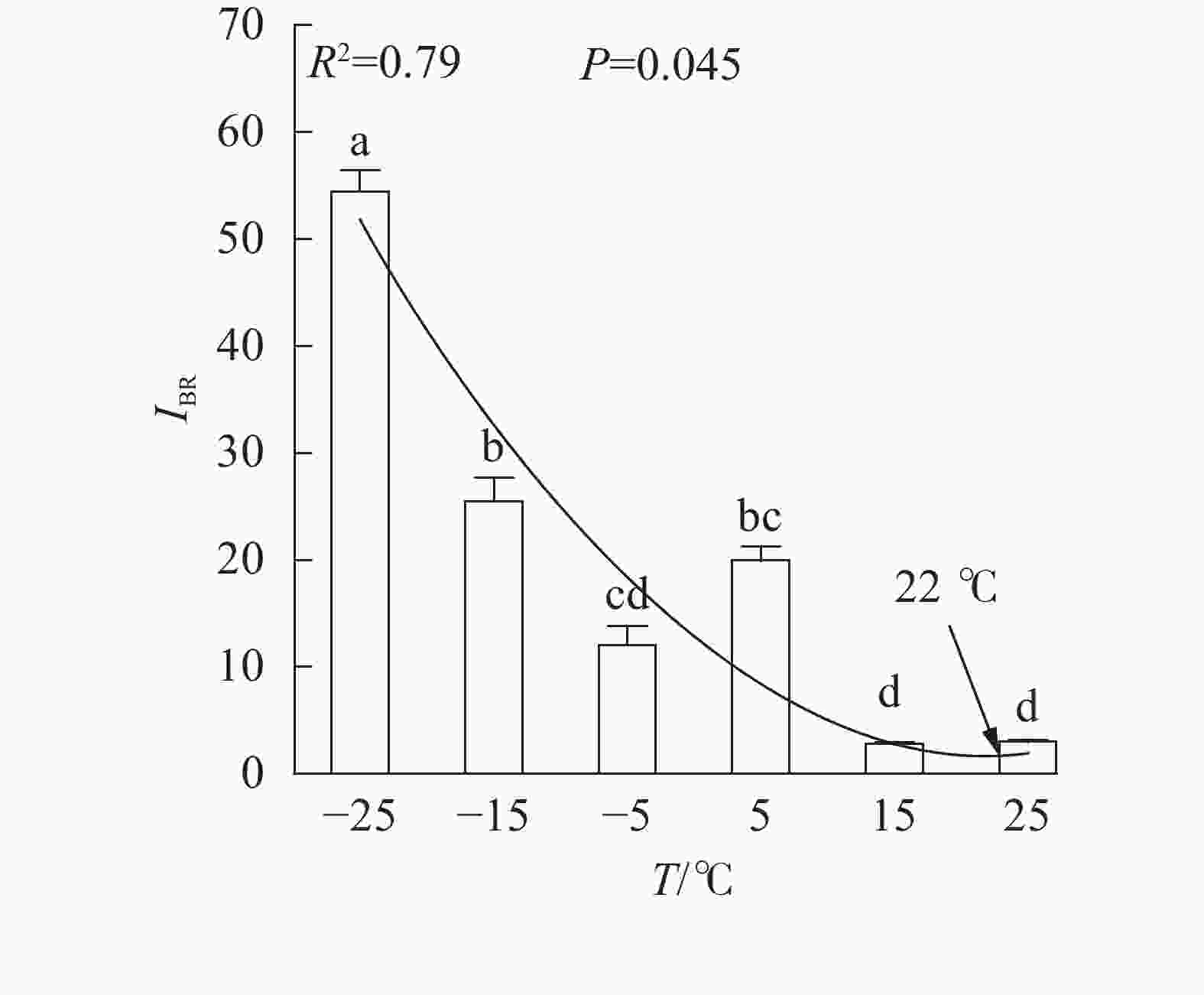
Figure 7. IBR values under different temperature treatments
-
对各温度处理下的Fv/Fm、PIABS、相对叶绿素含量、IBR和各测试温度进行相关性分析可见(表1):IBR、相对叶绿素含量和温度均为显著负相关,表明IBR能较准确反映温度对南极假山毛榉PSⅡ光化学活性以及对相对叶绿素含量的影响。Fv/Fm和PIABS虽然也与温度呈显著相关,但与相对叶绿素含量无显著相关,表明IBR与相对叶绿素含量的相关度高于上述2项参数。
参数 IBR PIABS Fv/Fm 相对叶绿
素含量测试
温度IBR 1 PIABS −0.73 1 Fv/Fm −0.72 0.94** 1 相对叶绿素含量 −0.95** 0.79 0.77 1 测试温度 −0.88* 0.90* 0.92* 0.90* 1 说明:*表示P<0.05,**表示P<0.01 Table 1. Correlation analysis of Fv
/Fm, PIABS, SPAD value, IBR and temperature
2.1. 温度对相对叶绿素含量的影响
2.2. 温度对快速叶绿素荧光诱导动力学曲线的影响
2.3. 温度对快速叶绿素荧光诱导动力学曲线参数的影响
2.3.1. 对PSⅡ供体的影响
2.3.2. 对PSⅡ反应中心的影响
2.3.3. 对PSⅡ受体侧的影响
2.3.4.
对Fv/Fm和

2.4. 温度对IBR的影响

2.5. 相关性分析
-
本研究中,当温度从25 ℃降低到15 ℃时,相对叶绿素含量、OJIP曲线、快速叶绿素荧光诱导动力学测量参数和IBR没有显著变化,说明15 ℃的环境温度对南极假山毛榉PSⅡ光化学活性的扰动轻微。当温度降低到5 ℃时,OJIP曲线出现K点和L点,PIABS开始显著下降,表明此阶段的低温主要对PSⅡ供体侧和反应中心构成胁迫,即放氧复合体(OCE)出现损伤,PSⅡ各单位之间的能量流通不畅。从−5 ℃降至−25 ℃,相对叶绿素含量显著降低,PSⅡ供体侧受损加剧;进入电子传递链的电子减少,引起电子传递效率下降;光反应的激发能不能及时传递到PSⅡ受体侧而大量积累,最终造成PSⅡ各单位之间的能量流通混乱,部分能量只能以热能的形式耗散。
OJIP曲线特征位点的变化能直观地反映出PSⅡ反应中心原初光化学反应的变化。HU等[9]发现低温处理下狗牙根Cynodon dactylon叶片OJIP曲线的相对荧光强度降低;曲丽娜等[7]进一步发现低温处理导致风箱果Physocarpus amurensis叶片的OJIP 曲线趋于平缓、P相下降、K点和L点出现;刘倩倩等[6]也得出相似结论,并发现低温处理导致O相上升。本研究发现:随着温度降低,南极假山毛榉叶片OJIP曲线的O相逐渐上升,−5、−15和−25 ℃的OJIP曲线趋于平滑,与前述研究结论相似;但前述物种均只涉及冷害胁迫(>0 ℃),而南极假山毛榉经历冻害胁迫(<0 ℃)才出现上述现象,这可能与其耐寒性强有关。−5、−15和−25 ℃处理时OJIP曲线在0.03 ms附近出现新拐点,可能因为冰冻胁迫导致PSⅡ反应中心成为能量陷阱,能吸收光能但不能推动电子传递[22],大量能量的积聚造成的。
JIP-test分析以OJIP曲线为数据基础,可提供大量反映光合器官的结构和功能的变化[23]。刘倩倩等[6]发现低温胁迫导致PSⅡ供体侧电子传递速率降低,PSⅡ反应中心数量减少;曲丽娜等[7]研究表明低温会造成放氧复合体受损,PSⅡ反应中心各单位之间的能量流通不畅,以及PSⅡ受体侧电子由初级电子受体(QA)向次级电子受体(QB)的传递过程受阻。伴随温度降低,南极假山毛榉的VK、VL、ABS/RC、TRO/RC和DIO/RC逐渐升高,RC/CSO逐渐减少,与前人研究结果相似;−15 ℃时RC/CSO出现短暂显著回升,推测是南极假山毛榉面对低温胁迫的应激反应,即通过增加PSⅡ单位面积光合机构含有的反应中心数量,以加大光能吸收量,弥补由于PSⅡ受体侧电子传递受阻而缺失的能量。
DUARTE等[20]利用IBR评价法,以JIP-test参数为数据源构建了光化学胁迫指数,证实了该参数与环境胁迫程度呈负相关关系。本研究发现IBR值与温度呈显著负相关,与DUARTE等[20]的结论相似。本研究还观察到,与Fv/Fm和
PIABS不同,IBR与相对叶绿素含量亦存在极显著负相关关系,可能预示IBR对低温胁迫的反映更具有广泛性。 多项研究显示,5 ℃的低温会引起Fv/Fm和PIABS显著降低[6-8,11];本研究发现:高于5 ℃时南极假山毛榉叶片的Fv/Fm和IBR无显著变化,高于15 ℃时PIABS无显著变化,表明南极假山毛榉叶片PSⅡ的温度适应性强,这可能是南极假山毛榉耐寒性强的生理基础之一。研究还发现:冰冻胁迫对PSⅡ的光吸收能力抑制明显,预示在南极假山毛榉的生长季需要预防零度以下的寒潮侵袭。STECCONI等[24]发现南极假山毛榉的最适生长温度为18.5 ℃/7.0 ℃(昼/夜),回归分析显示IBR的最小值出现在22 ℃,这一温度与上述研究结果仍有差距,可能与低温处理时间,以及参与计算IBR的参数种类有关,还需进一步探究。
 DownLoad:
DownLoad: