




-
城市绿地对改善城市生态环境恶化等问题具有举足轻重的作用[1]。目前,对城市绿地生态效益的研究因受生态系统服务功能多样、区域环境及人类社会活动需求差异等因素影响,呈现出评估方法众多但评价内容、指标及结果差异性大等问题[2-3]。随着大数据等现代技术的应用,城市绿地生态效益价值的评估已经由原来的传统方法逐渐发展到计算机模型测算法[4-5]。由美国林务局2006年开发的i-Tree模型因树种匹配灵活、分析结果精确、评估对象尺度多元等优点,自推出至今在国际上尤其是北美国家已得到了大量的应用[6]。近年来中国合肥[7]、杭州[8]、南京[9]等地已有大量基于该模型的生态效益研究成果,但呈现出以研究城市森林等大区域尺度及城市社区行道树等小微尺度为主的特征,仅有魏云龙[10]、施炜婷等[11]、施健健等[5]对城市公园、广场这类中小尺度的绿地进行评估,且仅以研究单种生态效益功能或植物个体生态效益为主。本研究以临安吴越钱王陵公园为研究对象,运用i-Tree模型对公园植物群落结构及节能、改善空气质量、吸收二氧化碳(CO2)、截留雨水等4个方面的生态效益价值进行定量评估,以期为钱王陵公园及当代城市公园植物群落提升建设过程中群落及树种选择提供一定参考,丰富该模型在城市绿地生态效益研究的理论与内容。
-
临安区位于杭州市西部,29°56'~30°23'N,118°51'~119°52'E,属亚热带季风气候,年平均降水量1 613.90 mm,降水日158.0 d,全年平均气温16.4 ℃,全年日照时数1 847.3 h。钱王陵公园坐落在临安区锦城太庙山南坡,2018年作为杭州地区唯一代表入选浙江省级考古遗址公园,在塑造城市形象、传承历史文化、市民公共游憩活动等方面起重要作用;该园建成至今已有20余a,占地10.30 hm2,海拔92.0 m[12]。园内植物生长稳定,群落结构水平和垂直结构差异明显,群落景观多样,类型丰富。
-
本研究调查对象为钱王陵公园内的典型人工植物群落。在对公园全面勘查的基础上,于2019年8−9月采用典型取样法,设置32个20 m×20 m,面积为400 m2的典型样地,按照L1至L32顺序对样地编号(表1);采用每木测量法,对样地内的植物种类、数量、胸径、树高、冠幅、健康状况以及立地条件、水体、道路、铺装广场的占比状况进行调查[5],后期整理汇总。
表 1 钱王陵公园植物群落基本类型
Table 1. Basic types of plant communities in Qianwangling Park
群落类型 垂直结构类型 样地编号(群落名称) 针阔混交型 乔-灌-草 L1(水杉Metasequoia glyptostroboides+樟树群落)、L2(圆柏Sabina chinensis+樟树群落)、L3(雪松 Cedrus deodara+樟树群落)、L4(塔柏Sabina chinensis ‘Pyramidalis’+圆柏+樟树群落)、L5(塔 柏+樟树群落)、L6(塔柏+樟树+棕榈Trachycarpus fortunei群落)、L9(桂花Osmanthus fragrans+水 杉群落)、L11(水杉+柳杉Cryptomeria fortunei+樟树群落)、L14(水杉+枫香Liquidambar formosana群落)、L18(樟树+罗汉松Podocarpus macrophyllus群落) 乔-灌 L7(水杉+广玉兰群落Magnolia grandiflora)、L17(马尾松Pinus massoniana+深山含笑Michelia maudiae群落) 乔-草 L25(马尾松+樟树群落) 常绿阔叶型 乔-灌-草 L8(桂花群落)、L13(樟树+广玉兰+桂花群落)、L16(樟树+广玉兰群落)、L21(樟树+广玉兰-山茶 Camellia japonical群落)、L26(樟树+桂花群落)、L30(樟树+女贞Ligustrum lucidum+桂花群落) 乔-草 L23(樟树+女贞群落)、L28(樟树+棕榈群落) 常绿落叶阔叶混交型 乔-灌-草 L10(樟树+山樱花Prunus serrulata群落)、L15(樟树+银杏Ginkgo biloba群落)、L26(樟树+枫香+ 构树Broussonetia papyrifera群落(L20) 乔-草 L19(樟树+枫香+油茶Camellia oleifera群落)、L22(樟树+枫香+棕榈群落)、L24(樟树+枫香群落)、 L27(樟树+栓皮栎Quercus variabilis群落) 乔-灌 L12(樟树+榔榆Ulmus parvifolia群落) 竹阔混交型 乔-灌-草 L29(毛竹Phyllostachys edulis+枫香群落)、L32(毛竹+樟树-山茶群落)、L31(毛竹+樟树群落) -
评估采用i-Tree V5.0版本的streets模块。将模型内测算树木节能效益的电费和天然气经济指标价格修正为2018年临安区居民生活用电的平均价格0.59元·kW−1·h−1和天然气平均价格3.66元·m−3;碳吸收、改善空气质量、截留雨水效益的计算参照魏云龙[10]基于该模型同一模块对花港观鱼公园生态效益测算价格指标,用于导入及输出的美元效益价值量采用2019年8月及9月美元对人民币平均汇率(1美元兑换7.04元人民币)进行换算[13]。基于临安亚热带季风气候条件及杭州地区该模型相关研究[5, 10],匹配沿海平面(coastal plain)气候区作为背景开展评估,对树种数据库中没有的树种依据其属性进行同属或同叶型的树种匹配,最后将整理好的树种实测数据整理成Access数据库,并处理成mdb格式文件导入模型中开展评估。
-
参照《中国植被》[14]、尚玉昌[15]生态学优势种命名法、武文婷[16]对于杭州城市绿地植被类型划分原则,以植物群落本身特征作为划分依据,以生态外貌为高级分类单位,中低级单位采用优势种(以重要值为依据),对所调查植物群落进行划分得出(表1):竹阔混交型群落占比为9.37%,常绿落叶阔叶混交型和常绿阔叶型群落占比同为25.00%,针阔混交型群落占比最高(40.63%);公园整体表现出以针阔混交型群落为主的植物景观风貌,营造出纪念性公园四季常青、庄严肃穆的景观氛围。
根据水平结构上的配置模式,将群落划分为纯林和混交林2种类型[17]。在水平结构上,混交林占比为93.75%,呈现出主导地位;在垂直结构上,“乔-灌-草”型复层模式群落占68.75%;双层模式群落只出现“乔-草”及“乔-灌”这2种类型,分别占21.88%和9.37%。复层型为公园植物群落的主要构建模式。
-
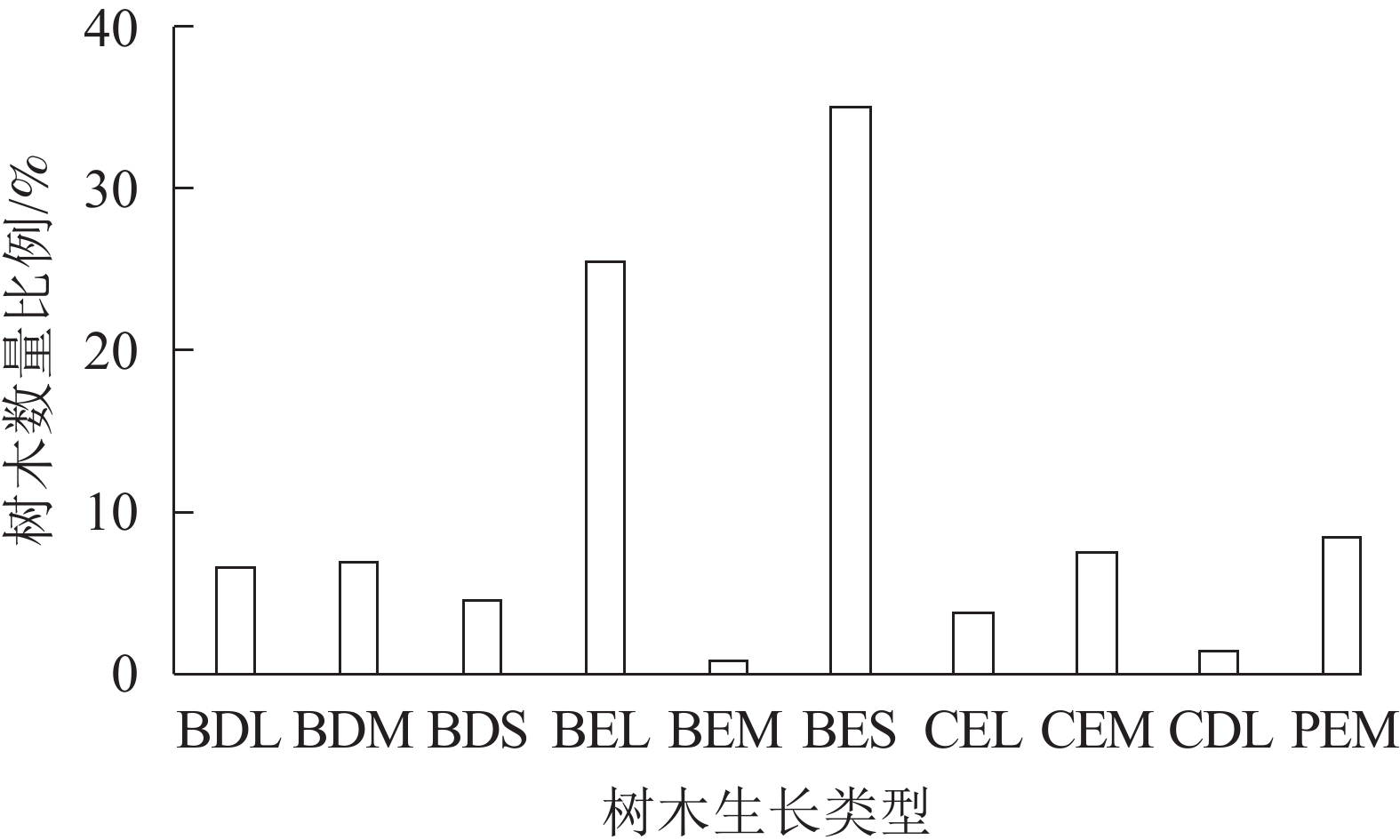
参照《浙江植物志》等[18-19],对所调查32个植物群落树种分析可知:调查植物共计113种,分属于63科98属。符合测量评估条件的乔、灌木共1 613株,隶属于39科62属74种。i-Tree模型内置的“Define Species”功能将树木分为常绿阔叶树种(BE)、落叶阔叶树种(BD)、常绿针叶树种(CE)、落叶针叶树种(CD)、常绿棕榈树种(PE)5种生长类型,且每一类型又分为大(L)、中(M)、小(S)3类[10]。由图1可知:公园内常绿树占比约是落叶树占比的4倍,群落总体表现出以常绿树为主的林相,季相变化不够丰富;公园内阔叶树种占79.11%,针叶树种占12.46%,阔叶树约为针叶树6倍,针叶树种数量偏少,不符合纪念性公园以针叶树偏多的特点。

图 1 植物群落树木生长类型
Figure 1. Plant community tree growth types
从数量上来看,株数排名前10位的树种分别是:樟树(22.13%)、毛竹(10.17%)、桂花(9.92%)、棕榈(8.43%)、枫香(6.01%)、山茶(4.03%)、圆柏(3.72%)、广玉兰(2.60%)、罗汉松(2.60%)、女贞(2.17%);这10个树种数量占调查总数的71.79%;但仅有樟树、桂花、棕榈这3种乡土树种应用频度在50%以上,构成整个公园的优势树种。总体来看,钱王陵公园树种丰富,但绝大多数乔、灌木应用频度低、景观单调。
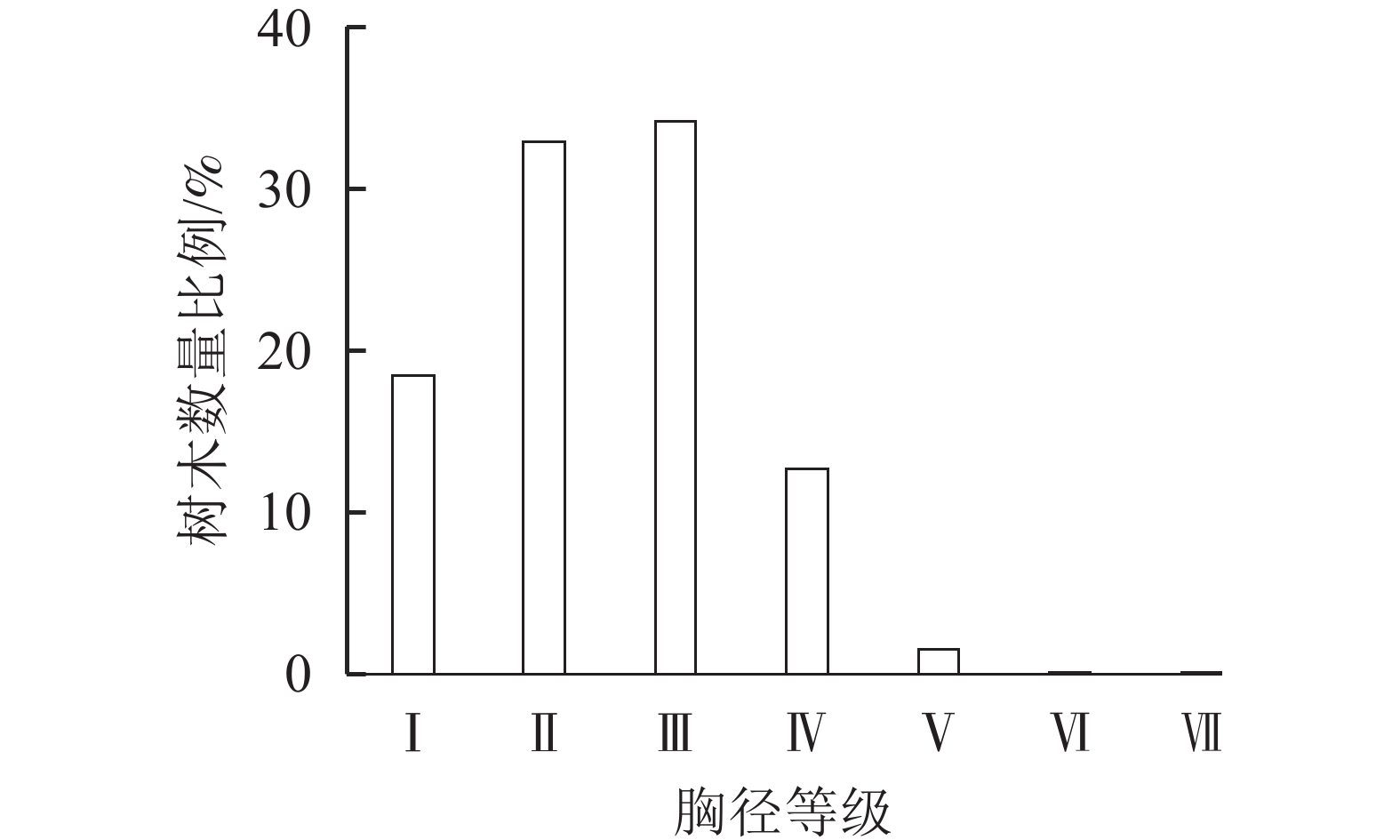
从胸径结构来看,i-Tree模型主要将树种胸径划分为7个等级。由图2可知所调查群落树种胸径主要集分布在≤45.7 cm,胸径在45.70~61.00 cm的占比1.55%,胸径在61.00~76.20 cm和76.20~94.10 cm的占比都为0.06%。公园内树种胸径呈现以中小径级为主,且随着胸径增大树种数量显著减少现象。
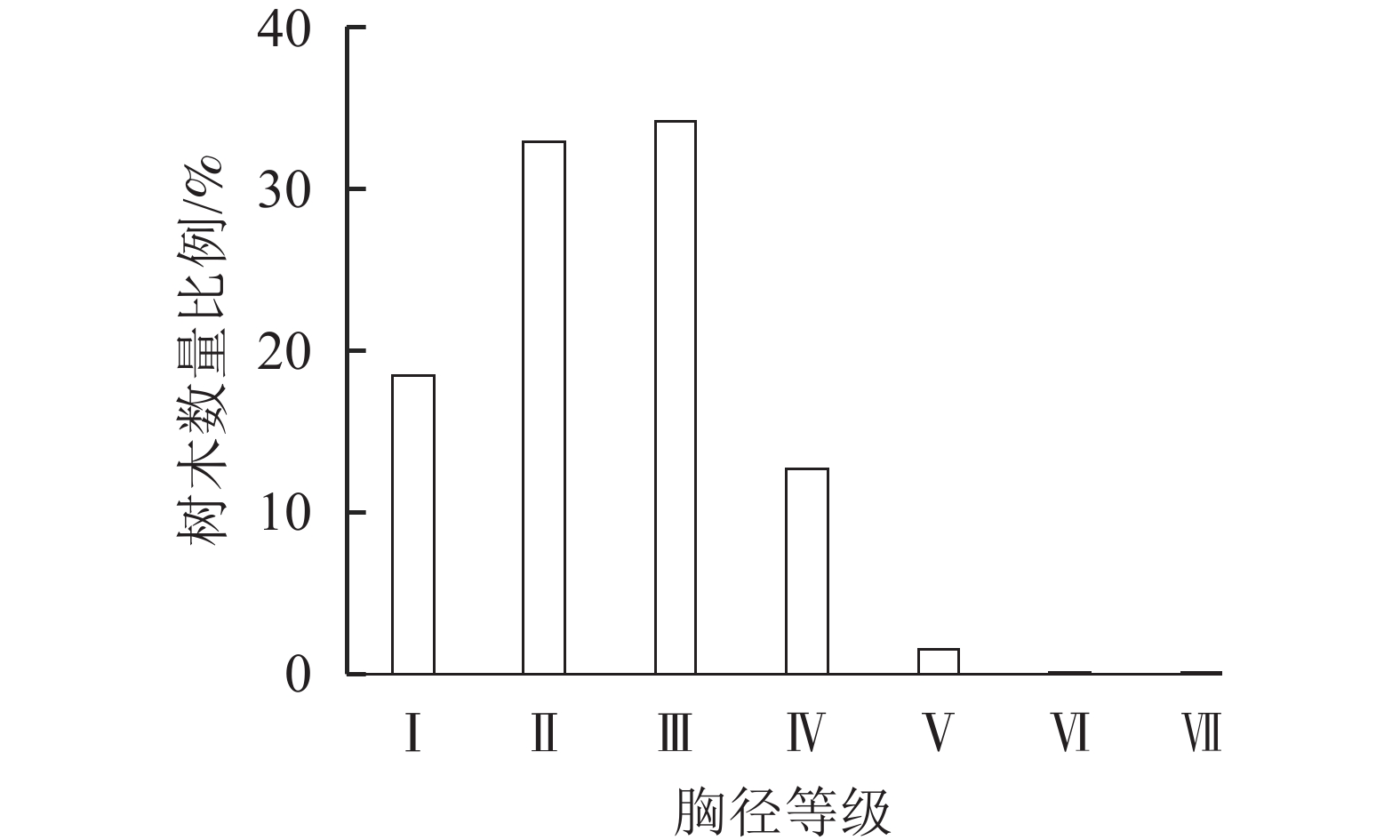
图 2 植物群落树种胸径等级
Figure 2. Plant community tree species’ diameter at breast height grades
-
树木通过遮光、降低风速、叶片蒸腾散热等方式,起到改善小气候、节约能源的作用[20];光合、呼吸等作用吸收CO2和水转化为有机物并释放氧气,实现碳氧循环减缓温室效应[21];通过树冠、树叶、树皮表面对雨水进行拦截,延缓暴雨峰值,起到减少雨水径流的效果[22]。运用i-Tree模型对钱王陵公园32个植物群落节约能源、截留雨水、吸收CO2和改善空气质量生态效益价值定量评估得出(表2):4类生态效益年总价值为208 588.16元·a−1,平均单株生态效益为129.25元·株−1·a−1。对生态效益价值贡献最大的是吸收CO2,占47.20%;节约能源效益与截留雨水效益价值相近,分别占23.10%和27.77%;改善空气质量效益最低,仅占1.93%。这4类生态效益按照价值量从高到低排名依次为吸收CO2、截留雨水、节约能源、改善空气质量。生态效益最高的是樟树+广玉兰-山茶群落(L21),为12 777.60元·a−1,占6.13%;最低的是樟树+银杏群落(L15),为1 203.84元·a−1,仅占0.58%。不同类型群落生态效益价值存在较大差距,对生态效益贡献最大的为针阔混交型群落,总生态效益占比为35.47%;其次是常绿阔叶型群落,占比为27.61%;常绿落叶阔叶混交型群落占比为26.17%;竹阔混交型最少,仅占10.75%。
表 2 钱王陵公园32个植物群落总生态效益
Table 2. Total ecological benefits of 32 plant communities in Qianwangling Park
群落 生态效益/(元·a−1) 效益
占比/
%群落 生态效益/(元·a−1) 效益
占比/
%节约
能源吸收
CO2改善空
气质量截留
雨水总生态
效益节约
能源吸收
CO2改善空
气质量截留
雨水总生态
效益L1 1 070.08 2 168.32 98.56 1 351.68 4 688.64 2.25 L18 1 830.40 3 365.12 28.16 2 400.64 7 624.32 3.66 L2 1 520.64 3 182.08 183.04 1 879.68 6 765.44 3.24 L19 1 457.28 2 872.32 49.28 1 696.64 6 075.52 2.91 L3 1 267.20 2 696.32 133.76 1 591.04 5 688.32 2.73 L20 1 584.00 2 977.92 112.64 1 809.28 6 483.84 3.11 L4 1 112.32 2 393.60 91.52 1 337.60 4 935.04 2.37 L21 2 851.20 5 850.24 232.32 3 843.84 12 777.60 6.13 L5 781.44 1 605.12 42.24 851.84 3 280.64 1.57 L22 2 175.36 4 238.08 126.72 2 745.60 9 285.76 4.45 L6 1 041.92 2 154.24 112.64 1 203.84 4 512.64 2.16 L23 1 802.24 3 808.64 161.92 2 259.84 8 032.64 3.85 L7 1 415.04 3 196.16 154.88 1 675.52 6 441.60 3.09 L24 2 245.76 4 540.80 112.64 2 520.32 9 419.52 4.52 L8 711.04 1 921.92 126.72 682.88 3 442.56 1.65 L25 1 457.28 2 949.76 112.64 1 619.20 6 138.88 2.94 L9 1 619.20 4 026.88 168.96 1 900.80 7 715.84 3.70 L26 1 760.00 3 597.44 161.92 2 316.16 7 835.52 3.76 L10 1 344.64 3 238.40 168.96 1 415.04 6 167.04 2.96 L27 2 696.32 5 005.44 253.44 3 182.08 11 137.28 5.34 L11 1 422.08 3 048.32 35.20 1 774.08 6 279.68 3.01 L28 2 041.60 4 055.04 140.80 2 907.52 9 144.96 4.38 L12 1 119.36 2 358.40 126.72 1 217.92 4 822.40 2.31 L29 1 809.28 3 013.12 197.12 1 752.96 6 772.48 3.25 L13 675.84 1 492.48 77.44 830.72 3 076.48 1.47 L30 1 506.56 3 104.64 84.48 1 844.48 6 540.16 3.14 L14 732.16 1 555.84 84.48 725.12 3 097.60 1.49 L31 1 774.08 3 372.16 133.76 2 337.28 7 617.28 3.65 L15 260.48 577.28 28.16 337.92 1 203.84 0.58 L32 2 062.72 3 548.16 239.36 2 189.44 8 039.68 3.85 L16 1 520.64 3 160.96 147.84 1 907.84 6 737.28 3.23 总计 48 188.80 98 447.36 4 019.84 57 932.16 208 588.16 100 L17 1 520.64 3 358.08 98.56 1 837.44 6 814.72 3.27 对群落各生态效益分析可知:i-Tree模型对节能效益的评估由节约电量和节约天然气效益两部分组成,其中节约电量为62 227.20 kW·h·a−1,产生生态效益为36 706.56元·a−1;节约天然气量3 560.50 m³·a−1,产生生态效益11 482.24元·a−1,节约能源的生态效益总价值为48 188.80元·a−1,单株树木平均生态效益为29.85元·株−1·a−1;节能效益最高的是樟树+广玉兰-山茶群落(L21),为2 851.20元·a−1;最低的是樟树+银杏群落(L15),为260.48元·a−1;从群落类型来看,节能效益总量占比最高的是针阔混交型群落,为34.84%,该类群落在节能效益价值方面优势较高。经模型计算植物群落吸收CO2总量为81 951.00 kg·a−1,产生生态效益价值98 447.36元·a−1,单株平均吸收CO2生态效益为61.04元·株−1·a−1;对不同类型群落生态效益分析可知:针阔混交型群落生态效益占比最高,为36.26%,在吸收CO2方面表现出一定优势。模型计算群落年截留雨水量为5 192.00 m³·a−1,产生生态效益为57 932.16元·a−1,单株树木平均截留雨水生态效益为35.90元·株−1·a−1。对单株树木平均截留雨水生态效益分析可以得出,针阔混交型群落中树木平均效益超过总平均生态效益的群落数量最多且效益排名靠前,说明相对其他类型该群落类型在截留雨水上具有优势。模型计算植物群落年改善空气质量生态效益为4 019.84元·a−1,单株树木平均生态效益为2.46元·株−1·a−1,明显低于以上3个功能,其中群落间接减排空气污染物[二氧化氮(NO2)、可吸入颗粒物(PM10)、挥发性有机化合物(VOC)、二氧化硫(SO2)]生态效益最高,为6 188.16元·a−1,SO2减排量最高为220.40 kg·a−1,占66.85%;群落吸附沉积空气污染物[NO2、臭氧(O3)、PM10和SO2]效益为2 731.52元·a−1,对O3的沉积效果最强,占比为58.65%;SO2的沉积效果最差仅6.14%。此外,树木自身代谢等作用会散发挥发性有机物污染(BVOCs)对空气造成污染[23],经计算该公园群落释放BVOCs量为213.20 kg·a−1,产生生态效益−4 899.84 元·a−1,其中樟树+广玉兰-山茶群落(L21)释放量最多,为13.30 kg·a−1;从改善空气质量综合能力来看,樟树+栓皮栎群落(L27)效果最强,产生生态效益最高,为253.44元·a−1,占6.30%,同时间接减排空气污染物的能力也最为显著。综合看来,树木在间接减排空气污染物方面上更具优势。
对群落树种单株平均生态效益分析可知,排名前10位树种为落叶阔叶树和针叶树(表3),针叶树有水杉、雪松、马尾松,其余的为落叶阔叶树。常绿阔叶树种未出现,生态效益偏低。综合树种平均胸径及各生态效益分析可知,在平均胸径相近情况下,落叶阔叶树各项生态效益高于针叶树,即落叶阔叶树在节能、吸收CO2、改善空气质量及截留雨水方面都表现出更高优势。单株平均生态效益价值由高到低排名前10位的树种为三角枫Acer buergerianum、黄山栾树Koelreuteria paniculata、水杉、栓皮栎、雪松、马尾松、山樱花、朴树Celtis bungeana、榔榆、垂柳Salix babylonica,在植物群落构建时对于落叶阔叶树和针叶树树种的选择可优先考虑。
表 3 单株平均效益前10位树种各生态效益
Table 3. Per plant benefit,top ten tree species, ecological benefits
效益排名 树种名称 生长类型 单株生态效益/(元·株−1·a−1) 单株平均生态效益/
(元·株−1·a−1)平均胸径/cm 节约能源 吸收CO2 改善空气质量 截留雨水 1 三角枫 BDM(落叶阔叶中型树) 119.82 300.40 18.66 119.33 558.21 35.04 2 黄山栾树 BDM(落叶阔叶中型树) 97.72 213.73 16.12 100.95 428.52 30.26 3 水杉 CDL(落叶针叶大型树) 95.46 203.03 −1.76 119.47 416.20 41.47 4 栓皮栎 BDL(落叶阔叶大型树) 79.20 177.76 12.88 76.03 345.87 37.05 5 雪松 CEL(常绿针叶大型树) 76.88 163.40 2.60 82.09 324.97 30.33 6 马尾松 CEL(常绿针叶大型树) 75.82 161.36 1.55 84.83 323.56 33.65 7 山樱花 BDM(落叶阔叶中型树) 69.98 161.43 11.26 66.81 309.48 26.50 8 朴树 BDL(落叶阔叶大型树) 79.62 130.17 13.38 78.14 301.31 27.44 9 榔榆 BDM(落叶阔叶中型树) 63.15 148.54 10.14 60.19 282.02 24.48 10 垂柳 BDM(落叶阔叶中型树) 49.63 122.78 7.88 46.96 227.25 26.48 结合表3和表4分析可知:L21群落(樟树+广玉兰-山茶群落)树种株数占比不高(排第10位),但群落平均胸径最高,单株生态效益高的落叶阔叶树种三角枫、栓皮栎和针叶树马尾松平均胸径都较高。而L22群落(樟树+枫香+棕榈群落)树种株数占比较低,但群落平均胸径高,因此生态效益也相对较高。L24群落(樟树+枫香群落)、L27群落(樟树+栓皮栎群落)和L28群落(樟树+棕榈群落)树种株数占比相似(分别为4.4%、4.0%和4.3%),而群落L27和群落L28相比,群落整体平均胸径相近,但群落L27中单株生态效益高的树木数量及平均胸径更具有优势,因此该群落效益更高;L24群落平均胸径偏低,但单株平均效益高树种株株多,因此生态效益也较高。在树种生长型方面,胸径>30 cm的落叶阔叶和针叶高生态效益树种株数占比越高对生态效益贡献越大。综上分析,群落生态效益的高低主要取决于树种的株数、生长类型和平均胸径。从构建高生态效益植物群落角度出发,建议植物群落构建时以三角枫、黄山栾树、水杉、栓皮栎、雪松、马尾松等高单株生态效益的树木为优势种,以提高植物群落整体生态效益。
表 4 生态效益排名前5位群落指标分析
Table 4. Analysis of the top five community indicators in benefit ranking
排名 样地
编号群落
类型主要组
成树种单株生态效益排名前10树种
(株数及平均胸径)群落整体平均
胸径/(cm·株−1)群落树种株数
占比/%(排名)1 L21 常绿阔叶型 樟树+广玉兰+棕榈-山茶 三角枫(4株,胸径35.06 cm)、栓皮栎(1株,
胸径45.39 cm)、马尾松(2株,胸径39.33 cm)25.70 3.6(10) 2 L27 常绿落叶阔叶
混交型樟树+栓皮栎+女贞+棕榈+
黄山栾树+枫香黄山栾树(3株,胸径29.47 cm)、朴树(1株,
胸径32.16 cm)、栓皮栎(10株,胸径30.86 cm)20.56 4.0(5) 3 L24 常绿落叶阔叶
混交型枫香+樟树+棕榈 栓皮栎(3株,胸径16.73 cm)、马尾松(2株,
胸径36.22 cm)17.96 4.4(3) 4 L22 常绿落叶阔叶
混交型枫香+樟树+棕榈-山茶 马尾松(2株,胸径34.00 cm) 25.31 2.7(19) 5 L28 常绿阔叶型 樟树+女贞+棕榈 马尾松(1株,胸径42.61 cm)、三角枫(1株,
胸径34.98 cm)、雪松(1株,胸径28.83 cm)20.03 4.3(4) -
本研究采用典型取样法及每木调查法对钱王陵公园32个植物群落内所有符合测量条件的乔、灌木进行实地调查。调查树木共计1 613株,分属39科62属74种。群落树种种类丰富,胸径结构分布合理,呈现出较明显上、中、下层分布。在水平结构上以混交型为主,垂直结构以复层模式为主,且表现出以针阔混交型群落为主的景观风貌。公园内存在常绿树、阔叶树数量占比远超落叶树、针叶树,树木生长类型分布不合理,多数灌木及草本植物生长状况不佳,林间凋落物堆积过厚等问题。这与公园建成时间久远缺乏管理与维护有关。从公园长远发展角度考虑,应基于该公园的纪念性特点,增加落叶树与松柏类针叶树种数和株数,营造庄重、肃穆的纪念氛围,以构建更加完善和稳定的植物群落结构体系。
钱王陵公园32个植物群落年总生态效益为208 588.16元·a−1,节约能源、吸收CO2、改善空气质量和截留雨水生态效益价值分别为48 188.80、98 447.36、4 019.84和57 932.16元·a−1,占比分别为23.10%、47.20%、1.93%和27.77%。这4类效益按照价值量高低排名为吸收CO2、截留雨水、节约能源、改善空气质量。结果与魏云龙[10]研究排名一致,其中吸收CO2生态效益最高,贡献最大。这与树种本身对CO2吸收能力较强有关。不同类型群落生态效益存在明显差异,针阔混交型群落总生态效益最高,占38.38%,表现出显著优势。这是由于该类型群落数量(占38.38%)及树种株数(占35.30%)最多,且水杉、马尾松、雪松等单株平均生态效益高的树种占比大。其次分别为常绿阔叶型、常绿落叶阔叶混交型和竹阔混交型群落;常绿阔叶型群落数量与常绿落叶阔叶混交型群落数量相同,占比为25%。常绿阔叶型群落总生态效益占比为27.61%,树种株数占24.1%;常绿落叶阔叶混交型群落总生态效益占比为26.71%及树种株数占26.20%。竹阔混交型群落在群落数量(占9.37%)及树种株数(占14.5%)上都为最少,导致生态效益占比最低为10.75%。
在城市绿地的建设过程中,i-Tree模型能从资源结构及生态效益方面对植物群落进行定量评估,将不同群落和树种的生态效益以货币形式更直观地展示出来,若将其引入城市绿地植物群落构建的“人工形成—自然进程—人工调控”的动态过程中,将使高生态效益绿地的建设更加具有科学性、高效性和便捷性。由于该模型是基于美国的生态环境而开发的,应用于本研究评估时在树种匹配上有一定的局限性,因此对于国内树种的研究以及完善更新模型基础数据库,使之能够更好地适用于本土树种,是今后重点研究方向。在生态服务功能评估上,由于i-Tree模型限制,只从节约能源、吸收CO2、改善空气质量、截留雨水等4个方面对钱王陵公园生态服务功能价值进行评估。植物群落生态服务功能的种类众多,还包括释放氧气、改良土壤及减少空气中负离子等,因此还需从深度和广度上加以进一步完善和提高。在成本参数上,模型中需要输入清除空气中O3、PM10和VOC等成本参数,因国内未有统一标准,参数获取难度大。在参照国内该模型的相关研究基础上,使用了模型的内置参数进行评估。在以后的研究中,应该建立符合国内情况的处理空气污染物和截留雨水相关参数指标,从而更加完善地运用评估模型对生态效益价值进行评估研究。
Plant community structure and ecological benefits of Qianwangling Park in Lin’an
-
摘要:
目的 分析浙江省杭州市临安区钱王陵公园植物群落结构,并对其生态效益进行定量评估。 方法 以临安钱王陵公园为研究对象,对公园内32个植物群落资源结构进行实地调查,运用i-Tree模型评估群落节能、吸收二氧化碳(CO2)、改善空气质量、截留雨水等生态效益。 结果 ①所调查群落树木共计1 613株,分属39科62属74种;群落结构以复层型为主,呈现出以针阔混交林为主的景观风貌。②群落年生态效益为208 588.16元·a−1,单株树木平均效益为129.25元·株−1·a−1,其中节约能源、吸收CO2、改善空气质量和截留雨水生态效益分别为48 188.80、98 447.36、4 019.84和57 932.16元·a−1,分别占为23.10%、47.20%、1.93%和27.77%;生态效益总价值最高的是樟树Cinnamomum camphora+广玉兰Magnolia grandiflora-山茶Camellia japonica群落(样地L21),为12 777.60元·a−1,占6.13%,且该群落在节约能源、吸收CO2、截留雨水3个方面生态效益均为最高;针阔混交型群落生态效益占35.47%,对生态效益价值贡献最大。③落叶阔叶及针叶树种单株平均生态效益较高,落叶阔叶树在节能、吸收CO2、改善空气质量及截留雨水效益方面更具优势。 结论 钱王陵公园植物群落生态效益的高低主要取决于群落内树种的株数、生长类型和平均胸径,针阔混交型群落在生态效益价值方面具有明显优势。图2表4参23 Abstract:Objective This research aims to analyze the plant community structure of Qianwangling Park in Lin’an District of Hangzhou City, Zhejiang Province, and evaluate its ecological benefits. Method Taking Qianwangling Park as the research object, the structure of 32 plant communities in the park was investigated, and the i-Tree model was used to evaluate the ecological benefits of community including energy conservation, carbon dioxide absorption, air quality improvement and rainwater interception. Result (1) There were 1 613 trees in the community, belonging to 39 families 62 genera and 74 species. The community structure was mainly of the multi-layer type, dominated by the landscape style of the mixed forest of needles and broads. (2) The annual ecological benefit of the community was 208 588.16 yuan·a−1, and the average benefit per tree was 129.25 yuan·plant−1·a−1, among which the benefits of energy conservation, carbon dioxide absorption, air quality improvement and rainwater interception were 48 188.80, 98 447.36, 4 019.84 and 57 932.16 yuan·a−1, accounting for 23.10%, 47.20%, 1.93% and 27.77% respectively. The highest total value of ecological benefits reached 12 777.60 yuan·a−1, found in sample L21 (Cinnamomum camphora+Magnolia grandiflora-Camellia japonica community), accounting for 6.13%. This community had the highest benefits in saving energy, absorbing carbon dioxide and intercepting rainwater. The mixed community contributed the most to ecological benefit values, accounting for 35.47%. (3) The deciduous broad-leaved and coniferous trees had higher average benefits per plant, and deciduous broad-leaved trees had more advantages in energy conservation, CO2 absorption, air quality improvement and rainwater interception. Conclusion The ecological benefit of plant community in Qianwangling Park is mainly determined by the number of tree species, growth type and average diameter at breast height. The coniferous and broad-leaved mixed community has obvious advantages in ecological benefit value. [Ch, 2 fig. 4 tab. 23 ref.] -
Key words:
- forest ecology /
- plant community /
- i-Tree model /
- ecological benefit /
- Qianwangling Park
-
图 1 植物群落树木生长类型
BDL. 落叶阔叶大型树;BDM. 落叶阔叶中型树; BDS. 落叶阔叶小型树;BEL. 常绿阔叶大型树; BEM. 常绿阔叶中型树;BES. 常绿阔叶小型树; CEL. 常绿针叶大型树;CEM. 常绿针叶中型树; CDL. 落叶针叶大型树;PEM. 常绿棕榈中型树
Figure 1 Plant community tree growth types
图 2 植物群落树种胸径等级
Ⅰ. DBH≤7.6 cm;Ⅱ. 7.6<DBH≤15.2 cm; Ⅲ. 15.2<DBH≤30.5 cm;Ⅳ. 30.5<DBH≤45.7 cm; Ⅴ. 45.7<DBH≤61.0 cm;Ⅵ. 61.0<DBH≤76.2 cm; Ⅶ. 76.2<DBH≤91.4 cm
Figure 2 Plant community tree species’ diameter at breast height grades
表 1 钱王陵公园植物群落基本类型
Table 1. Basic types of plant communities in Qianwangling Park
群落类型 垂直结构类型 样地编号(群落名称) 针阔混交型 乔-灌-草 L1(水杉Metasequoia glyptostroboides+樟树群落)、L2(圆柏Sabina chinensis+樟树群落)、L3(雪松 Cedrus deodara+樟树群落)、L4(塔柏Sabina chinensis ‘Pyramidalis’+圆柏+樟树群落)、L5(塔 柏+樟树群落)、L6(塔柏+樟树+棕榈Trachycarpus fortunei群落)、L9(桂花Osmanthus fragrans+水 杉群落)、L11(水杉+柳杉Cryptomeria fortunei+樟树群落)、L14(水杉+枫香Liquidambar formosana群落)、L18(樟树+罗汉松Podocarpus macrophyllus群落) 乔-灌 L7(水杉+广玉兰群落Magnolia grandiflora)、L17(马尾松Pinus massoniana+深山含笑Michelia maudiae群落) 乔-草 L25(马尾松+樟树群落) 常绿阔叶型 乔-灌-草 L8(桂花群落)、L13(樟树+广玉兰+桂花群落)、L16(樟树+广玉兰群落)、L21(樟树+广玉兰-山茶 Camellia japonical群落)、L26(樟树+桂花群落)、L30(樟树+女贞Ligustrum lucidum+桂花群落) 乔-草 L23(樟树+女贞群落)、L28(樟树+棕榈群落) 常绿落叶阔叶混交型 乔-灌-草 L10(樟树+山樱花Prunus serrulata群落)、L15(樟树+银杏Ginkgo biloba群落)、L26(樟树+枫香+ 构树Broussonetia papyrifera群落(L20) 乔-草 L19(樟树+枫香+油茶Camellia oleifera群落)、L22(樟树+枫香+棕榈群落)、L24(樟树+枫香群落)、 L27(樟树+栓皮栎Quercus variabilis群落) 乔-灌 L12(樟树+榔榆Ulmus parvifolia群落) 竹阔混交型 乔-灌-草 L29(毛竹Phyllostachys edulis+枫香群落)、L32(毛竹+樟树-山茶群落)、L31(毛竹+樟树群落)  下载: 导出CSV
下载: 导出CSV
表 2 钱王陵公园32个植物群落总生态效益
Table 2. Total ecological benefits of 32 plant communities in Qianwangling Park
群落 生态效益/(元·a−1) 效益
占比/
%群落 生态效益/(元·a−1) 效益
占比/
%节约
能源吸收
CO2改善空
气质量截留
雨水总生态
效益节约
能源吸收
CO2改善空
气质量截留
雨水总生态
效益L1 1 070.08 2 168.32 98.56 1 351.68 4 688.64 2.25 L18 1 830.40 3 365.12 28.16 2 400.64 7 624.32 3.66 L2 1 520.64 3 182.08 183.04 1 879.68 6 765.44 3.24 L19 1 457.28 2 872.32 49.28 1 696.64 6 075.52 2.91 L3 1 267.20 2 696.32 133.76 1 591.04 5 688.32 2.73 L20 1 584.00 2 977.92 112.64 1 809.28 6 483.84 3.11 L4 1 112.32 2 393.60 91.52 1 337.60 4 935.04 2.37 L21 2 851.20 5 850.24 232.32 3 843.84 12 777.60 6.13 L5 781.44 1 605.12 42.24 851.84 3 280.64 1.57 L22 2 175.36 4 238.08 126.72 2 745.60 9 285.76 4.45 L6 1 041.92 2 154.24 112.64 1 203.84 4 512.64 2.16 L23 1 802.24 3 808.64 161.92 2 259.84 8 032.64 3.85 L7 1 415.04 3 196.16 154.88 1 675.52 6 441.60 3.09 L24 2 245.76 4 540.80 112.64 2 520.32 9 419.52 4.52 L8 711.04 1 921.92 126.72 682.88 3 442.56 1.65 L25 1 457.28 2 949.76 112.64 1 619.20 6 138.88 2.94 L9 1 619.20 4 026.88 168.96 1 900.80 7 715.84 3.70 L26 1 760.00 3 597.44 161.92 2 316.16 7 835.52 3.76 L10 1 344.64 3 238.40 168.96 1 415.04 6 167.04 2.96 L27 2 696.32 5 005.44 253.44 3 182.08 11 137.28 5.34 L11 1 422.08 3 048.32 35.20 1 774.08 6 279.68 3.01 L28 2 041.60 4 055.04 140.80 2 907.52 9 144.96 4.38 L12 1 119.36 2 358.40 126.72 1 217.92 4 822.40 2.31 L29 1 809.28 3 013.12 197.12 1 752.96 6 772.48 3.25 L13 675.84 1 492.48 77.44 830.72 3 076.48 1.47 L30 1 506.56 3 104.64 84.48 1 844.48 6 540.16 3.14 L14 732.16 1 555.84 84.48 725.12 3 097.60 1.49 L31 1 774.08 3 372.16 133.76 2 337.28 7 617.28 3.65 L15 260.48 577.28 28.16 337.92 1 203.84 0.58 L32 2 062.72 3 548.16 239.36 2 189.44 8 039.68 3.85 L16 1 520.64 3 160.96 147.84 1 907.84 6 737.28 3.23 总计 48 188.80 98 447.36 4 019.84 57 932.16 208 588.16 100 L17 1 520.64 3 358.08 98.56 1 837.44 6 814.72 3.27
下载: 导出CSV
表 3 单株平均效益前10位树种各生态效益
Table 3. Per plant benefit,top ten tree species, ecological benefits
效益排名 树种名称 生长类型 单株生态效益/(元·株−1·a−1) 单株平均生态效益/
(元·株−1·a−1)平均胸径/cm 节约能源 吸收CO2 改善空气质量 截留雨水 1 三角枫 BDM(落叶阔叶中型树) 119.82 300.40 18.66 119.33 558.21 35.04 2 黄山栾树 BDM(落叶阔叶中型树) 97.72 213.73 16.12 100.95 428.52 30.26 3 水杉 CDL(落叶针叶大型树) 95.46 203.03 −1.76 119.47 416.20 41.47 4 栓皮栎 BDL(落叶阔叶大型树) 79.20 177.76 12.88 76.03 345.87 37.05 5 雪松 CEL(常绿针叶大型树) 76.88 163.40 2.60 82.09 324.97 30.33 6 马尾松 CEL(常绿针叶大型树) 75.82 161.36 1.55 84.83 323.56 33.65 7 山樱花 BDM(落叶阔叶中型树) 69.98 161.43 11.26 66.81 309.48 26.50 8 朴树 BDL(落叶阔叶大型树) 79.62 130.17 13.38 78.14 301.31 27.44 9 榔榆 BDM(落叶阔叶中型树) 63.15 148.54 10.14 60.19 282.02 24.48 10 垂柳 BDM(落叶阔叶中型树) 49.63 122.78 7.88 46.96 227.25 26.48
下载: 导出CSV
表 4 生态效益排名前5位群落指标分析
Table 4. Analysis of the top five community indicators in benefit ranking
排名 样地
编号群落
类型主要组
成树种单株生态效益排名前10树种
(株数及平均胸径)群落整体平均
胸径/(cm·株−1)群落树种株数
占比/%(排名)1 L21 常绿阔叶型 樟树+广玉兰+棕榈-山茶 三角枫(4株,胸径35.06 cm)、栓皮栎(1株,
胸径45.39 cm)、马尾松(2株,胸径39.33 cm)25.70 3.6(10) 2 L27 常绿落叶阔叶
混交型樟树+栓皮栎+女贞+棕榈+
黄山栾树+枫香黄山栾树(3株,胸径29.47 cm)、朴树(1株,
胸径32.16 cm)、栓皮栎(10株,胸径30.86 cm)20.56 4.0(5) 3 L24 常绿落叶阔叶
混交型枫香+樟树+棕榈 栓皮栎(3株,胸径16.73 cm)、马尾松(2株,
胸径36.22 cm)17.96 4.4(3) 4 L22 常绿落叶阔叶
混交型枫香+樟树+棕榈-山茶 马尾松(2株,胸径34.00 cm) 25.31 2.7(19) 5 L28 常绿阔叶型 樟树+女贞+棕榈 马尾松(1株,胸径42.61 cm)、三角枫(1株,
胸径34.98 cm)、雪松(1株,胸径28.83 cm)20.03 4.3(4)
下载: 导出CSV
-
[1] 潘剑彬. 北京奥林匹克森林公园绿地生态效益研究[D]. 北京: 北京林业大学, 2011. PAN Jianbin. Study on the Ecological Benefits of Green Space in Beijing Olympic Forest Park[D]. Beijing: Beijing Forestry University, 2011. [2] 周鹏, 周婷, 彭少麟. 生态系统服务价值测度模式与方法[J]. 生态学报, 2019, 39(15): 5379 − 5388. ZHOU Peng, ZHOU Ting, PENG Shaolin. Measurement modalities and methodologies of ecosystem services valuation [J]. Acta Ecol Sin, 2019, 39(15): 5379 − 5388. [3] 赵军, 杨凯. 生态系统服务价值评估研究进展[J]. 生态学报, 2007, 27(1): 346 − 356. ZHAO Jun, YANG Kai. Research progress on ecosystem service value assessment [J]. Acta Ecol Sin, 2007, 27(1): 346 − 356. [4] WANG Qian, ZHANG Zhongxia, WANG Ping. An assessment of ecosystem services of urban green spaces based on i-Tree [J]. J Landscape Res, 2019, 11(1): 53 − 56. [5] 施健健, 蔡建国, 刘朋朋, 等. 杭州花港观鱼公园森林吸收二氧化碳效益评估[J]. 浙江农林大学学报, 2018, 35(5): 829 − 835. SHI Jianjian, CAI Jianguo, LIU Pengpeng, et al. Evaluation of carbon dioxide absorption in forests of Hangzhou Huagang Guanyu Park [J]. J Zhejiang A&F Univ, 2018, 35(5): 829 − 835. [6] 马宁, 何兴元, 石险峰, 等. 基于i-Tree模型的城市森林经济效益评估[J]. 生态学杂志, 2011, 30(4): 810 − 817. MA Ning, HE Xingyuan, SHI Xianfeng, et al. Urban forest economic benefit evaluation based on i-Tree model [J]. Chin J Ecol, 2011, 30(4): 810 − 817. [7] 陶晓, 吴泽民, 郝焰平. 合肥市行道树生态效益研究[J]. 中国农学通报, 2009, 25(3): 75 − 82. TAO Xiao, WU Zemin, HAO Yanping. Study on ecological benefits of street trees in Hefei City [J]. Chin Agric Sci Bull, 2009, 25(3): 75 − 82. [8] 刘朋朋, 蔡建国, 章毅, 等. 基于i-Tree模型的杭州市环西湖景区行道树生态效益分析[J]. 科技通报, 2019, 35(2): 132 − 137. LIU Pengpeng, CAI Jianguo, ZHANG Yi, et al. Ecological benefit analysis of street trees in Hangzhou West Lake Scenic Area based on i-Tree Model [J]. Bull Sci Technol, 2019, 35(2): 132 − 137. [9] 熊金鑫, 祁慧君, 王倩茹, 等. 基于i-Tree模型的城市小区行道树生态效益评价[J]. 南京林业大学学报(自然科学版), 2019, 43(2): 128 − 136. XIONG Jinxin, YAN Huijun, WANG Qianru, et al. Evaluation of ecological benefits of urban community street trees based on i-Tree model [J]. J Nanjing For Univ Nat Sci Ed, 2019, 43(2): 128 − 136. [10] 魏云龙. 杭州花港观鱼公园生态服务功能价值评估研究[D]. 杭州: 浙江农林大学, 2017. WEI Yunlong. Study on the Evaluation of Ecological Service Function Value of Hangzhou Huagang Guanyu Park[D]. Hangzhou: Zhejiang A&F University, 2017. [11] 施炜婷, 王燕, 陈聃, 等. 基于i-Tree Eco模型的城市绿地生态功能与价值评估: 以常州市民广场为例[J]. 常州工学院学报, 2018, 31(5): 16 − 21. SHI Weiting, WANG Yan, CHEN Dan, et al. Evaluation of urban green space ecological function and value based on i-Tree Eco Model: taking changzhou citizen square as an example [J]. J Changzhou Inst Technol, 2018, 31(5): 16 − 21. [12] 裘国华. 临安钱王陵公园绿化景观的综合改造[J]. 现代园艺, 2018, 40(6): 68. QIU Guohua. The comprehensive transformation of the green landscape of Lin’an Qianwangling Park [J]. Mod Hortic, 2018, 40(6): 68. [13] 中国人民银行. 2019年统计数据: 货币统计概览[EB/OL]. 2019-10-08[2019-10-20]. http://www.pbc.gov.cn/diaochatongjisi/116219/116319/3245697/3245856/index.html. [14] 中国植被编辑委员会. 中国植被[M]. 北京: 科学出版社, 1980. [15] 尚玉昌. 普通生态学[M]. 北京: 北京大学出版社, 2002. [16] 武文婷. 杭州市城市绿地生态服务功能价值评估研究[D]. 南京: 南京林业大学, 2011. WU Wenting. Study on the Evaluation of Urban Green Space Ecological Service Function Value in Hangzhou[D]. Nanjing: Nanjing Forestry University, 2011. [17] 达良俊, 方和俊, 李艳艳. 上海中心城区绿地植物群落多样性诊断和协调性评价[J]. 中国园林, 2008, 24(3): 87 − 90. DA Liangjun, FANG Hejun, LI Yanyan. Diagnosis and coordination evaluation of green space plant community diversity in central district of Shanghai [J]. Chin Landscape Archit, 2008, 24(3): 87 − 90. [18] 浙江植物志编辑委员会. 浙江植物志[M]. 杭州: 浙江科学技术出版社, 1993. [19] 吴征镒, 周浙昆, 李德铢, 等. 世界种子植物科的分布区类型系统[J]. 云南植物研究, 2003, 25(3): 245 − 257. WU Zhengyi, ZHOU Zhekun, LI Dezhen, et al. The arel-types of the world families of seed plants [J]. Acta Bot Yunnan, 2003, 25(3): 245 − 257. [20] DIMOUDI A, NIKOLOPOULOU M. Vegetation in the urban environment: microclimatic analysis and benefits [J]. Energy Build, 2003, 35(1): 69 − 76. [21] NOWAK D J, CRANE D E. Carbon storage and sequestration by urban trees in the USA [J]. Environ Poll, 2002, 36(16): 381 − 389. [22] XIAO Qingfu, MCPHERSON E G, USTIN S L, et al. Winter rainfall interception by two mature open-grown trees in Davis, California [J]. Hydrol Process, 2000, 14(4): 763 − 784. [23] SMIATEK G, STEINBRECHER R. Temporal and spatial variation of forest VOC emissions in Germany in the decade 1994−2003 [J]. Atmos Environ, 2006, 40: 166 − 177. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20190464
 点击查看大图
点击查看大图
计量
- 文章访问数: 2694
- HTML全文浏览量: 1001
- PDF下载量: 153
- 被引次数: 0
