




-
同一植物长期生长在异质环境条件下,经过自然选择和适应过程,在形态结构与生理代谢方面会发生分化,从而表现出丰富的表型可塑性[1-3]及生理生态特征变异[4-6],这反映了种群应对不同生境条件的进化方式和适应对策[7-8]。为了区分同一植物不同种源的性状差异是源自遗传效应还是环境效应,通常将相同种源的植物移植至不同环境下来研究环境效应,而将不同种源的植物移植至相同环境下来研究遗传效应[9]。此外,母体植株经受的环境胁迫也会显著影响其子代的生长发育和生理特性[10],但这种母体效应的影响程度和机理还有待深入研究。目前,关于不同地理种群/种源植物的光合特性方面研究已有大量报道[11-21]。KUNDU等[11]比较研究了印楝Azadirachta indica在南亚和非洲10个种源间的光合生理特征,发现其净光合速率、气孔导度、气孔密度、保卫细胞长度等在不同种源间均存在显著差异,印楝各种源早期的生物量积累与光合作用及气孔特征密切相关。黄秦军等[12]研究表明:5个种源蒙古栎Quercus mongolica幼苗的生长生理特性均存在差异,江密峰种源的生物量显著高于其他4个种源,同时单叶面积、栅栏组织厚度、光合色素含量及净光合速率也优于其他种源。熊彩云等[13]基于净光合速率、最大光合速率、光补偿点、光饱和点等光合作用参数,评价了3个不同纬度的木荷Schima superba种源对环境的适应能力,认为南部开平种源对当地环境具有较强的适应能力和生长潜力。张玲菊等[14]测定分析了浙江和云南5个种源铁皮石斛Dendrobium officinale的光合速率及光响应曲线,表明铁皮石斛不同种源的光合特性均表现出阴生植物特点,对光照的需求不强,其中云南广南种源的光照生态幅相对更宽。由此可见,植物在不同地理种源间通过改变自身光合生理特性来更好地适应新的环境。然而,针对极端“干热”环境的干热河谷分布区,关于牛角瓜Calotropis gigantea不同种源的光合能力及水分利用研究尚未见报道。牛角瓜原产于亚洲和非洲南部,后被引入南美洲和加勒比海一带的岛屿[22]。现广泛分布于亚洲和非洲热带及亚热带地区,是印度、印度尼西亚、马来西亚、泰国、尼泊尔、斯里兰卡、中国的特有种,在中国主要分布于云南、海南、广东、四川等省[23]。牛角瓜在干热河谷的荒山野坡、旷野瘠地、江滩河岸等严酷生境下生长迅速,长势良好,花果期全年,具有改善干热河谷脆弱生境与防治水土流失的生态功能[23]。同时,近年来牛角瓜被发现具有药用[24-27]、纤维[22, 28-32]、能源[33-34]等多种用途的资源开发价值,是不可多得的生态经济型植物。尽管如此,由于牛角瓜的自然分布较为分散,且尚无系统的集约化栽培管理技术,加之牛角瓜在中国还未受到足够的重视,导致其生态经济价值的推广受到极大限制。为此,本研究比较分析了干热河谷牛角瓜不同种源光合生理参数的日变化动态与日均值差异,筛选出干热河谷牛角瓜的优良种质资源,旨在为牛角瓜的定向培育及开发利用提供理论依据。
HTML
-
研究地位于云南省元江县种苗站,地处23°19′~23°55′N,101°39′~102°22′E,海拔390~420 m,属典型元江干热河谷气候区。年平均气温为23.7 ℃,最高日平均气温28.8 ℃,≥10 ℃的年积温4 000.0~8 700.0 ℃,无霜期200.0~364.0 d,年平均降水量787.0 mm[35]。
于2014年4月进行不同种源牛角瓜的播种育苗,供试种子采自元江河谷区内的元江、红河、个旧、建水、元阳等典型代表地域自然分布的野生牛角瓜成熟果实。牛角瓜不同种源的地理位置和生境条件见表 1。播种200粒·种源-1,种子播在10 cm × 15 cm的营养袋中,土壤基质为燥红土,育苗期间常规水肥管理。于2014年8月,选择每个种源正常生长、长势一致的盆栽幼苗12株(1株·盆-1),开展牛角瓜苗期光合生理特性的种源试验。
种源 种源地点 海拔/m 地理坐标 树高/m 地径/cm 冠幅/m 结果数/个 主要植被 分布位置 元江 元江那塘 608 23°31'14.2"N 102°04'41.0"E 1.9 4.5 1.5 14 合欢、酸角、香蕉 山脚公路旁 红河 红河乌弯寨 467 23°17'37.5"N 102°37'30.9"E 2.6 7.0 3.1 32 车桑子 山腰公路旁 个旧 个旧老新寨 255 23°10'2.3"N 102°59'48.5"E 2.5 6.0 3.3 38 榕树、檻木 山腰公路旁 建水 建水自营矿山 234 23°12'49.4"N 102°53'29.5"E 2.5 8.0 2.8 35 膏桐、芒果 山腰公路旁 元阳 元阳南沙-蛮柏沙路 222 23°12'43.6"N 102°52'30.9"E 1.6 5.0 1.6 15 车桑子 江边 说明:合欢Albizia julibrissin,酸角Tamarindus indica,香蕉Musa paradisiaca,车桑子Dodonaea viscosa,榕树Ficus microcarpa,檵木Loropetalum chinensis,膏桐Jatropha carcas,芒果Mangifera indica。 Table 1. Geographical locations and habitat conditions of Calotropis gigantea from different provenances
-
采用便携式CO2/H2O分析系统(Li-Cor 6400; Li-Cor Inc.,Lincoln,美国),于2014年8月中上旬连续晴朗天气条件下,选待测植株顶端第5,6,7枚健康成熟的叶片(3次重复),各测定日从8:00-18:00隔2 h原位测量牛角瓜不同种源幼苗的气体交换进程。净光合速率(Pn),气孔导度(Gs),胞间二氧化碳摩尔分数(Ci),蒸腾速率(Tr),空气温度(Ta),光合有效辐射(PAR),空气二氧化碳摩尔分数(Ca)由Li-6400光合作用系统同步自动记录。水分利用效率(WUE)=Pn/Tr,光能利用效率(LUE)=(Pn/PAR)×1 000,瞬时羧化效率(CE)=Pn/Ci,气孔限制值(ls)=1-Ci/Ca[36-37]。
将牛角瓜的光合生理参数在日变化进程中每个时刻的实测值加以平均,得到各光合生理参数的日均值并比较分析它们在不同种源间的差异。
-
数据的统计分析通过SPSS 13.0软件(SPSS Inc. 美国)完成,文中图形在Microsoft Office Excel 2003软件中完成。图表中所有数据均由平均值±标准误表示。方差分析采用多重比较分析中的邓肯检验(Duncan’s test),不同字母表示在P<0.05水平差异显著。
1.1. 研究地概况
1.2. 气体交换测量
1.3. 统计分析
-
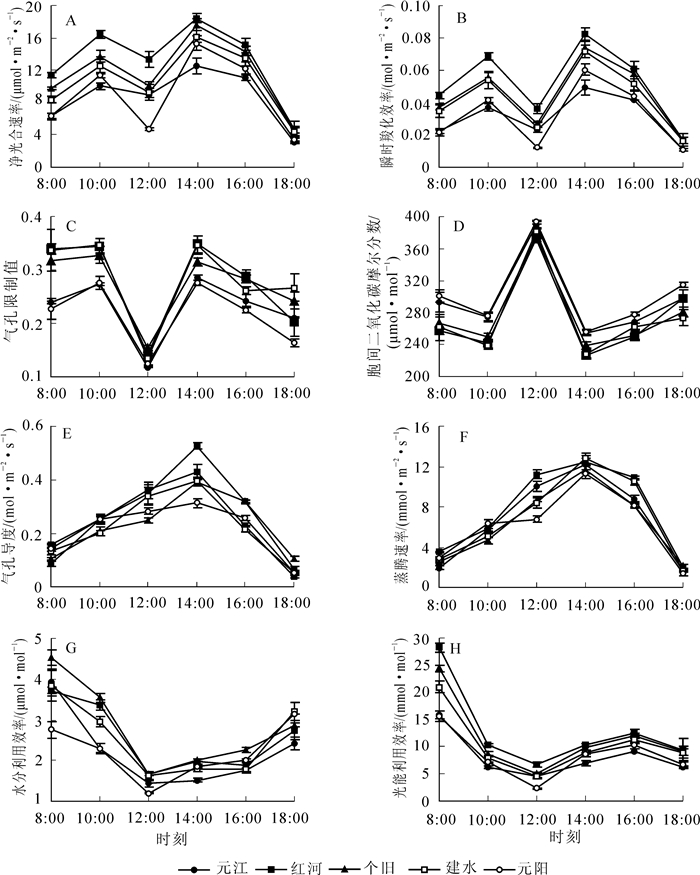
为了了解牛角瓜不同种源在育苗期的光合生理特性,分析了牛角瓜幼苗净光合速率、气孔导度、胞间二氧化碳摩尔分数、蒸腾速率、水分利用效率、光能利用效率、瞬时羧化效率和气孔限制值等在元江、红河、个旧、建水、元阳等种源间的日变化动态(图 1)。牛角瓜5个种源的净光合速率日变化趋势近似一致,均呈典型的双峰“M型”曲线,但在不同时刻的数值存在种源间的差异,其2次峰值分别出现在10:00和14:00,第2次峰值高于第1次,且在不同种源间均表现为:红河>个旧>建水>元阳>元江,而在12:00具有明显的“午休”现象,其中尤以元阳种源降幅最大(图 1A)。

Figure 1. Diurnal dynamics of photosynthetic physiology characteristics of Calotropis gigantea from different provenances
牛角瓜不同种源的瞬时羧化效率和气孔限制值日变化曲线与净光合速率基本相同,大致呈“M型”,峰值、谷值出现时间与净光合速率一致(图 1B,C),而胞间二氧化碳摩尔分数则呈现“W型”的相反趋势,于12:00达到全日最大值(图 1D)。根据气孔限制值和胞间二氧化碳摩尔分数的变化方向判别牛角瓜净光合速率下降的限制性因素:当净光合速率下降时,若气孔限制值增大且胞间二氧化碳摩尔分数降低,则为气孔限制;若气孔限制值减小且二氧化碳摩尔分数增大或不变,则为非气孔限制。因此,牛角瓜净光合速率在中午和下午的2次下降是由于非气孔限制所致。
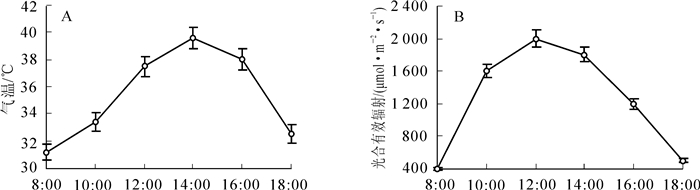
牛角瓜不同种源的气孔导度和蒸腾速率日变化规律表现为典型的单峰曲线,均于14:00升至全日最大峰值(图 1E,F),与干热河谷气温的日变化动态趋于一致(图 2A)。此时气温高达40.0 ℃,12:00-16:00也一直维持在38.0 ℃以上,而牛角瓜的净光合速率、气孔导度、蒸腾速率、瞬时羧化效率在14:00达到最高水平,表明干热河谷5个种源的牛角瓜在育苗期对高温具有极强的适应性,通过增加气孔导度和蒸腾作用来提高光合同化效率。因此,干热河谷的极端高温不是影响牛角瓜幼苗光合作用与羧化反应的限制因素。

Figure 2. Diurnal dynamics of (A) Ta and (B) PAR in dry-hot valley
牛角瓜水分利用效率和光能利用效率日进程呈现近似“V型”曲线,均于8:00达最大值,8:00-12:00呈快速下降趋势,12:00之后呈逐渐上升趋势,仅光能利用效率在16:00以后轻微降低(图 1G,H)。牛角瓜的净光合速率、水分利用效率、光能利用效率、瞬时羧化效率在午间急剧下降,主要是由于此时的环境光合有效辐射达到全日最大值(约2 000·mol·m-2·s-1)。由此可见,干热河谷正午的强光辐射显著抑制了牛角瓜幼苗的光合作用与水分利用。
-
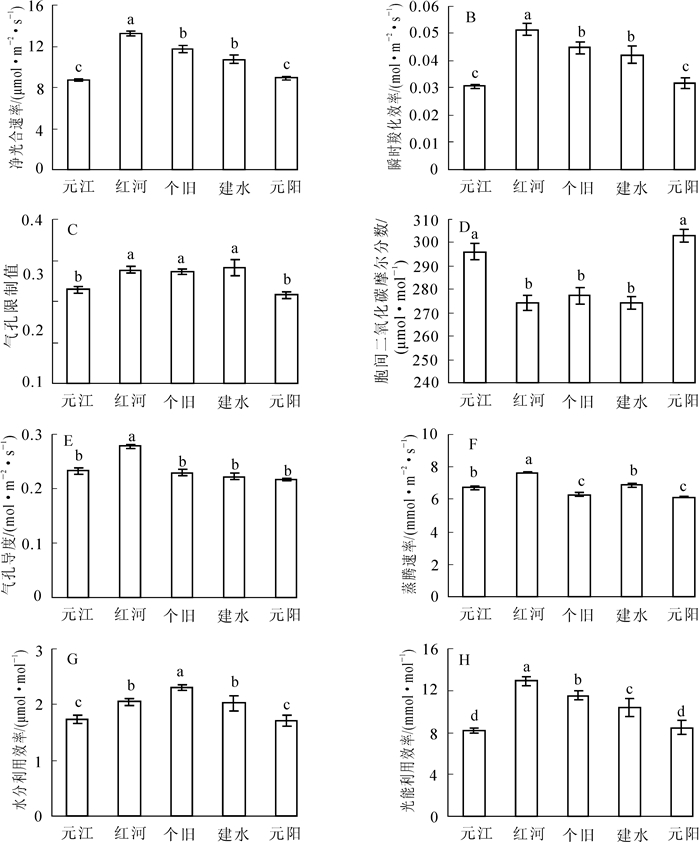
为了进一步弄清牛角瓜苗期光合生理特性的种源间差异,比较了牛角瓜5个种源幼苗的光合生理参数日均值(图 3)。总体上看,牛角瓜的净光合速率、气孔导度、胞间二氧化碳摩尔分数、蒸腾速率、水分利用效率、光能利用效率、瞬时羧化效率、气孔限制值日均值在不同种源间均具显著差异(P<0.05)。其中,净光合速率、瞬时羧化效率、水分利用效率、光能利用效率等关键参数的变化幅度分别为8.67~13.22 μmol·m-2·s-1(图 3A),0.03~0.05 mol·m-2·s-1(图 3B),1.71~2.31 μmol·mmol-1(图 3G),8.18~12.91 μmol·mmol-1(图 3H),表明牛角瓜苗期不同种源间的光合生理特性及水分利用能力具有较大差别。

Figure 3. Comparisons in daily mean values of photosynthetic physiology characteristics of Calotropis gigantea from different provenances
综合比较牛角瓜5个种源的主要光合生理参数可知,红河种源的净光合速率、蒸腾速率、光能利用效率和羧化效率显著高于其他4个种源(P<0.05);个旧种源表现出“高光合、低蒸腾”的显著特点,从而具有最高的水分利用效率,分别比红河、建水、元江、元阳种源高出12.68%,13.79%,34.30%,35.09%,其净光合速率、光能利用效率和羧化效率也维持在较高水平;而建水、元江、元阳种源的光合生理活性相对较弱。因此,筛选出红河、个旧为具有高水平光合生产与水分利用潜力的干热河谷牛角瓜种质资源。
2.1. 牛角瓜不同种源光合生理特性的日变化动态
2.2. 牛角瓜不同种源光合生理参数的日均值对比
-
项目组前期调查分析了元江干热河谷野生牛角瓜不同种源的18个果实性状指标,发现其变异范围为3.99%~42.84%,表明干热河谷区内牛角瓜果实在种源间存在广泛的遗传变异性。基于果实品质性状和棉纺经济性状,初步筛选出红河、个旧为干热河谷牛角瓜不同地理分布的优良种源[38]。此外,在牛角瓜不同种源子代的育苗期,观察发现红河、个旧种源的幼苗生长势优于元江、建水、元阳种源。本研究进一步证明红河、个旧种源的幼苗光合生理活性更高,说明在牛角瓜苗期从光合生理水平上能够揭示其生长表现出的种源间差异,而且本研究结果与前期的牛角瓜野外种质资源调查及果实性状分析结果基本一致。
在牛角瓜的光合特性方面,高柱等[39]研究了2年生牛角瓜在红河河谷的光合日进程。结果表明:牛角瓜的净光合速率、气孔导度、蒸腾速率呈现先升后降趋势,并未出现光合“午休”现象。牛角瓜的净光合速率与光合有效辐射相关性极显著,而与气温相关性不显著,气温不是直接限制牛角瓜光合作用的因素,表明牛角瓜作为干热河谷典型阳性植物,对高温、高光具有较强的耐性。本研究中,5个种源牛角瓜苗期净光合速率的日变化动态均呈双峰曲线,有明显的“午休”现象,与上述研究结果不同,但与前人对不同种源铁皮石斛幼苗[14]、青钱柳Cyclocarya paliurus幼苗[19]的研究结论相似。分析认为,导致牛角瓜在苗期发生光合“午休”主要是因为正午更大程度的强光抑制引起光合速率下降,而非由于气温过高导致酶钝化或水分胁迫造成气孔部分关闭。即便如此,红河种源的牛角瓜幼苗在午间较低的净光合速率(13.37 μmol·m-2·s-1)仍与牛角瓜幼树在全日最高的Pn(13.67 μmol·m-2·s-1)水平相当,表现出更高的光合能力。
气温在很大程度上决定了光合作用的生化反应速度和光合酶活性。同一种植物生长在气温更高的地区,其温度生态位的范围往往更大,光合最适温度也会更高[21]。通常光合作用关键酶1,5-二磷酸核酮糖羧化/加氧酶Rubisco的最适活化温度在25~30 ℃,叶温过高会抑制Rubisco的羧化活性而催化加氧反应,导致光呼吸速率增高,不利于光合作用的进行[40-43]。本研究发现,5个种源牛角瓜的光合最适温度更高,净光合速率最大值出现的温度范围为33~40 ℃,它们在强烈高温条件下通过增加气孔导度和蒸腾作用,来提高光合速率和羧化效率,从而增强它们对干热河谷高温环境的适应能力。
干热河谷牛角瓜不同种源间的光合生理特性存在显著差异。综合分析净光合速率、瞬时羧化效率、水分利用效率、光能利用效率等主要光合生理参数,本研究筛选出红河、个旧为具有高水平光合生产与水分利用潜力的干热河谷牛角瓜种质资源。今后将继续扩大牛角瓜的种源收集范围,进一步对比研究中国云南、海南、广东、四川和泰国等地理种源的生长及生理特性,以便在不同地域环境条件下筛选出合适的牛角瓜优良种源。
 DownLoad:
DownLoad: