




-
干热河谷在中国主要分布于西南横断山脉的金沙江、怒江、澜沧江和红河流域的高山峡谷区,其主要环境特点是热量高、降水少且蒸发量大,存在季节性干旱。干热河谷自然植被中乔灌层发育差,多以扭黄茅Heteropogon contortus等禾本科Poaceae植物为背景,其间分布少量低矮乔灌木树种。植被覆盖率低、土壤发育差、水土流失严重[1-3]。人类活动(樵取、火烧、放牧等)也加剧了此类问题[4-5]。因此,在干热河谷地区如何进行以保护物种多样性为目标的植被恢复工作就显得尤为重要。在生态系统中,生物多样性既反映了物种的分布及其对环境的适应情况,也反映了群落的物种组成、结构及变化趋势[6-7]。物种多样性作为生物多样性在物种水平上的表现形式,在很大程度上决定了群落稳定性,它是生物多样性的核心,也是认识森林生态系统结构和功能的基础[8-10]。目前,已有部分学者对干热河谷地区不同植被类型的群落特征[11-12]、多样性[13-14]及植被恢复[2-3, 15]等进行了研究,但对红河干热河谷区不同恢复群落与林下物种组成、多样性特征及土壤理化性质之间的关系研究还需要进一步完善。基于此,本研究以4种人工恢复群落和天然次生稀树灌草丛为研究对象,分析不同人工恢复群落与天然次生稀树灌草丛在物种组成及多样性方面的差异,探究不同人工恢复群落对林下植物多样性与土壤理化性质的影响,以期为该地区的植被恢复提供科学参考和理论依据。
-
研究区域位于云南红河县境内干热河谷区(23°19′38″~23°24′55″N,102°19′38″~102°25′36″E),地处云南省东南部,海拔为400~1 000 m,以干热为主要特点,旱、雨季分明。年平均气温为20.3 ℃,年降水量为865.2 mm,年平均干燥度为1.5~2.0,属半干旱偏湿亚类型。土壤以红壤和燥红土为主。选取久树Schleichera oleosa群落、车桑子Dodonaea viscosa群落、铁刀木Cassia siamea群落、清香木Pistacia weinmannifolia群落等4种人工恢复群落和天然次生稀树灌草丛为研究对象,其中,人工恢复群落于2003年采用百日袋装苗,在稀树灌草丛退化区域营造,密度为1 600~3 300株·hm−2。营造前,沿等高线开挖带状水平台地并且松土,带宽≥30 cm,带间距3 m,定植穴为1 m×1 m。稀树灌草丛中优势种为虾子花Woodfordia fruticosa和扭黄茅,是红河干热河谷区典型的天然次生植被之一。
-
在4种人工恢复群落及稀树灌草丛中各设置6个10 m×10 m的样地,在每个样地中各设置4个1 m×1 m的样方调查草本层植物。对每个10 m×10 m样地内树高1.5 m以上的植株进行每木检尺(包括1.3 m以下分枝的乔木),记录物种名称、胸径(灌木记录株丛数)、枝下高(灌木记录灌高)、冠幅等指标,在120个1 m×1 m的样方中,记录草本层植物种名、株(丛)数、平均高度及盖度等。不同样地基本情况见表1。
表 1 样地基本情况
Table 1. Basic information of sample plots
样地 海拔/
m平均胸
径/cm枝下高/
m平均冠幅/
m林分密度/
(株·hm−2)久树群落 530 5.06 1.58 3.32 2 867 车桑子群落 675 3.02 1.94 1.81 1 650 铁刀木群落 520 3.63 0.82 2.85 2 017 清香木群落 850 4.73 0.77 4.72 3 267 稀树灌草丛 702 1.82 1.84 2.03 1 200 在每个10 m×10 m的样地中,沿对角线随机选择6个取样点,用土壤采样器(内径为5 cm)采集土样(0~20 cm土层),形成混合样,除去植物残体后,土样风干,于4 ℃下保存。
-
以每个样地为单位分别计算乔木、灌木和草本植物的重要值。计算公式如下:乔木重要值=(相对多度+相对显著度+相对频度)/3×100,灌木和草本重要值=(相对多度+相对频度+相对盖度)/3×100[16-17]。
-
采用丰富度指数(R=S)、Shannon-Wiener多样性指数
$\left( H=- \displaystyle \sum\limits_{i=1}^{S}{P}_{i}\mathrm{lg}{P}_{i} \right)$ 、Simpson优势度指数$\left( D=1- \displaystyle \sum\limits_{i=1}^{S}{P}_{i}^{2} \right)$ 及Pielou均匀度指数(J=$ H/\mathrm{l}\mathrm{g}S $ )对群落多样性进行评价。计算式中:S是样地内的物种总数,Pi为种i在样地内的重要值[18-19]。 -
Jaccard相似系数(SIj):SIj=j/(A+B−j)。其中:A为样方A中种的总数,B为样方B中种的总数,j为A和B样方中共有物种数。根据Jaccard相似性原理,当SIj为0~0.25时为极不相似,当SIj为0.25~0.50时为中等不相似,当SIj为0.50~0.75时为中等相似,当SIj为0.75~1.00时为极相似[20]。
-
土壤容重和含水量等物理性质采用环刀法测定。土壤总孔隙度=(1−容重/比重)×100%。土壤化学性质测定:采用NY/T 1377—2007《土壤pH的测定》测定土壤pH;采用NY/T 1121.6—2006《土壤检测 第6部分:土壤有机质的测定》测定土壤有机质;采用LY/T 1228—2015《森林土壤氮的测定》测定土壤氮;采用NY/T 1121.7—2014《土壤检测 第7部分:土壤有效磷的测定》测定土壤有效磷;采用NY/T 889—2004《土壤速效钾和缓效钾含量的测定》测定土壤速效钾。
-
采用SPSS 24.0和Excel 2010完成。
-
在5种处理的林下植被中,共有60种植物,隶属于20科49属。植物种类较多的科为豆科Leguminosae、菊科Compositae、禾本科和大戟科Euphorbiaceae,共占林下所有植物属的61.22%和所有植物种类的61.67%,林下植物中只含1个物种的属较多,占所有属的81.63%。在稀树灌草丛中发现有23种植物,占林下所有植物种类的38.30%,其中金合欢Acacia farnesiana、小蓟Cirsium setosum、矛叶荩草Arthraxon lanceolatus、水蔗草Apluda mutica、土丁桂Evolvulus alsinoides、野拔子Elsholtzia rugulosa和独脚金Striga asiatica等7种植物仅在稀树灌草丛中有分布。
在科水平上,不同植物群落中物种分布特征存在一定差异。豆科植物在清香木群落中分布最多,占林下植物种类的16.67%,其次是铁刀木群落,占林下植物种类的13.30%;菊科植物在稀树灌草丛中分布最多,占林下植物种类的10.00%;禾本科植物在清香木群落中分布最多,占林下植物种类的6.67%;大戟科植物在久树群落、清香木群落、铁刀木群落中分布特征一致,占林下植物种类的5.00%。
-
不同植物群落中,扭黄茅的重要值从大到小依次为铁刀木群落(51.74)、车桑子群落(42.77)、稀树灌草丛(40.81)、久树群落(37.33)和清香木群落(29.87),且在各恢复群落中均为最大,表明扭黄茅在干热河谷地区的林下植被中处于重要的地位,具有极强的竞争力。除扭黄茅、车桑子以外,不同群落中重要值处于前10位的植物种类不完全相同(表2),表明不同人工恢复群落中,各种植物在群落中找到了新的生态位,群落也朝着更稳定的方向演替。
表 2 各群落中重要值位于前10位的物种
Table 2. Species with top 10 important values in each community
序号 稀树灌草丛 车桑子群落 久树群落 清香木群落 铁刀木群落 植物 重要值 植物 重要值 植物 重要值 植物 重要值 植物 重要值 1 扭黄茅 40.81 扭黄茅 42.77 扭黄茅 37.33 扭黄茅 29.87 扭黄茅 51.74 2 水蔗草 18.34 独穗飘拂草 19.37 狸尾豆 15.90 耳草 18.11 蔓草虫豆 9.57 3 独穗飘拂草 8.77 车桑子 6.67 飞机草 12.15 藿香蓟 11.08 三点金 7.69 4 蔓草虫豆 4.20 三点金 5.81 迎春花 7.52 飞机草 6.92 车桑子 3.22 5 矛叶荩草 3.70 飞机草 4.01 华西小石积 4.07 蔓草虫豆 4.86 猪屎豆 2.65 6 野拔子 3.11 蔓草虫豆 3.11 单叶拿身草 4.06 链荚豆 3.31 羽芒菊 2.38 7 椭圆叶木蓝 2.76 黄珠子草 3.10 车桑子 3.42 三点金 3.21 黄珠子草 2.26 8 白花叶 2.76 响铃豆 2.22 久树 2.04 铁刀木 3.11 鬼针草 1.98 9 车桑子 2.73 狸尾豆 1.79 三点金 1.74 黄珠子草 2.44 链荚豆 1.87 10 狸尾豆 1.88 羽芒菊 1.47 椭圆叶木蓝 1.48 车桑子 2.19 白羊草 1.81 说明:独穗飘拂草Fimbristylis monostachya,狸尾豆Uraria lagopodioides,耳草Hedyotis auricularia,蔓草虫豆Cajanus scarabaeoides, 飞机草Eupatorium odoratum,藿香蓟Ageratum conyzoides,三点金Desmodium triflorum,迎春花Jasminum nudiflorum,矛叶荩 草Arthraxon lanceolatus,华西小石积Osteomeles schwerinae,猪屎豆Crotalaria pallida,单叶拿身草Desmodium zonatum,链荚 豆Alysicarpus vaginalis,羽芒菊Tridax procumbens,红花材(椭圆叶木蓝)Indigofera cassoides,黄珠子草Phyllanthus virgatus,白 花叶Porana henryi,响铃豆Crotalaria albida,鬼针草Bidens pilosa,白羊草Bothriochloa ischaemum -
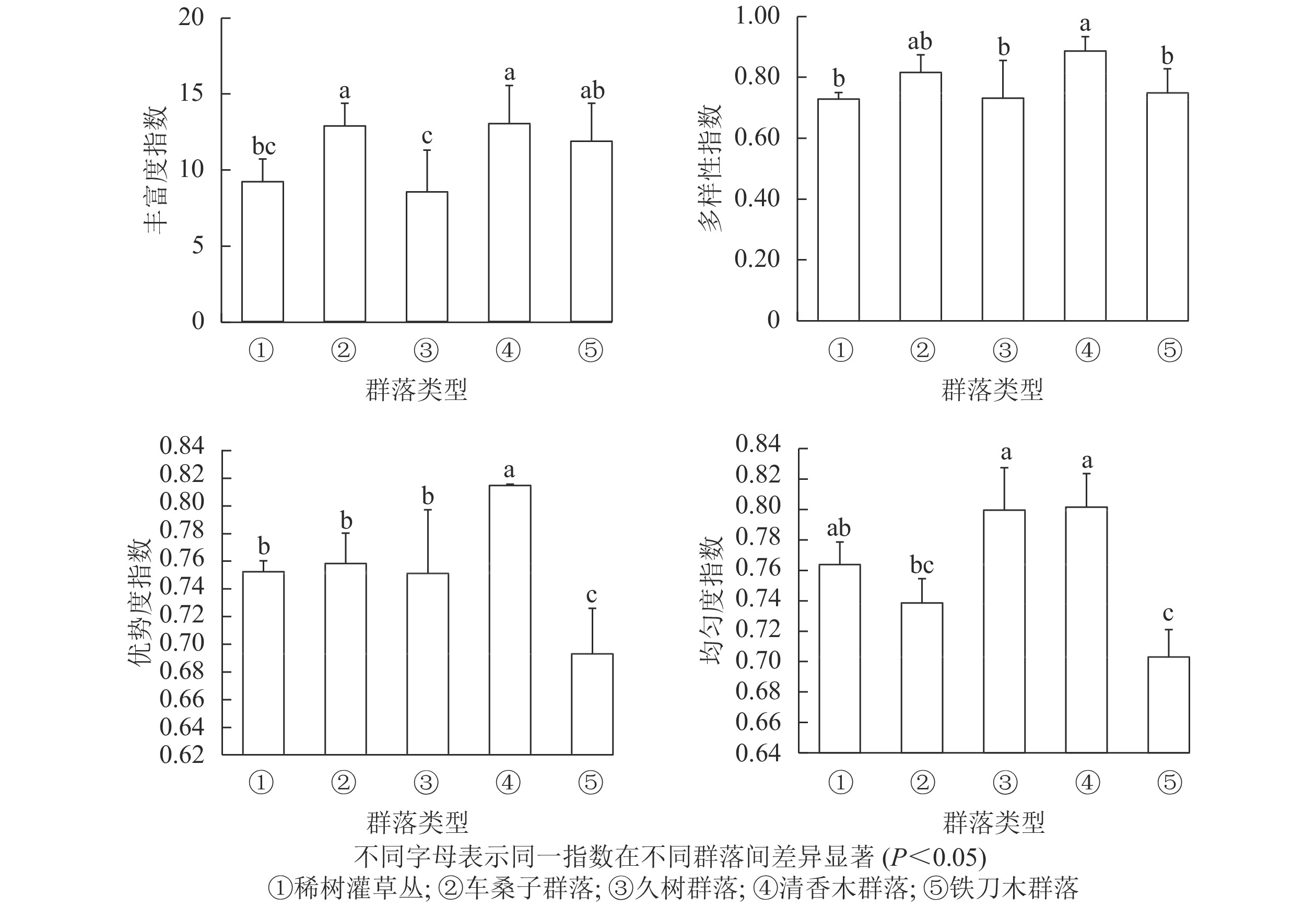
不同植物群落的丰富度指数从大到小依次为清香木群落、车桑子群落、铁刀木群落、稀树灌草丛、久树群落。其中,清香木群落和车桑子群落的丰富度指数显著高于稀树灌草丛(P<0.05),久树群落和铁刀木群落的丰富度指数与稀树灌草丛无显著差异(P>0.05)。说明人工恢复群落能够增加群落内物种总数,但不同恢复群落对林下物种丰富度的影响存在一定差异(图1)。在物种多样性方面,稀树灌草丛的多样性指数显著低于清香木群落(P<0.05),与其他人工恢复群落无显著差异(P>0.05)。
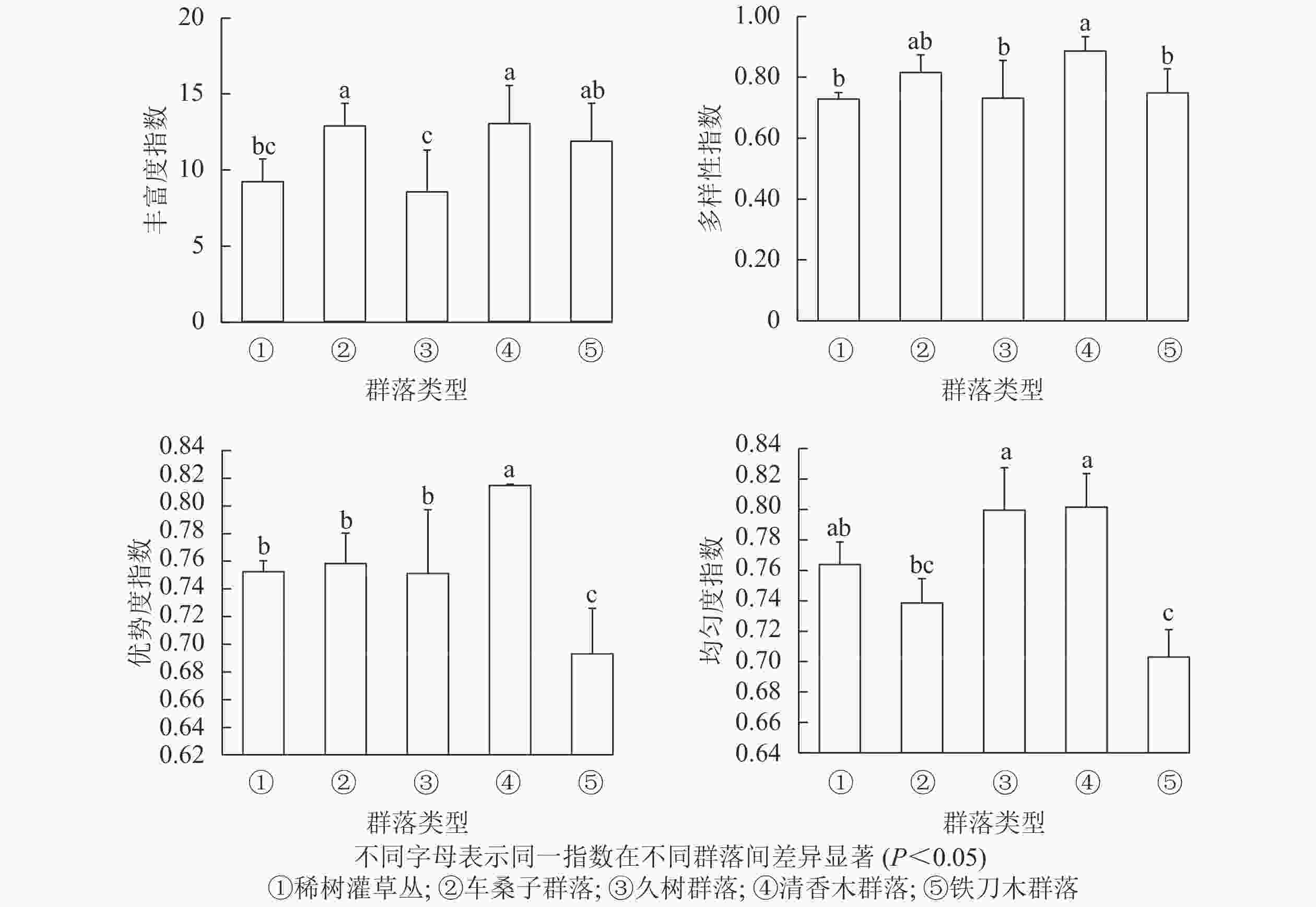
图 1 不同植被类型林下多样性指数比较
Figure 1. Comparison of understory species diversity index of different vegetation types
稀树灌草丛的优势度指数显著低于清香木群落(P<0.05)。铁刀木群落的优势度指数小于各人工恢复群落,且存在显著差异(P<0.05),说明相较于其他人工恢复群落,铁刀木群落不太稳定。稀树灌草丛与车桑子群落、久树群落和清香木群落的均匀度指数无显著差别(P>0.05),但显著高于铁刀木群落的均匀度指数(P<0.05)。
-
表3表明:不同类型群落间Jaccard相似系数为0.200 0~0.500 0,其中,车桑子群落与稀树灌草丛的相似系数最大,为0.424 2,清香木群落与稀树灌草丛的相似系数最小,为0.250 0,其他群落与稀树灌草丛的相似系数为0.250 0~0.300 0。人工恢复群落与稀树灌草丛之间的相似性程度较低,处于中等不相似水平,不同人工恢复群落在物种组成上存在较大差异。这表明通过人工植被恢复能够丰富红河干热河谷地区林地的物种组成,对物种多样性的建立具有重要作用。
表 3 不同群落间Jaccard相似系数
Table 3. Jaccard’ indexs among different communities
群落 车桑子
群落稀树灌
草丛久树
群落清香木
群落铁刀木
群落车桑子群落 1 稀树灌草丛 0.424 2 1 久树群落 0.305 6 0.263 1 1 清香木群落 0.384 6 0.250 0 0.250 0 1 铁刀木群落 0.457 1 0.300 0 0.238 1 0.404 8 1 -
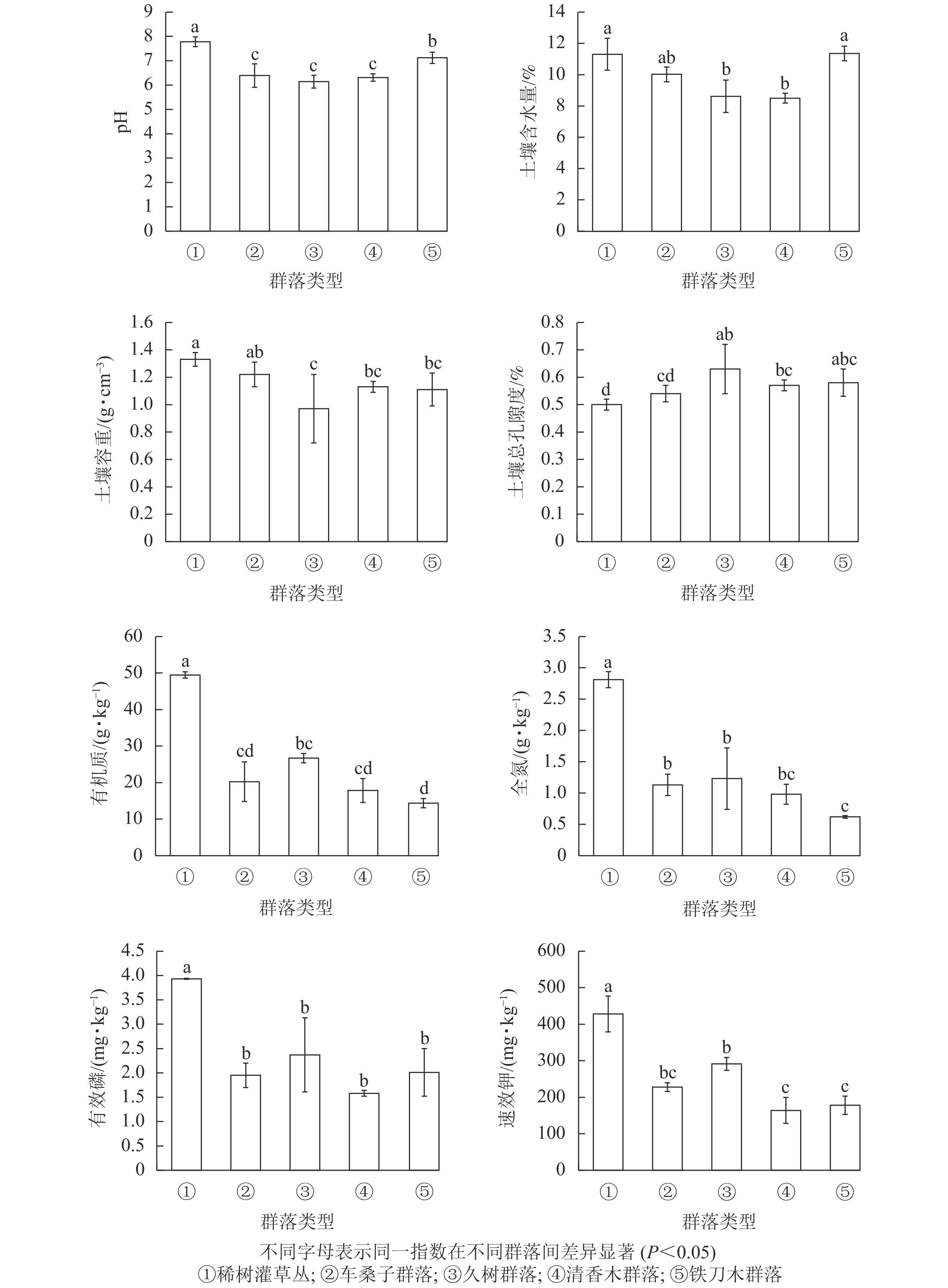
不同植物群落的土壤理化性质存在著显差异(图2),其中久树群落、车桑子群落和清香木群落的土壤pH为6.14~6.39,呈弱酸性,与稀树灌草丛的土壤pH存在显著差异(P<0.05)。久树群落及清香木群落的土壤含水量显著低于稀树灌草丛(P<0.05)。铁刀木群落、久树群落及清香木群落的土壤容重显著低于稀树灌草丛(P<0.05),其土壤总孔隙度显著高于稀树灌草丛(P<0.05)。另外,人工恢复群落的土壤有机质、全氮、有效磷以及速效钾质量分数显著低于稀树灌草丛(P<0.05)。
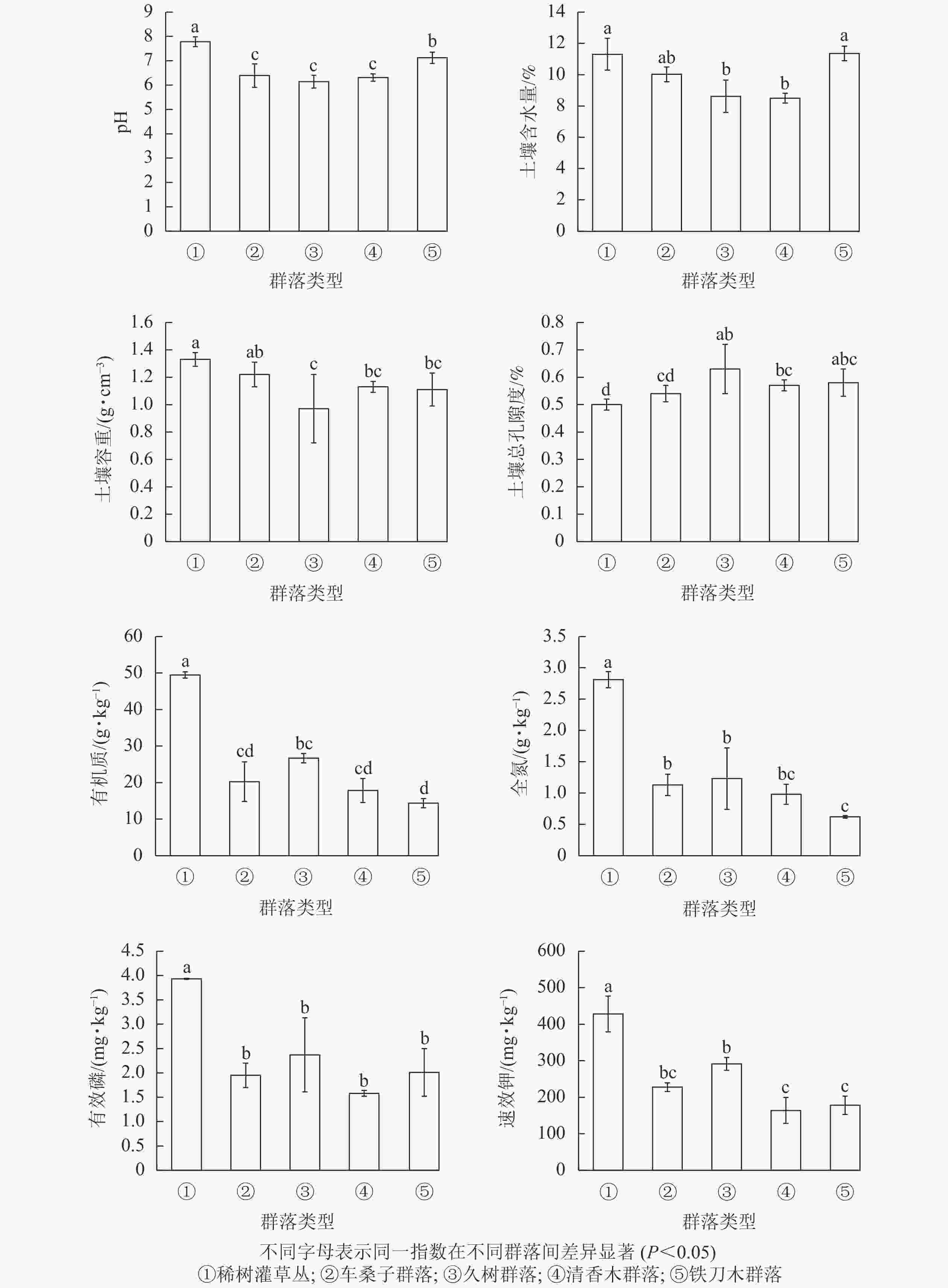
图 2 不同植被土壤理化性质变化情况
Figure 2. Changes in soil physical and chemical properties under different vegetation types
-
群落构建规则认为:环境筛选和生物的相互作用在不同尺度上决定了群落结构、组成和多样性[21-22],其中群落结构是群落中植物与植物间、植物与环境间相互关系的体现[23]。自然条件下,群落结构的更替是一个相对漫长且不确定的过程。人类活动会对植物群落结构产生多方面干扰。红河干热河谷区域植物种类稀少,群落结构简单,生态环境脆弱,在受到放牧和采樵等人为干扰后,限制了物种生存、群落发展及多样性维持,导致干热河谷生态系统进一步退化,结构和功能也濒临崩溃[3]。为了有效恢复红河干热河谷区的生态系统与物种多样性,除了限制放牧、采樵等负面人为干扰,还需要进行合理的正面干扰,其中,人工植被恢复可以作为物种多样性恢复与生态治理的有效方法。人工恢复群落可以绕过植物群落演替早期物种扩散和建立的障碍,增加新生植物群落中的物种数量,加快物种多样性的恢复[17, 24]。本研究中,大多数人工恢复群落的植物多样性指数和物种丰富度指数已经高于稀树灌草丛,表明红河干热河谷不同人工恢复群落在恢复15 a后,林下植物的多样性得到了显著提高。这一结果与以往研究基本一致[25]。在人工恢复群落中,植物群落的上层分别是清香木、车桑子、铁刀木和久树等乔灌木。这些阳性植被恢复先锋树种在恢复初期通过萌生扩展了林冠面积,从而适应或利用干热河谷的高光条件。作为上层植被,这些树种可以通过遮光并产生更多的凋落物等来减轻土壤水分挥发,避免强光直接照射,为下层植被营造较好的生存环境,让中性乃至耐阴物种占据此生态位。这可能是该区域进行人工植被恢复后林下植物多样性提高的主要原因。
物种多样性和稳定性作为植物群落的2个属性,其相互关系已经受到多位生态学研究者的关注[23, 26-27]。植物群落的物种多样性能够体现群落结构的稳定程度和生境差异等,因此可以作为判断群落稳定性的指标[28-29]。此外,均匀度也可以衡量物种多样性与群落稳定性之间的关系,物种均匀度指数越高表明群落的稳定性也越高[23-24]。本研究结果表明:不同人工恢复群落不仅改变了植物群落的物种组成,而且导致其结构特征和物种多样性发生改变。清香木群落的丰富度指数、多样性指数和优势度指数均显著高于稀树灌草丛(P<0.05),较高的物种多样性为该群落的稳定性提供了必要条件。研究结果证明:清香木群落与稀树灌草丛的均匀度指数无显著差异(P>0.05),说明选择清香木作为恢复树种不仅能增加群落内的物种个数,而且各物种在群落中仍具有合适的生态位,使植物群落维持在一个相对稳定、合理的状态,因此可以优先考虑将清香木作为红河干热河谷区植被恢复的先锋树种。
土壤养分与植被群落结构和功能关系密切,因此,不同人工恢复群落不仅改变了红河干热河谷区的物种组成,而且也在一定程度上影响土壤的理化性质[30]。有研究表明:植被能够增加土壤中有机质和全氮质量分数,有效地恢复土壤养分[31-32]。然而本研究发现:在红河干热河谷区不同人工恢复群落中的有机质、全氮、有效磷和速效钾质量分数均显著低于自然条件下恢复的稀树灌草丛(P<0.05),不同人工恢复群落土壤中有机质和全氮质量分数从高到低依次为稀树灌草丛、久树群落、车桑子群落、清香木群落、铁刀木群落,有效磷和速效钾质量分数从高到低依次为稀树灌草丛、久树群落、车桑子群落、铁刀木群落、清香木群落。本研究涉及的人工恢复群落种植密度均较大,这样人工植被可以在短期内形成乔木层的结构,也能在地面形成一定数量的凋落物层,具有较高的生产力和养分存储能力,但是,乔木层的生长需要获取更多的土壤营养物质,其产生的凋落物又无法很快将养分归还土壤,影响了土壤养分的再分配,从而使人工恢复群落的土壤养分相对较低。天然次生的稀树灌草丛中缺少高大的乔灌木且密度较低,对有机质、全氮、有效磷和速效钾的利用不多,所以大部分营养物质被存储在土壤中,同时人工恢复群落增加了土壤总孔隙度,使土壤功能下降,土壤微生物活性和营养循环受限[33-34],也将造成人工恢复群落的土壤养分下降。另一方面,在人工恢复群落前进行了整地松土等工作,不可避免地对土壤表层产生破坏,造成新的水土与养分流失[35],说明在过度退化的干热河谷地区,短期内自然恢复群落的水土保持功能要高于人工恢复群落。有研究表明:干热河谷区优势乡土草本植物扭黄茅、孔颖草Bothriochloa pertusa及拟金茅Eulaliopsis binat等能显著增加土壤的全碳、全氮含量[31],作为干热河谷的优势物种,有利于恢复土壤功能。因此,为了加快土壤养分的恢复,在干热河谷区进行人工恢复时,可采用乔草或灌草并行的恢复模式。
干热河谷区是中国生态环境极为脆弱的区域之一。严酷的自然条件和长期的人为干扰,已使该地区成为中国生态极度退化和造林极为困难的地区之一。本研究结果表明:在红河干热河谷区选择车桑子、久树、铁刀木和清香木等4个树种进行人工植被恢复后,能够在短期内提高林下植被的物种多样性。恢复后的植物群落结构也能够朝着较稳定的方向演替。但不同人工恢复群落产生的效果不同,其中,清香木群落在人工恢复15 a后,林下植被具有较高的物种多样性,且群落稳定性较高,可优先考虑作为植物多样性恢复的先锋树种。同时,本研究也证实,不同恢复群落对土壤理化性质的改善存在较大差异,而且,在红河干热河谷区,人工恢复群落对土壤理化性质的改善相对于植物多样性的提高是一个更为缓慢的过程,需要不断地研究和探讨更好的人工促进恢复模式。
Effects of different restoration communities on understory species diversity and soil physical and chemical properties in dry-hot valley
-
摘要:
目的 揭示红河干热河谷不同恢复群落对林下物种多样性和土壤理化性质的影响。 方法 以红河干热河谷人工恢复15 a的久树Schleichera oleosa群落、车桑子Dodonaea viscosa群落、铁刀木Cassia siamea群落、清香木Pistacia weinmannifolia群落等植物群落和天然次生稀树灌草丛为对象,采用典型样地和随机取样的方法,对林下草本层植物的物种组成、多样性特征以及土壤理化性质进行比较研究。 结果 ①不同恢复群落内,共发现维管植物60种,隶属于20科49属,其中优势科为豆科Leguminosae、菊科Asteraceae、禾本科Poaceae和大戟科Euphorbiaceae。②4种人工恢复群落中,清香木群落的丰富度指数、Shannon-Wiener多样性指数、优势度指数均显著高于天然次生稀树灌草丛(P<0.05)。③4种人工恢复群落在物种组成上均与天然次生稀树灌草丛处于中等不相似水平。④4种人工恢复群落的土壤有机质、全氮、有效磷、速效钾均显著低于天然次生稀树灌草丛(P<0.05)。 结论 红河干热河谷不同人工恢复群落中,林下物种多样性和土壤理化性质均存在显著差异。与天然次生稀树灌草丛相比,人工恢复群落可以在相对较短的时间内增加林下物种多样性,但不能快速恢复土壤养分。图2表3参35 Abstract:Objective This study aims to reveal the effects of different restoration communities on understory species diversity and soil physical and chemical properties in the dry-hot valley of the Red River. Method The plant communities of Schleichera oleosa, Dodonaea viscosa, Cassia siamea, Pistacia weinmannifolia and natural secondary sparse shrub and grass artificially restored for 15 years in the dry-hot valley of the Red River were taken as the objects, the species composition, diversity characteristics and soil physical and chemical properties of understory herbaceous plants were studied by using typical plots and random sampling methods. Result (1) A total of 60 species of vascular plants were found in different restoration communities, belonging to 20 families and 49 genera, of which the dominant families were Leguminosae, Compositae, Gramineae, and Euphorbiaceae. (2) Among the four artificial restoration communities, the Richness index, Shannon-Wiener diversity index and dominance index of P. weinmannifolia were significantly higher than those of natural secondary sparse shrub and grass (P<0.05). (3) The similarity degree of species composition between the four artificial restoration communities and the sparse shrub and grass was at a moderate dissimilarity level. (4) The soil organic matter, total nitrogen, available phosphorus and available potassium of the four artificially restored plant communities were significantly lower than those of sparse shrub and grass (P<0.05). Conclusion There are significant differences in understory species diversity and soil physical and chemical properties among different restoration communities in the dry-hot valley of the Red River. Compared with the natural secondary sparse shrub and grass, the artificial restoration community can increase the species diversity in a relatively short period of time, but cannot quickly restore soil nutrients. [Ch, 2 fig. 3 tab. 35 ref.] -
图 1 不同植被类型林下多样性指数比较
Figure 1 Comparison of understory species diversity index of different vegetation types
图 2 不同植被土壤理化性质变化情况
Figure 2 Changes in soil physical and chemical properties under different vegetation types
表 1 样地基本情况
Table 1. Basic information of sample plots
样地 海拔/
m平均胸
径/cm枝下高/
m平均冠幅/
m林分密度/
(株·hm−2)久树群落 530 5.06 1.58 3.32 2 867 车桑子群落 675 3.02 1.94 1.81 1 650 铁刀木群落 520 3.63 0.82 2.85 2 017 清香木群落 850 4.73 0.77 4.72 3 267 稀树灌草丛 702 1.82 1.84 2.03 1 200  下载: 导出CSV
下载: 导出CSV
表 2 各群落中重要值位于前10位的物种
Table 2. Species with top 10 important values in each community
序号 稀树灌草丛 车桑子群落 久树群落 清香木群落 铁刀木群落 植物 重要值 植物 重要值 植物 重要值 植物 重要值 植物 重要值 1 扭黄茅 40.81 扭黄茅 42.77 扭黄茅 37.33 扭黄茅 29.87 扭黄茅 51.74 2 水蔗草 18.34 独穗飘拂草 19.37 狸尾豆 15.90 耳草 18.11 蔓草虫豆 9.57 3 独穗飘拂草 8.77 车桑子 6.67 飞机草 12.15 藿香蓟 11.08 三点金 7.69 4 蔓草虫豆 4.20 三点金 5.81 迎春花 7.52 飞机草 6.92 车桑子 3.22 5 矛叶荩草 3.70 飞机草 4.01 华西小石积 4.07 蔓草虫豆 4.86 猪屎豆 2.65 6 野拔子 3.11 蔓草虫豆 3.11 单叶拿身草 4.06 链荚豆 3.31 羽芒菊 2.38 7 椭圆叶木蓝 2.76 黄珠子草 3.10 车桑子 3.42 三点金 3.21 黄珠子草 2.26 8 白花叶 2.76 响铃豆 2.22 久树 2.04 铁刀木 3.11 鬼针草 1.98 9 车桑子 2.73 狸尾豆 1.79 三点金 1.74 黄珠子草 2.44 链荚豆 1.87 10 狸尾豆 1.88 羽芒菊 1.47 椭圆叶木蓝 1.48 车桑子 2.19 白羊草 1.81 说明:独穗飘拂草Fimbristylis monostachya,狸尾豆Uraria lagopodioides,耳草Hedyotis auricularia,蔓草虫豆Cajanus scarabaeoides, 飞机草Eupatorium odoratum,藿香蓟Ageratum conyzoides,三点金Desmodium triflorum,迎春花Jasminum nudiflorum,矛叶荩 草Arthraxon lanceolatus,华西小石积Osteomeles schwerinae,猪屎豆Crotalaria pallida,单叶拿身草Desmodium zonatum,链荚 豆Alysicarpus vaginalis,羽芒菊Tridax procumbens,红花材(椭圆叶木蓝)Indigofera cassoides,黄珠子草Phyllanthus virgatus,白 花叶Porana henryi,响铃豆Crotalaria albida,鬼针草Bidens pilosa,白羊草Bothriochloa ischaemum
下载: 导出CSV
表 3 不同群落间Jaccard相似系数
Table 3. Jaccard’ indexs among different communities
群落 车桑子
群落稀树灌
草丛久树
群落清香木
群落铁刀木
群落车桑子群落 1 稀树灌草丛 0.424 2 1 久树群落 0.305 6 0.263 1 1 清香木群落 0.384 6 0.250 0 0.250 0 1 铁刀木群落 0.457 1 0.300 0 0.238 1 0.404 8 1
下载: 导出CSV
-
[1] 刘颖, 贺静雯, 李松阳, 等. 干热河谷优势灌木种类的根系结构及碳氮磷元素含量特征[J]. 应用与环境生物学报, 2020, 26(1): 31 − 39. LIU Ying, HE Jingwen, LI Songyang, et al. Root structure and element (C, N, P) content characteristics of dominant shrub species in a dry-hot valley [J]. Chin J Appl Environ Biol, 2020, 26(1): 31 − 39. [2] 杨济达, 张志明, 沈泽昊, 等. 云南干热河谷植被与环境研究进展[J]. 生物多样性, 2016, 24(4): 462 − 474. YANG Jida, ZHANG Zhiming, SHEN Zehao, et al. Review of research on the vegetation and environment of dry-hot valleys in Yunnan [J]. Biodiversity Sci, 2016, 24(4): 462 − 474. [3] 李昆, 刘方炎, 杨振寅, 等. 中国西南干热河谷植被恢复研究现状与发展趋势[J]. 世界林业研究, 2011, 24(4): 55 − 60. LI Kun, LIU Fangyan, YANG Zhenyin, et al. Study status and trends of vegetation restoration of dry-hot valley in southwest China [J]. World For Res, 2011, 24(4): 55 − 60. [4] YUAN Yong, XIONG Dong, WU Han, et al. Spatial variation of soil physical properties and its relationship with plant biomass in degraded slopes in dry-hot valley region of Southwest China [J]. J Soil Sediment, 2020, 20(5): 2354 − 2366. [5] 金振洲. 云南元江干热河谷半萨王纳植被的植物群落学研究[J]. 广西植物, 1999, 19(4): 289 − 302. JIN Zhenzhou. A phytosociological study on the semi-savanna vegetation in the dry-hot valleys of Yuanjiang River, Yunnan [J]. Guihaia, 1999, 19(4): 289 − 302. [6] FLYNN D F B, MIROTCHINK N, JAIN M, et al. Functional and phylogenetic diversity as predictors of biodiversity-ecosystem-function relationships [J]. Ecology, 2011, 92(8): 1573 − 1581. [7] WANG Jianming, CHEN Chen, LI Jingwen, et al. Different ecological processes determined the alpha and beta components of taxonomic, functional, and phylogenetic diversity for plant communities in dryland regions of Northwest China[J/OL]. Peer J, 2019, 6: e6220[2021-04-22]. doi: 10.7717/peerj.6220. [8] THOMPSON P L, RAYFIELD B, GONZALEZ A. Loss of habitat and connectivity erodes species diversity, ecosystem functioning, and stability in metacommunity networks [J]. Ecography, 2017, 40(1): 98 − 108. [9] ISBELL F I, CRAVEN D, CONNOLLY J, et al. Biodiversity increases the resistance of ecosystem productivity to climate extremes [J]. Nature, 2015, 526(7574): 574 − 577. [10] 金艳强, 李敬, 张一平, 等. 元江干热河谷稀树灌草丛植被碳储量及净初级生产力[J]. 生态学报, 2017, 37(17): 5584 − 5590. JIN Yanqiang, LI Jing, ZHANG Yiping, et al. Carbon storage and net primary productivity of a savanna ecosystem in a dry-hot valley in Yuanjiang, Yunnan [J]. Acta Ecol Sin, 2017, 37(17): 5584 − 5590. [11] 沈蕊, 张建利, 何彪, 等. 元江流域干热河谷草地植物群落结构特征与相似性分析[J]. 生态环境学报, 2010, 19(12): 2821 − 2825. SHEN Rui, ZHANG Jianli, HE Biao, et al. The structure characteristic and analysis on similarity of grassland community in dry-hot valley of Yuanjiang River [J]. Ecol Environ Sci, 2010, 19(12): 2821 − 2825. [12] 金振洲, 欧晓昆. 干热河谷植被[M]. 昆明: 云南大学出版社, 2000. JIN Zhenzhou, QU Xiaokun. Dry and Hot Valley Vegetation [M]. Kunming: Yunnan University Press, 2000. [13] 夏顺颖. 绿汁江流域河谷植被的植物区系与群落类型的研究[D]. 昆明: 云南大学, 2018. XIA Shunying. The Study of Plant Community and Flora in Luzhi River Valley in Yunnan, China[D]. Kunming: Yunnan University, 2018. [14] 王小庆, 刘方炎, 李昆, 等. 元谋干热河谷滇榄仁群落林下物种多样性与幼苗更新特征[J]. 浙江农林大学学报, 2011, 28(2): 241 − 247. WANG Xiaoqing, LIU Fangyan, LI Kun, et al. Species diversity and seedling regeneration of three Terminalia franchetii communities in the hot-dry Yuanmou valley [J]. J Zhejiang A&F Univ, 2011, 28(2): 241 − 247. [15] 校亮, 熊东红, 张宝军, 等. 干热河谷冲沟沟岸葛藤不同覆被状况及土壤性质差异分析[J]. 生态学报, 2018, 38(14): 5047 − 5055. XIAO Liang, XIONG Donghong, ZHANG Baojun, et al. Analysis of soil properties in gully banks under different coverage of Kudzu in a dry-hot valley region [J]. Acta Ecol Sin, 2018, 38(14): 5047 − 5055. [16] ZHANG Hao, JIM C Y. Contributions of landscape trees in public housing estates to urban biodiversity in Hong Kong [J]. Urban For Urban Greening, 2014, 13(2): 272 − 284. [17] 何斌, 李青, 刘勇. 草海国家级自然保护区华山松群落特征及物种多样性研究[J]. 热带亚热带植物学报, 2020, 28(1): 44 − 52. HE Bin, LI Qing, LIU Yong. Community characteristics and species diversity of Pinus armandii in Caohai National Nature Reserve [J]. J Trop Subtrop Bot, 2020, 28(1): 44 − 52. [18] LI Wen, DOU Zhiguo, CUI Lijuan, et al. Soil fauna diversity at different stages of reed restoration in a lakeshore wetland at Lake Taihu, China[J/OL]. Ecosyst Health Sustainability, 2020, 6(1)[2021-04-22]. doi: 10.1080/20964129.2020.1722034. [19] YANG Haijun, WU Mingyu, LIU Wenxing, et al. Community structure and composition in response to climate change in a temperate steppe [J]. Global Change Biol, 2011, 17(1): 452 − 465. [20] 谭速进, 魏翰均, 刘丹碧. 成都地区居室庭院蚂蚁区系及群落相似性分析[J]. 环境昆虫学报, 2010, 32(1): 11 − 19. TAN Sujin, WEI Hanjun, LIU Danbi. Study on fauna and similarity coefficients among communities of house and courtyard ants in Chengdu area [J]. J Environ Entomol, 2010, 32(1): 11 − 19. [21] KEDDY P A. Assembly and response rules: two goals for predictive community ecology [J]. J Veg Sci, 1992, 3(2): 157 − 164. [22] 王铮屹, 戴其林, 柏宬, 等. 天目山皆伐毛竹林自然更新群落类型与多样性分析[J]. 浙江农林大学学报, 2020, 37(4): 710 − 719. WANG Zhengyi, DAI Qilin, BAI Cheng, et al. Types and diversity of natural regeneration community after clear cutting of Phyllostachys edulis forests in Mount Tianmu, China [J]. J Zhejiang A&F Univ, 2020, 37(4): 710 − 719. [23] 谢立红, 曹宏杰, 黄庆阳, 等. 五大连池火山森林群落多样性与稳定性[J]. 浙江农林大学学报, 2021, 38(2): 235 − 245. XIE Lihong, CAO Hongjie, HUANG Qingyang, et al. On the diversity and stability of forest communties in Wudalianchi volcanoes [J]. J Zhejiang A&F Univ, 2021, 38(2): 235 − 245. [24] 刘斌, 张参参, 汪金松, 等. 江西九连山不同恢复模式林分的物种多样性特征[J]. 林业科学研究, 2020, 33(4): 42 − 52. LIU Bin, ZHANG Cancan, WANG Jinsong, et al. Characteristic of species diversity in stands of different restoration models in Jiulian mountain, Jiangxi Province [J]. For Res, 2020, 33(4): 42 − 52. [25] 杨振寅, 苏建荣, 李从富, 等. 元谋干热河谷主要植物群落物种多样性研究[J]. 林业科学研究, 2008, 21(2): 200 − 205. YANG Zhenyin, SU Jianrong, LI Congfu, et al. Species diversity of main communities in hot and dry valley, Yuanmao County [J]. For Res, 2008, 21(2): 200 − 205. [26] LOREAU M, de MAZANCOURT C. Biodiversity and ecosystem stability: a synthesis of underlying mechanisms [J]. Ecol Lett, 2013, 16(suppl 1): 106 − 115. [27] 谢晋阳, 陈灵芝. 中国暖温带若干灌丛群落多样性问题的研究[J]. 植物生态学报, 1997, 21(3): 197 − 207. XIE Jinyang, CHEN Lingzhi. The studies of some aspects of biodiversity on scrubs in the warm temperate zone in China [J]. Acta Phytoecol Sin, 1997, 21(3): 197 − 207. [28] 吴昊, 杜奎, 李万通, 等. 空心莲子草入侵对豫南草本植物群落多样性及稳定性的影响[J]. 草业科学, 2019, 36(2): 382 − 393. WU Hao, DU Kui, LI Wangtong, et al. Influence of Alternanthera philoxeroides invasion on species diversity and stability in the herbaceous community in southern Henan Province [J]. Pratacultural Sci, 2019, 36(2): 382 − 393. [29] GHALEY B B, PORTER J R. Ecosysten function and service quantification and valuation in a conventional winter wheat production system with DAISY model in Denmark [J]. Ecosystem Serv, 2014, 10: 79 − 83. [30] THOMPSON J, BROKAW N, ZIMMERMAN J, et al. Land use history, environment, and tree composition in a tropical forest [J]. Ecol Appl, 2002, 12(5): 1344 − 1363. [31] 和润莲, 闫帮国, 孙毅, 等. 植物种和氮添加对元谋干热河谷草地土壤有机碳分解的影响[J]. 应用与环境生物学报, 2021, 27(4): 1 − 13. HE Runlian, YAN Bangguo, SUN Yi, et al. Effects of plant species and nitrogen addition on soil organic carbon decomposition in the Yuanmou dry-hot valley grassland, China [J]. Chin J Appl Environ Biol, 2021, 27(4): 1 − 13. [32] 杨智姣, 温晨, 杨磊, 等. 半干旱黄土小流域不同恢复方式对生态系统多功能性的影响[J]. 生态学报, 2020, 40(23): 8606 − 8617. YANG Zhijiao, WEN Chen, YANG Lei, et al. Effects of different restoration methods on ecosystem multifunctionality in the semi-arid loess small watershed [J]. Acta Ecol Sin, 2020, 40(23): 8606 − 8617. [33] RABOT E, WIESMEIER M, SCHLUTER S, et al. Soil structure as an indicator of soil functions: a review [J]. Geoderma, 2018, 314: 122 − 137. [34] LI Xingfu, ZHANG Ying, DING Chengxiang, et al. Water addition promotes vegetation recovery of degraded alpine meadows by regulating soil enzyme activity and nutrients in the Qinghai-Tibetan Plateau[J/OL]. Ecol Eng, 2020, 158[2021-04-22]. doi: 10.1016/j.ecoleng.2020.106047. [35] 唐国勇, 高成杰, 李昆. 植被恢复对干热河谷退化土壤改良的影响[J]. 生态学报, 2015, 35(15): 5157 − 5167. TANG Guoyong, GAO Chenjie, LI Kun. Effects of vegetation restoration on the amelioration of degraded soil in a dry-hot valley [J]. Acta Ecol Sin, 2015, 35(15): 5157 − 5167. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210348
 点击查看大图
点击查看大图
计量
- 文章访问数: 2312
- HTML全文浏览量: 509
- PDF下载量: 295
- 被引次数: 0



