







-
山核桃Carya cathayensis是果木兼用的优良经济树种,主要分布在浙皖交界的天目山区[1]。目前,山核桃主要采用实生苗繁殖与嫁接繁殖。实生苗繁殖难以保持品种的优良特性[2]。山核桃体细胞胚具有完整的两极结构,能一次再生形成完整植株[3]。因为生产对良种苗木的需要,山核桃的离体繁殖技术已得到了较深入的研究[4]。表观遗传是指在DNA序列不改变的情况下发生的可遗传的基因表达改变,DNA甲基化是表观遗传修饰的一种方式[5],是指在DNA甲基转移酶催化下以S-腺苷甲硫氨酸为甲基供体,将甲基转移到胞嘧啶上形成5-甲基胞嘧啶,不改变DNA一级结构对基因序列进行修饰而影响基因的表达,影响其功能,这种甲基修饰主要发生在CpG双核甘酸序列的胞嘧啶上[6]。已有众多证据表明:DNA甲基化参与了植物不同发育阶段的生长调控[7]。研究发现,以山核桃幼胚为外植体进行体胚诱导,不同发育时期幼胚胚性感受态差异较大,其中自然授粉后8~10周可诱导产生愈合组织,11~12周可从幼胚子叶表面直接诱导产生体细胞胚[8]。山核桃不同程度胚性感受态是否由于不同发育时期幼胚基因组表观遗传的变化引起,尚未见相关研究报道。本研究应用简单序列重复区间扩增(inter-simple sequence repeat,ISSR)DNA分子标记技术对山核桃幼胚生长过程中DNA甲基化变化动态进行研究,揭示山核桃幼胚发育过程中甲基化变化规律,以期对建立高效的山核桃体细胞胚发生体系提供理论依据。
HTML
-
7月上旬于浙江省临安市板桥乡罗塘村采集自然授粉后9,10,11,12,13周的山核桃幼果(分别以样本第9周,第10周,第11周,第12周,第13周代替)。将幼果带回实验室中剪去花柱及柱头,洗涤剂洗净表面后,用体积分数为75%乙醇涮洗30 s,自来水冲洗干净,于超净工作台上去除果皮和种皮后取出完整的幼胚,提取基因组DNA。
-
用改良十六烷基三甲基溴化铵(CTAB)法提取实验材料的基因组DNA[9]。
-
用提取的不同时期培养的山核桃幼胚的一组DNA为模版,所用引物参照加拿大哥伦比亚大学网站公布的第9套ISSR引物序列,对100个引物进行筛选,选取扩增条带多而清晰、分离度高的引物。本实验共筛选出共17个引物,分别是:UBC808,809,815,816,817,822,825,827,830,834,840,855,858,862,867,888,889。17个引物序列如表 1。
引物 序列 808 AGAGAGAGAGAGAGAGC 809 AGA GAG AGA GAG AGA GG 815 CTC TCT CTC TCT CTC TG 816 CACACACACACACACAT 817 CACACACACACACACAA 822 TCT CTC TCT CTC TCT CA 825 ACA CAC ACA CAC ACA CT 827 ACA CAC ACA CAC ACA CG 830 TGTGTGTGTGTGTGTGG 834 AGAGAGAGAGAGAGAGYT 840 GAG AGA GAG AGA GAG AYT 855 ACACACACACACACACYT 858 TGTGTGTGTGTGTGTGRT 862 AGC AGC AGC AGC AGC AGC 867 GGC GGC GGC GGC GGC GGC 888 BDB CAC ACA CAC ACA CA 889 DBD ACA CAC ACA CAC A 说明% N=(A,G,C,T),R=(A,G),Y=(C,T),B=(C,G,T) (I.e.not A),Dr (A,G,T)(I. e. not C),Hr (A,C,T)(I. e. not G),V=(A,C,G) (I. e. not T)。 Table 1. Primers for ISSR
-
取5个不同时期培养得幼胚DNA样品,分别用Hap Ⅱ和MSPⅠ等2种酶切,反应体系为30 μL,所用DNA质量浓度为50 mg·L-1,为了使酶切完全,加入3倍完全酶切的用量,酶切过夜后在65 ℃水浴锅10 min使酶失活。
-
分别以各个材料未酶切,Hap Ⅱ和MSPⅠ等2种酶切的基因组DNA为模版,以筛选的17个UBC引物进行聚合酶链式反应(PCR)。PCR反应条件:94 ℃预变性5 min,94 ℃变性30 s,适温退火30 s,72 ℃延伸1 min,35个循环后72 ℃延伸10 min。扩增产物用质量分数为20.0 g·kg-1琼脂糖凝胶电泳检测[10]。
-
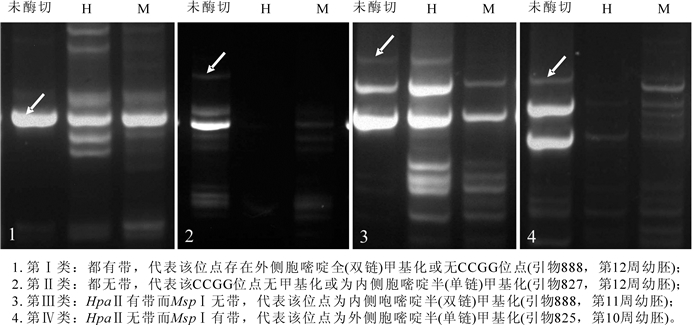
MSPⅠ和HapⅡ对相同酶切位点甲基化的敏感程度不同以及ISSR是显性标记是分析甲基化变异的理论基础[11]。同裂酶MSPⅠ和HpaⅡ的酶切位点均为“CCGG/GGCC”,由于两者对该位点胞嘧啶甲基化的敏感程度不同,经PCR反应扩增多态性片段就反映出该位点的甲基化状态及程度。MSPⅠ能切割无甲基化和内甲基化位点而不能切割外甲基化位点,HpaⅡ能切割无甲基化和单链甲基化(双链中只有链甲基化)而不能切割双链甲基化[12]。因而DNA分别用MSPⅠ和HpaⅡ酶切后选扩,根据带型的有无可能出现4种甲基化类型,Ⅰ型:都有带,代表该位点存在外侧胞嘧啶全(双链)甲基化或无CCGG位点;Ⅱ型:都无带,代表该CCGG位点无甲基化或为内侧胞嘧啶半(单链)甲基化;Ⅲ型:HpaⅡ有带而MSPⅠ无带,代表该位点为内侧胞嘧啶半(双链)甲基化;Ⅳ型:HpaⅡ无带而MSPⅠ有带,代表该位点为外侧胞嘧啶半(单链)甲基化。由表 2可知:Ⅲ型为全甲基化,Ⅳ型为半甲基化,但Ⅰ型和Ⅱ型的全/半甲基化状态仅根据条带位点还不确定,需进一步根据不同时期同一引物的同一位点条带差异来判断胞嘧啶全/半甲基化变异(表 3)。
类型 甲基化程度 内切酶活性 限制性条带 HpaⅡ Msp I 未酶切 H酶切 M酶切 I 无CCGG位点或5mCCGG GGCC5m 无活性 无活性 + + + Ⅱ CCGG或C5mCGG GGCC GGCC 有活性 有活性 + - - Ⅲ C5mCGG GGC5m: 无活性 有活性 + + - Ⅳ 5mCCGG CCGG 有活性 无活性 + - + Table 2. DNA methylation band types of uncut, HpaⅡand MSPⅠ of ISSR
条带类型变化 甲基化状态变化 Ⅰ-► Ⅱ 甲基化减少, Ⅰ为全甲基化变为Ⅱ为半甲基化 Ⅲ 甲基化减少, Ⅰ为全甲基化 Ⅳ 甲基化减少, Ⅰ为全甲基化 Ⅱ-► Ⅰ 甲基化增加, Ⅱ为半甲基化变成Ⅰ全甲基化 Ⅲ 甲基化增加, Ⅱ为半甲基化 Ⅳ 甲基化增加, Ⅱ为半甲基化 Ⅲ-► Ⅰ 甲基化增加, Ⅰ为全甲基化 Ⅱ 甲基化减少, Ⅱ为半甲基化 Ⅳ 甲基化变化情况不定 Ⅳ-► Ⅰ 甲基化增加, Ⅰ为全甲基化 Ⅱ 甲基化减少, Ⅱ为半甲基化 Ⅲ 甲基化变化情况不定 Table 3. DNA methylation of type Ⅰ and typeⅡ
1.1. 实验材料
1.2. 实验方法
1.2.1. 山核桃幼胚DNA的提取
1.2.2. 哥伦比亚大学(UBC)公布的引物的筛选
1.2.3. DNA酶切
1.2.4. ISSR-PCR建立
1.2.5. 山核桃幼胚基因组DNA甲基化水平分析
-
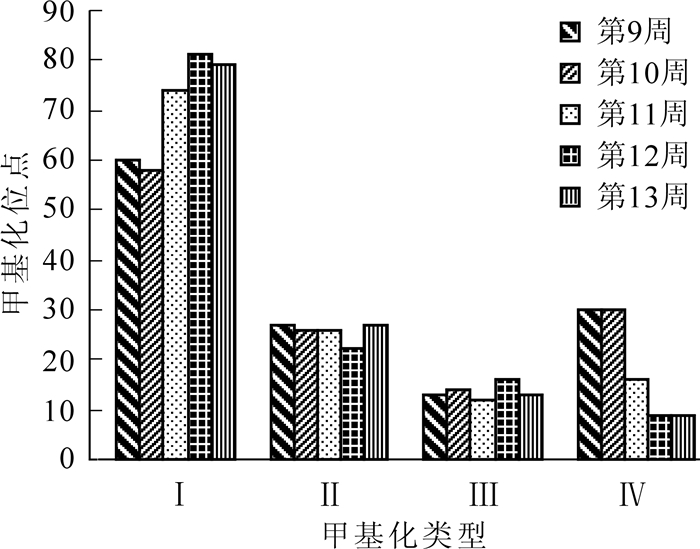
17个引物在各个发育时期山核桃幼胚样品中共扩增检测到的128个甲基化位点。5个时期幼胚甲基化比例依次为23.44%,25.78%,28.90%,40.62%和42.19%。甲基化比例随着胚的发育逐渐升高。研究结果同时表明,5个时期幼胚甲基化类型较丰富,4种甲基化类型均存在(图 1-1~4)。由表 4可知:在17个引物中,引物809扩增位点数最多,为11个,其中,第9周的Ⅰ~Ⅳ类型的扩增位点数分别为7,4,0,0;第10周的Ⅰ~Ⅳ类型的位点数分别为7,4,0,0;第11周的Ⅰ~Ⅳ型的位点数分别为7,4,0,0;第12周的Ⅰ~Ⅳ型的位点数分别为6,4,1,0;第13周的Ⅰ~Ⅳ型的位点数分别为6,4,1,0。图 2表明:随着幼胚的发育,4种DNA甲基化类型呈现规律性变化。第1种类型位点数最多,从第9周至13周DNA甲基化类型Ⅰ呈逐渐上升趋势,而Ⅳ型呈逐渐下降趋势;但在Ⅱ型、Ⅲ型中,各发育阶段变化相对平稳(图 2)。
引物 扩增
位点第9周 第10周 第11周 第12周 第13周 Ⅰ Ⅱ Ⅲ Ⅳ Ⅰ Ⅱ Ⅲ Ⅳ Ⅰ Ⅱ Ⅲ Ⅳ Ⅰ Ⅱ Ⅲ Ⅳ Ⅰ Ⅱ Ⅲ Ⅳ 808 7 6 0 1 0 5 1 1 0 6 1 0 0 6 0 0 1 6 1 0 0 809 11 7 4 0 0 7 4 0 0 7 4 0 0 6 4 1 0 6 4 1 0 815 7 1 4 0 2 2 4 0 1 2 4 0 1 6 1 0 0 5 2 0 0 816 5 3 0 0 2 3 0 0 2 3 0 0 2 3 0 0 2 3 0 0 2 817 8 3 0 0 5 3 0 0 5 3 0 0 5 5 0 0 3 5 0 0 3 822 9 5 3 1 0 5 3 1 0 5 4 0 0 5 4 0 0 5 4 0 0 825 8 3 0 0 5 3 0 0 6 6 0 2 0 6 0 2 0 6 0 2 0 827 9 1 2 0 6 1 2 0 6 6 1 1 1 5 1 0 3 5 3 0 1 830 6 4 2 0 0 4 2 0 0 4 2 2 0 5 1 0 0 5 1 0 0 834 7 3 2 0 2 3 2 0 2 3 2 0 2 3 2 0 2 4 2 0 1 840 9 5 2 2 0 5 2 2 0 5 2 2 0 6 3 0 0 6 3 0 0 855 6 1 1 1 3 2 1 0 3 1 1 1 3 2 1 0 3 3 0 0 3 858 9 4 3 2 0 4 3 2 0 4 3 2 0 5 3 1 0 6 3 0 0 862 6 2 2 1 1 5 0 0 1 5 0 1 0 2 2 1 1 4 1 1 0 867 6 5 1 0 0 5 1 0 0 5 1 0 0 5 0 1 0 5 1 0 0 888 7 3 1 0 3 4 0 0 3 6 1 0 0 6 1 0 0 7 0 0 0 889 8 3 0 0 5 3 0 0 5 7 0 1 0 7 0 1 0 7 0 1 0 总计 128 59 27 14 30 60 25 13 30 74 26 12 16 79 23 17 9 81 25 13 9 甲基化
比例/%23.44 25.78 28.90 40.62 42.19 Table 4. DNA methylation types on different development stages of immature embryos of ISSR

Figure 1. DNA methylation band types of different development stages of immature embryos of Carya cathayensis

Figure 2. Effect of development stage on DNA methylation types of immature embryos of Carya cathayensis
-
进一步分析表明:5个时期均存在全甲基化位点和半甲基化位点。5个时期全甲基化位点数分别为10,11,21,31,32,所占比例分别为7.81%,8.59%,16.40%,24.22%和25.00%;5个时期半甲基化位点数分别为20,22,16,21,22,所占比例分别为15.62%,17.19%,12.50%,16.41%,17.19%。由此可知:山核桃幼胚5个发育时期在基因组DNA甲基化水平上存在着较大差异,不同时期幼胚之间基因组DNA全/半甲基化位点的绝对数量和相对数量都有不同程度的改变。尽管总甲基化比例及全甲基化位点比例随着胚龄的增加而逐渐升高;半甲基化位点所占比例呈小幅波动并基本持平状态(表 5)。
引物 扩增位点 第9周 第10周 第11周 第12周 第13周 全甲基化
位点半甲基化
位点全甲基化
位点半甲基化
位点全甲基化
位点半甲基化
位点全甲基化
位点半甲基化
位点全甲基化
位点半甲基化
位点808 7 1 0 1 1 1 1 1 1 1 1 809 11 0 0 0 0 0 0 1 0 1 0 815 7 0 2 1 1 1 1 3 1 4 1 816 5 0 2 0 2 0 2 0 2 0 2 817 8 0 0 0 0 0 0 2 0 2 0 822 9 2 0 2 0 2 1 2 1 2 1 825 8 0 3 0 3 3 0 4 0 4 0 827 9 2 0 2 0 2 0 2 1 2 1 830 6 0 0 0 0 0 0 2 1 2 1 834 7 0 3 0 3 1 3 1 3 1 3 840 9 2 0 2 0 2 0 2 1 2 1 855 6 1 3 1 3 1 3 1 3 1 3 858 9 2 0 2 0 2 1 2 1 2 2 862 6 0 1 0 1 1 0 1 3 2 2 867 6 0 0 0 0 0 0 1 0 0 1 888 7 0 3 0 3 0 3 1 3 1 3 889 8 0 5 0 5 5 0 5 0 5 0 总计 128 10 20 11 22 21 16 31 21 32 22 甲基化比例/% 7.81 15.62 8.59 17.19 16.40 12.50 24.22 16.41 25.00 17.19 23.44 25.78 28.90 40.62 42.19 Table 5. Total/half DNA methylation on different development stages of ISSR
2.1. 山核桃幼胚不同发育时期对DNA甲基化类型的影响
2.2. 山核桃幼胚不同发育时期对胞嘧啶全/半甲基化变异的影响
-
DNA甲基化是植物正常生长发育所必需的,但是甲基化水平不足,也会导致不正常生长[13]。在高等植物基因组中,DNA甲基化在发育过程中处于一个动态的变化之中,同一物种在不同的发育时期,甲基化水平会发生变化,而且不同植物的变化趋势是不一样的[14]。越来越多的试验结果表明,DNA甲基化对植物正常发育起重要作用。对某些植物的研究发现:如果在基因组的重要位点上诱导产生或自然发生DNA甲基化的变异,就会引起广泛的发育异常现象[15]。油菜Brassica campestris种子在萌发时伴随着大量的去甲基化事件,导致甲基化水平的降低[16]。在同一植株的同一发育时期在不同组织中,DNA甲基化也有所差异,研究发现水稻Oryza sativa在幼苗时期的甲基化水平高于旗叶[17]。在不同的植物之间,同源的DNA序列的甲基化水平差异也非常明显,对11个不同被子植物的25S rDNA的甲基化研究发现,这些物种的甲基化水平差异较大,其中拟南芥Arabidopsis thaliana低至4%,而豌豆Pisum sativum和洋葱Allium cepa高达80%和90%[18]。前人的研究资料表明[19],植物在发育过程中,胞嘧啶的甲基化对基因具有重要的表达调控作用,在基因的内部或邻近区域发生甲基化可以抑制这些基因的表达,而去甲基化后又可以激活基因的表达。
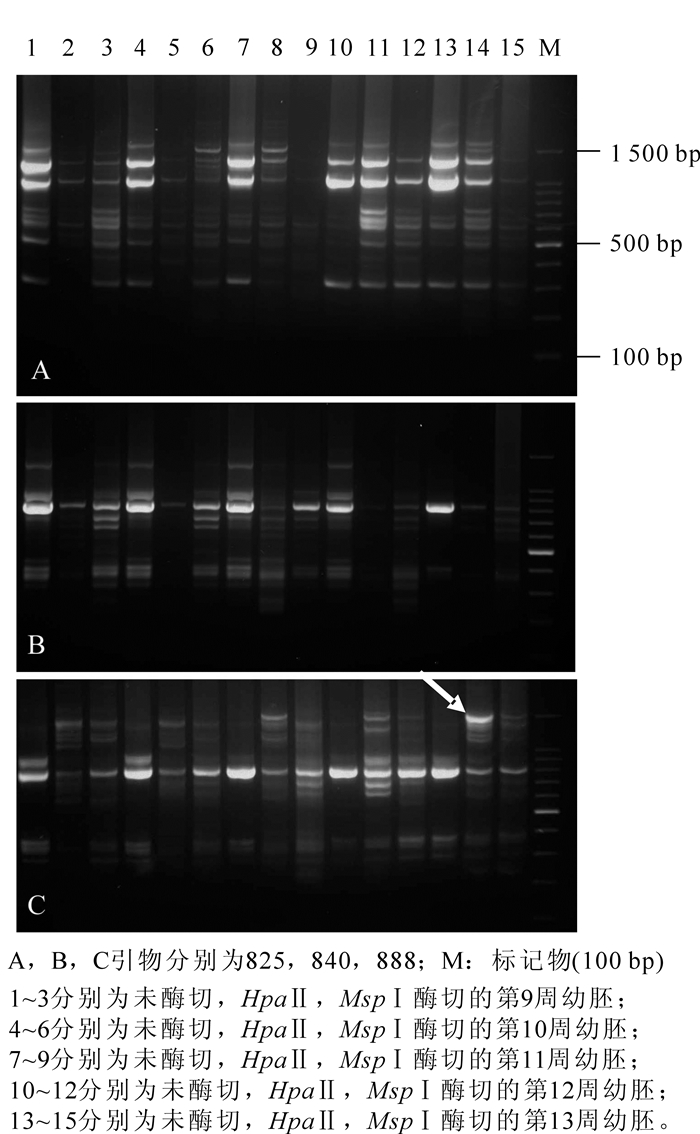
本研究用适当的PCR扩增反应从100个ISSR引物中筛选了17个引物,选择引物的标准是它们能产生和再次产生清晰的、多态性丰富的扩增产物(图 3)。在5个时期山核桃幼胚中,17条ISSR标记共产生了128个扩增条带。研究结果表明:5个发育时期山核桃发生了不同程度的甲基化模式和水平的变化,如DNA甲基化比例随着幼胚的发育逐渐升高,暗示部分基因正在关闭。同时,对于这些基因而言,去甲基化可能是基因进一步表达的必须步骤。这似乎表明:植物通过DNA甲基化和去甲基化的方式实现基因的有序表达[20]。根据全/半胞嘧啶甲基化均被观察到的结果,我们推测甲基化变异可能与幼胚成熟的一些重要属性有关。本研究在用MSPⅠ和HapⅡ酶切ISSR图谱和未酶切图谱比较时,还观察到了一种特殊的现象,即酶切后出现了未酶切没有的新条带(图 3,箭头所示)。理论上酶切ISSR图谱的出现的条带必然在未酶切ISSR图谱中出现,这种情况可能由于未酶切基因组存在一定的构象造成PCR引物结合困难,酶切后引物可以结合上去[21]。因此,尽管本研究已获得山核桃幼胚发育DNA甲基化状态的初步结果,但由于本研究尚未应用特异条带回收并杂交验证,关于山核桃幼胚不同发育阶段基因表达与基因组DNA甲基化以及胚性感受态之间关系还需要进一步探索。

Figure 3. Amplification results of immature embryos in five development stages by different ISSR primers
 DownLoad:
DownLoad: