






-
农作物秸秆具有十分巨大的利用潜力和价值,通过制备复合材料等方式成为生物质资源利用的重点方向之一[1-2]。但秸秆表面由硅和蜡质形成的非极性表层结构会影响胶黏剂在秸秆纤维表面的润湿和吸附,无法形成良好的胶合界面,导致复合材料性能下降[3]。为了获得表面性能良好的秸秆纤维原料,增加秸秆纤维与胶黏剂的相容性,需要对秸秆纤维进行表面改性处理。常用的改性处理方式有物理、化学和生物改性等3种[4],相比之下,生物改性更加环保,但目前应用较少,发展潜力巨大[5-6]。秸秆纤维的生物改性方法包括酶处理和微生物发酵等[7],其中酶处理法用到的生物酶较多,如纤维素酶、木聚糖酶、果胶酶以及降解木质素的氧化酶等[8],但处理过程中释放的糖易被内源性微生物迅速消耗;生物发酵利用某些微生物的消耗分解作用,可得到表面性能良好的秸秆纤维,但通常需要数周,历时较长。QU等[9]使用微生物接种剂,包括枯草芽孢杆菌Bacillus subtilis、地衣芽孢杆菌Bacillus licheniformis、罗氏链霉菌Streptomyces gougerotii和黑曲霉Aspergillus niger,发酵改性小麦Triticum aestivum秸秆,降低了小麦秸秆纤维的长度和宽度,去除秸秆表面蜡质,提高了纤维结晶度。LI等[10]对比了碱处理和酶处理秸秆纤维对秸秆/高密度聚乙烯复合材料的影响,结果表明碱和酶处理均提高了复合材料的界面性能,界面附着力良好。XU等[11]利用不同的酶(纤维素酶、半纤维素酶、漆酶和蛋白酶)及不同的发酵方式(好氧发酵、混合厌氧发酵和浸泡厌氧发酵)对小麦秸秆进行组合处理,结果表明处理之后的小麦秸秆表面蜡质脱落,秸秆纤维的物理结构发生改变,结晶度均大于对照组。可见,生物改性预处理可有效改善纤维与聚酯间的界面相容性,提高秸秆基复合材料的力学性能和热稳定性,在减少环境污染、改善秸秆纤维性能以及提高木质纤维素生物质的综合利用等方面具有广阔的发展前景[12-14]。本研究利用微生物发酵改性秸秆,并将改性秸秆纤维与脲醛树脂混合制备复合材料,探讨了微生物发酵时长对秸秆纤维及制备的复合材料的性能的影响,为秸秆基复合材料的制备应用及发展提供理论依据。
-
水稻Oryza sativa秸秆取自江苏省农业科学院农业资源与环境研究所,风干后(含水率9.17%)粉碎至1~2 cm备用;微生物菌剂(秸秆腐熟剂)购买于南京宁粮生物工程有限公司,含有枯草芽孢杆菌、酵母Saccharomyces、粪肠球菌Enterococcus faecalis、蛋白酶、纤维素酶等。
-
30 kg水稻秸秆接种545 g微生物菌剂(微生物菌剂质量为秸秆绝干质量的2%),调节体系的碳氮比(C/N)为30∶1,含水率为65%,混合均匀后均分转移至自制生物发酵箱中(尺寸为1 m×1 m×1 m,3层箱壁结构,内、外壁为铁质,之间填充有保温泡沫),在30 ℃以上室温环境内静置培养,定期翻动物料以保证充足的新鲜空气。发酵至第5天,从各发酵箱中取出一半物料混合均匀后作为生物改性处理5 d的秸秆纤维,记为S5,剩余物料继续发酵至第10天后全部取出,混合均匀作为生物改性处理10 d的秸秆纤维,记为S10。未生物改性秸秆纤维记为S0。
-
采用“碱-酸-碱”三段制备工艺制备脲醛树脂,3个阶段甲醛与尿素的摩尔比分别为2.0∶1.0,1.5∶1.0和1.4∶1.0,第3阶段另加入占尿素总质量20%的聚乙二醇二缩水甘油醚(PEGDGE)进行增韧改性。物料及添加质量见表1。制得的脲醛树脂黏度为315.6 mPa·s,固含量为54.18%,游离甲醛含量(质量分数)为0.38%,pH 8.30。
表 1 脲醛树脂制备过程中物料添加情况
Table 1. Adding quality of materials during the preparation of urea-formaldehyde resin
阶段 37%(质量分数)
甲醛溶液/g尿素/g 聚乙烯
醇-124/g三聚氰
胺/gPEGDGE/g 1 1 000.0 371.0 5.3 2 106.0 10.6 3 53.0 106.0 将脲醛树脂与秸秆纤维按照20%的施胶量混合均匀(施胶量为脲醛树脂固体质量占复合材料总质量的比),使用平板硫化机(QLB-D 400×400×2,上海第一橡胶机械厂,中国)压制秸秆纤维复合材料,热压工艺为温度130 ℃,时间3 min,压力3.0 MPa,尺寸规格300.0 mm×300.0 mm×3.0 mm (长×宽×厚),密度为(0.80±0.05) g·cm−3。热压完成后将纤维板陈放48 h以平衡含水率并释放内部应力。秸秆S0、S5和S10制备的复合材料分别记为F0、F5和F10。
-
采用范式洗涤纤维测定法[15]测定生物改性秸秆的半纤维素、纤维素、酸不溶木质素以及灰分的变化,将20~40目的秸秆纤维使用纤维测定仪(F800,山东海能科学仪器有限公司,中国)先后经中性洗涤剂和酸性洗涤剂消煮后,使用质量分数为72%硫酸溶液浸没2 h以上,最后使用马弗炉灼烧,测定计算生物改性秸秆纤维组分变化。
-
使用XRD-D2PHASER型X射线衍射仪(Bruker AXS,德国)测定样品晶体结构变化,铜靶,衍射范围为5°~80°,测试步长为0.02。结晶度(ICr)计算方法为ICr=(1−Iam/I002)×100%。其中:I002是纤维素在结晶相的衍射强度(2θ)为22°~23°时结晶强度的最大值;Iam是在非晶相的衍射强度(2θ)为17°~18°时结晶强度的最小值。
-
在10 kV加速电压下采用扫描电子显微镜(SEM,EVO-LS10,Carl Zeiss Jena,德国)观察生物改性秸秆样品的表面微观结构变化。
-
使用JC2000D5型接触角测量仪(上海中晨数字技术设备有限公司,中国)测量水滴(极性液体)在复合材料表面的接触角变化,每间隔30 s拍照测量1次,测试总时长为10 min;使用相同方法测量二碘甲烷(非极性液体)在复合材料表面的接触角变化,然后利用Owens二液法[16]计算复合材料的表面能。
-
参照GB/T 17657—2013《人造板及饰面人造板理化性能试验方法》,采用三点弯曲方法,利用微机控制电子万能力学试验机(CMT4304,深圳新三思计量技术有限公司,中国)测量计算复合材料的抗弯强度(MOR)、弹性模量(MOE),加载速率 2 mm·min−1。每组测量5个试样。
-
参照GB/T 17657—2013《人造板及饰面人造板理化性能试验方法》,使用塑料摆冲击试验机(501J,深圳万测试验设备有限公司,中国)测量计算复合材料的冲击韧性,摆锤能量为2 J。每组测量10个试样。
-
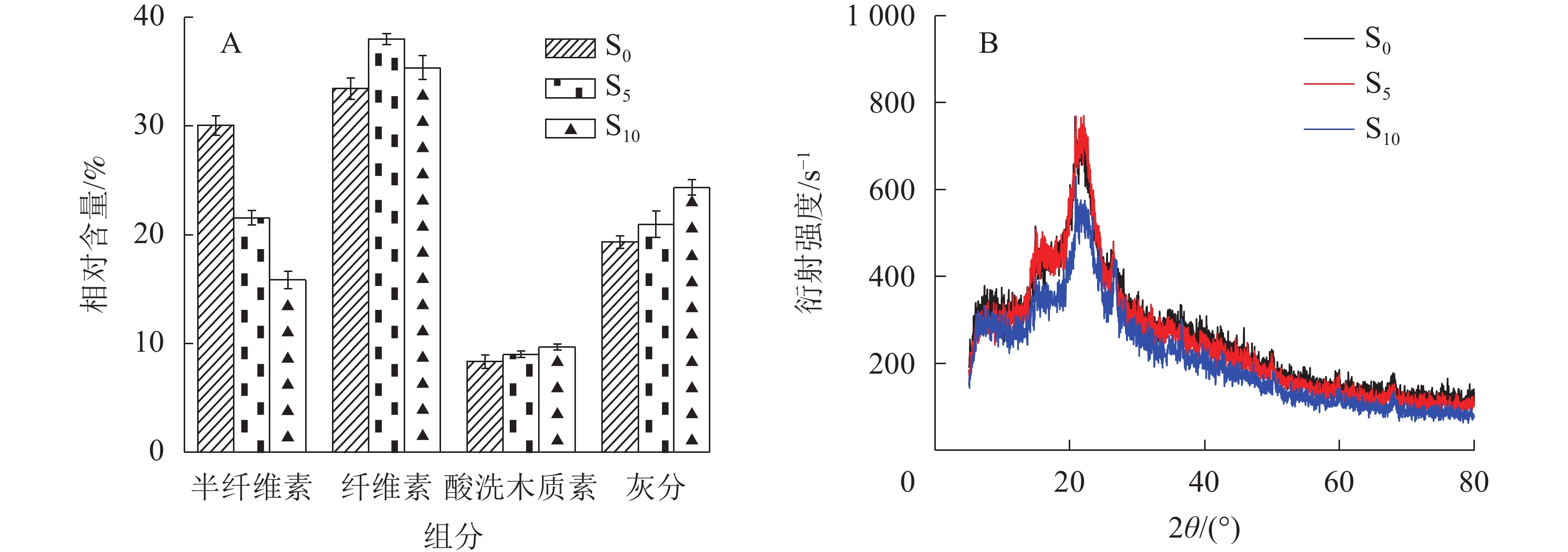
由图1A可知:随着生物改性处理时间延长,秸秆纤维的半纤维素相对含量逐渐降低。这是因为半纤维素在天然状态下聚合度低,化学活性较强,在生物改性时更易降解。纤维素相对含量随生物改性处理时间延长先提高后降低,是因为纤维素结构存在不定形区和定形区,定形区结晶度较好不易降解,生物降解速度低于半纤维素[17]。生物改性处理前期,半纤维素快速而大量地降解,纤维素相对含量提高,而后由于半纤维素相对含量较低,微生物消耗纤维素的速率加快,纤维素的不定形区首先开始降解,随后定形区降解,相对含量开始下降。木质素与灰分为相对不易降解的物质[18],因此在10 d的生物改性处理时间内秸秆纤维的木质素和灰分的相对含量逐渐增加。
由图1B可知:S0、S5和S10的结晶度分别为43.6%、47.8%和41.4%。在微生物作用下,半纤维素及纤维素不定形区在改性处理前期优先降解,纤维素定形区占比增加,聚合度提高,因此在生物改性处理前期S5的结晶度相比于S0有较大幅度提高。随着生物改性处理时间的延长,半纤维素含量逐渐降低,微生物开始大量分解并消耗纤维素,S10的结晶度因此降低,甚至低于S0,可见生物改性处理10 d后秸秆纤维素定形区部分分解。
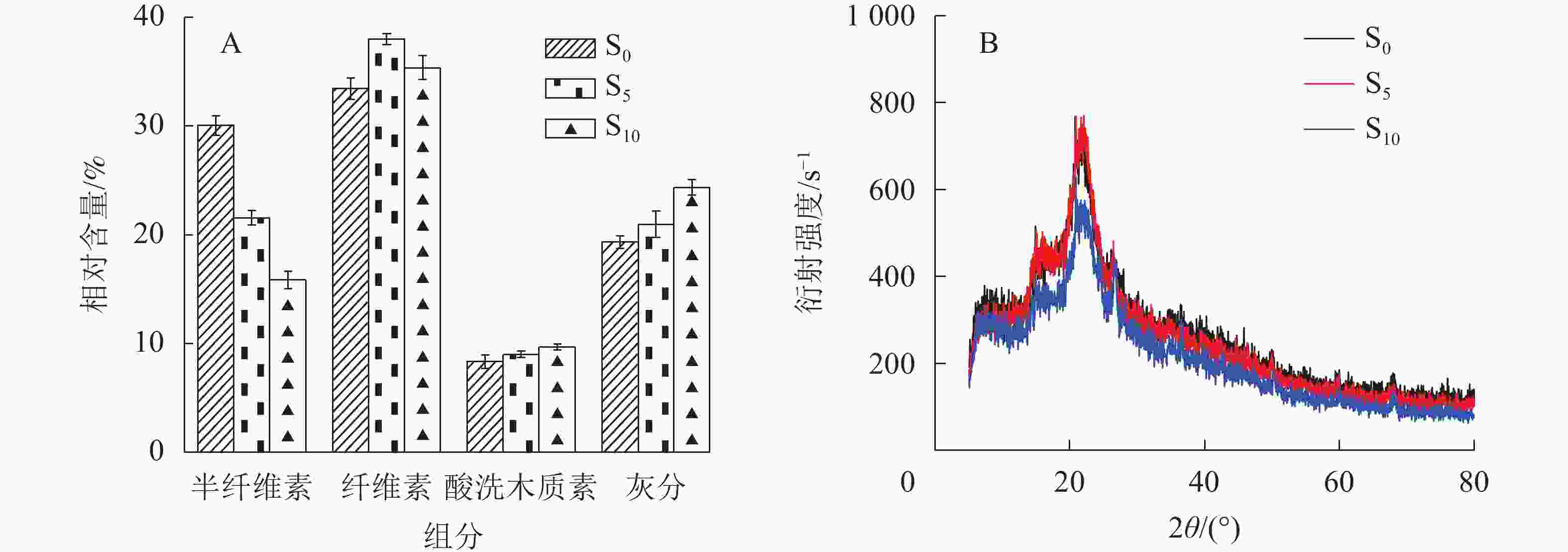
图 1 生物预处理秸秆组分相对含量变化(A)及X射线衍射图谱(B)
Figure 1. Relative content changes of the components (A) and X-ray diffraction pattern (B) of biologically pretreated straw fiber
-
张晖[19]研究发现:随着生物改性处理时间的延长秸秆逐渐由浅黄色变为深褐色,可能是木质素逐渐暴露出来所导致,本研究结果与此一致。未改性的秸秆纤维表面(图2A)存在硅和蜡质层,并含有淀粉等物质,这些结构紧凑的层状结构会影响脲醛树脂在秸秆表面的浸入及渗透,可能导致秸秆纤维/脲醛树脂复合材料力学强度的降低[10]。生物改性处理5 d后,S5表面的淀粉被消耗,硅等无法被微生物消耗的物质逐渐脱落,同时蜡质层被破坏,只在秸秆纤维表面零散分布(图2B)。硅质和蜡质的减少可提高脲醛树脂与秸秆纤维的相容性,形成良好的胶合界面,在一定程度上提高复合材料的力学性能[20]。随着生物改性处理时间的延长,微生物由秸秆纤维表面进入细胞腔内部,消耗秸秆组分,破坏纤维结构,使纤维表面形成裂隙(图2C)。裂隙的形成会造成秸秆纤维自身强度下降,但另一方面,由于这些裂隙的存在,脲醛树脂更容易进入秸秆纤维内部,在胶合过程中可能形成 “胶钉”,提高复合材料强度[21-22]。
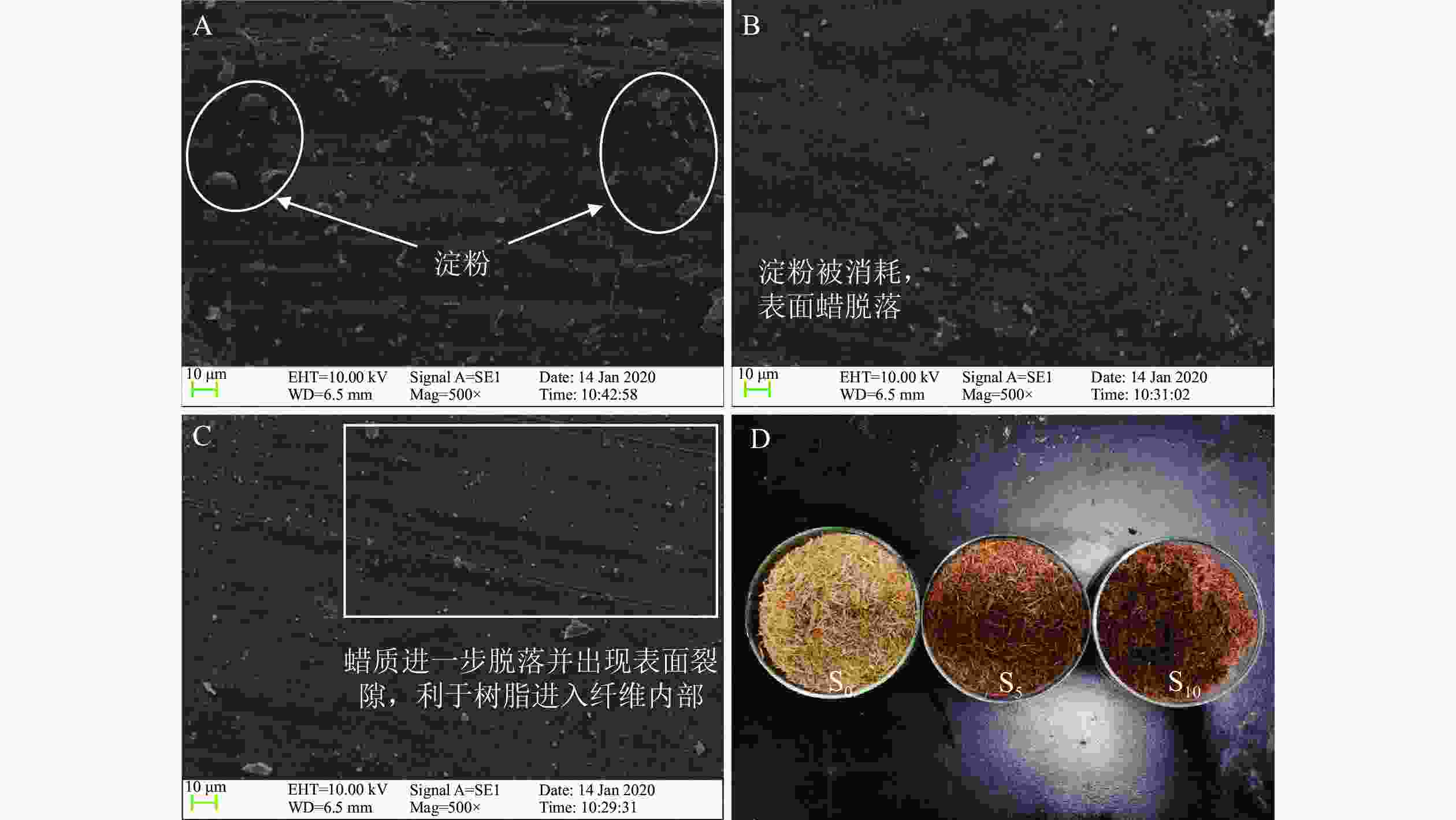
图 2 不同秸秆纤维S0(A)、S5(B)和S10(C)的表面微观形貌及其宏观对比(D)
Figure 2. Surface morphology of biologically pretreated straw fiber S0 (A), S5 (B) and S10 (C) and their macroscopic comparison (D)
-
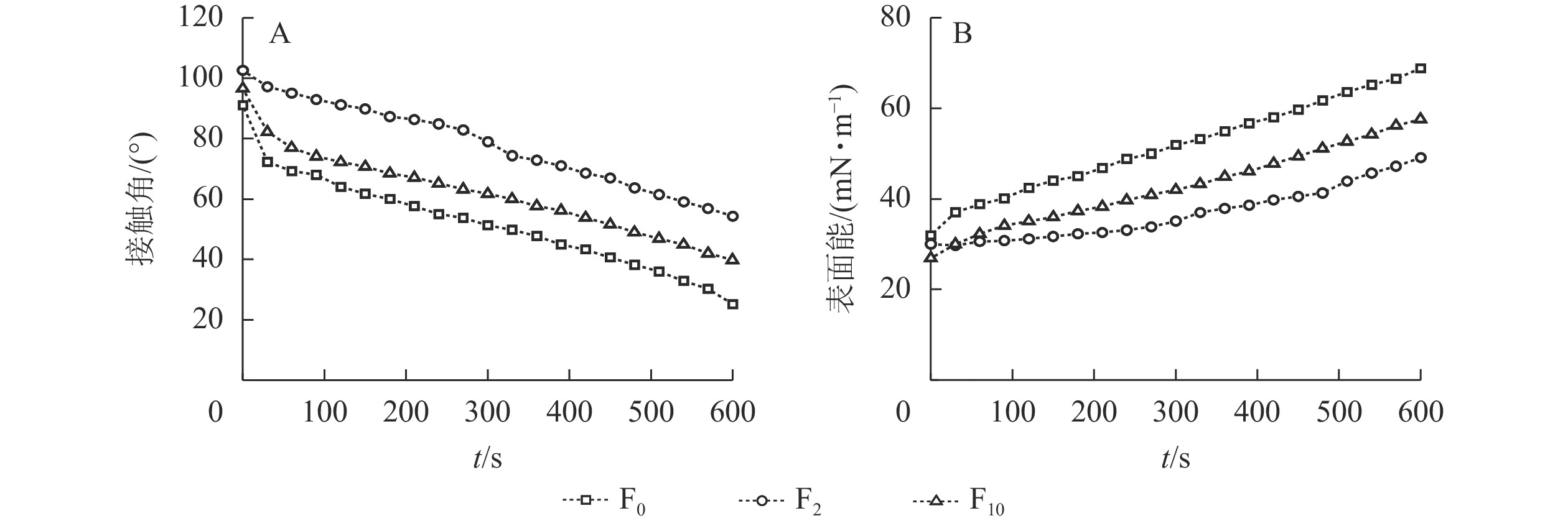
水滴在材料表面的接触角是表征材料表面润湿性直观而重要的手段,材料表面接触角越大表明材料疏水性越好,动态接触角则是量化润湿动力学的主要参数[23]。由图3A可知:与F0相比,生物改性处理秸秆纤维制备的复合材料接触角明显变大,在相同的测试时间内,F5的接触角更大,材料的疏水性更好。经过生物改性处理后秸秆纤维表面的蜡质等物质被去除,理论上秸秆纤维的润湿性更趋向于增强,但经过改性后,脲醛树脂更易附着在纤维表面或渗透进纤维内部,填充了纤维间隙,使得复合材料的疏水性增加。经过10 d的生物改性处理,S10表面出现裂隙,在复合材料制备过程中改性脲醛树脂易渗透到秸秆纤维内部,经热压固化后秸秆纤维表面的树脂固体含量低于S5表面, 因而F10的疏水性降低。
表面能是固体表面特征和表面现象形成的主要推动力,是描述和决定固体表面性质的重要物理量[24]。由图3B可知:未改性处理秸秆纤维中的半纤维素及纤维素含有大量的极性基团,F0的表面能因而更高;生物改性处理后秸秆中的半纤维素以及纤维素相对含量降低,极性基团减少,复合材料的表面能降低。另一方面,生物改性亦改变了秸秆纤维原有的表层结构,暴露出更多的木质素,这些木质素当中的羟基等极性基团有助于提高材料的表面能,导致F10的表面能高于F5[25],较低的表面能能够增加材料的抗污及抗氧化性能[26],有利于延长材料的使用寿命。
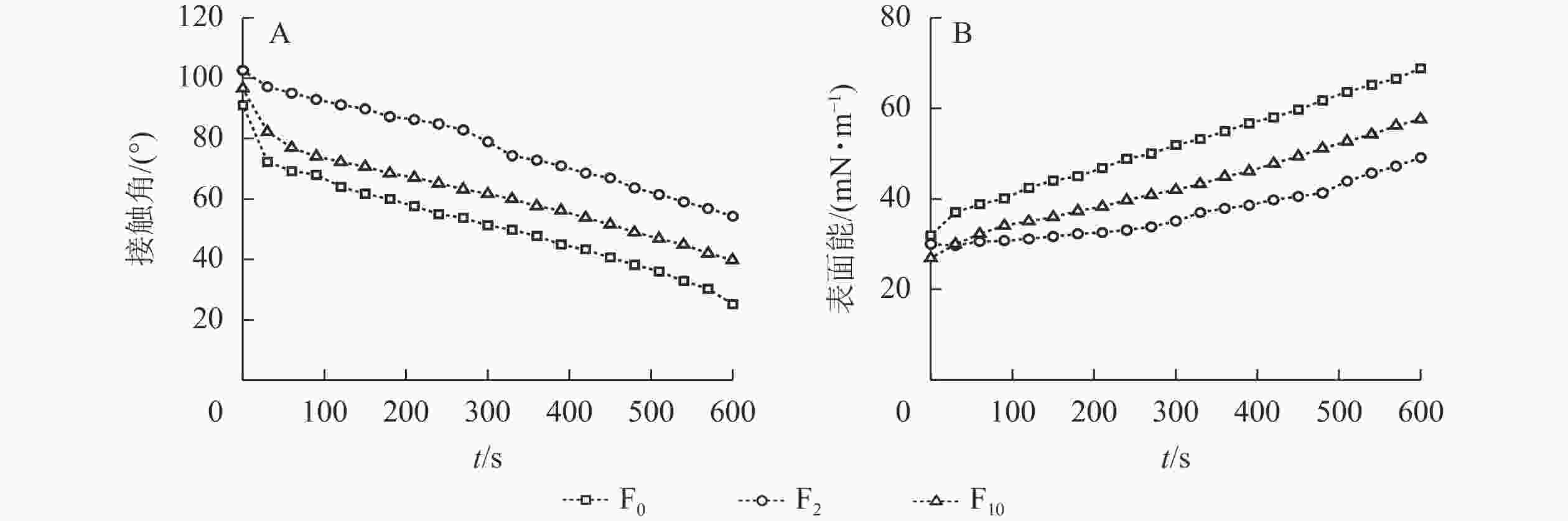
图 3 复合材料的接触角(A)及表面能(B)
Figure 3. Contact angle (A) and surface energy (B) of composites
-
由图4A可知:F0、F5和F10的静曲强度和弹性模量依次增强,相比于F0,F5和 F10的静曲强度分别提高30.05%和59.00%,弹性模量分别提高12.24%和50.17%。生物改性处理可以去除秸秆纤维表面的硅和蜡等物质,从而改善脲醛树脂与秸秆纤维的相容性,提高复合材料的抗弯性能。同时,随着生物改性处理时间的延长秸秆纤维表面出现裂隙,脲醛树脂易进入秸秆纤维内部,在增强秸秆纤维的同时,制备的复合材料的胶合界面形成 “胶钉”,提高复合材料的强度。
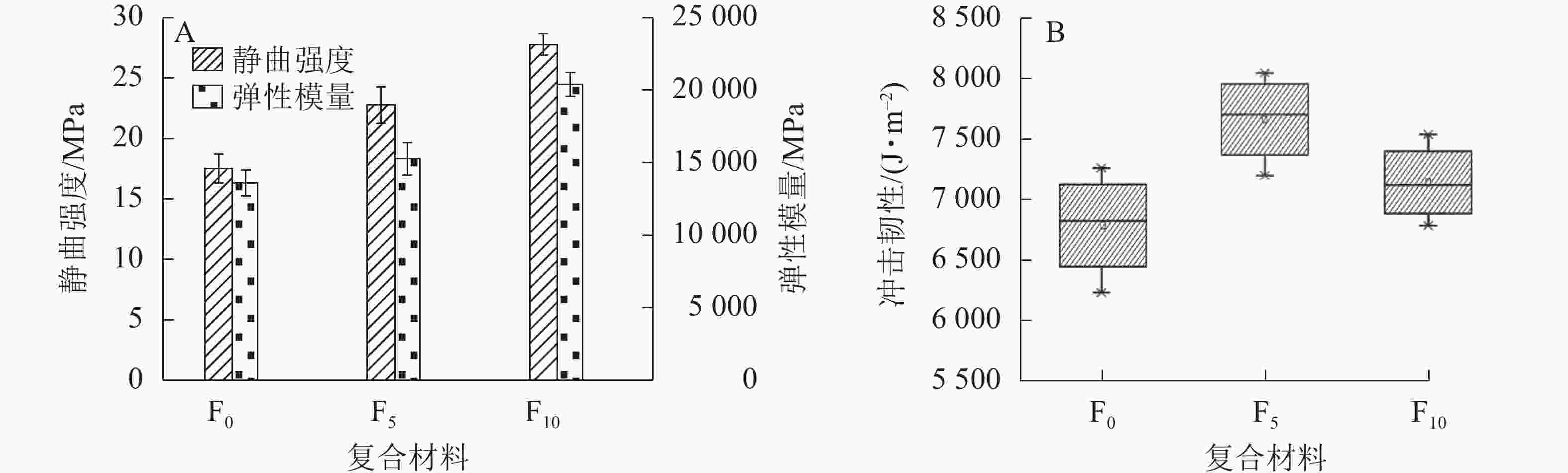
图 4 复合材料的抗弯强度(A)及冲击韧性(B)
Figure 4. Flexural strength (A) and impact toughness (B) of composites
如图4B所示:在不同生物改性处理时长的秸秆纤维制备的复合材料中, F5的冲击韧性更好,平均值为7 665.64 J·m−2,F10为7 143.26 J·m−2,而F0仅为6 786.40 J·m−2。复合材料的冲击韧性受到原材料自身力学性能、材料之间的相容性等多方面因素的影响[27-28],生物改性处理提高了秸秆纤维与脲醛树脂之间的相容性,脲醛树脂在秸秆纤维表面更加容易浸润和渗透,从而形成较为优异的胶合界面,提高复合材料的冲击韧性。随着生物改性处理时间的延长,秸秆纤维结构被微生物破坏,秸秆自身的冲击力学性能下降导致复合材料冲击韧性降低,F10的冲击韧性低于F5。
-
在特定微生物菌剂作用下,秸秆纤维的半纤维素最先降解,随后纤维素降解,而木质素和灰分则为不易降解物质。随生物改性处理时间延长,秸秆纤维的半纤维素相对含量逐渐降低,木质素和灰分的相对含量逐渐提高。生物改性前期由于半纤维素及纤维素不定形区被降解,纤维素定形区相对含量提高,因此S5的纤维素相对含量更高,结晶度更好。生物改性处理可去除秸秆纤维表面的硅质和蜡质,一定程度上提高了脲醛树脂与秸秆纤维的表面相容性,但随着改性时间的延长,秸秆纤维表面出现裂隙(S10)。生物改性处理减少极性基团,因而复合材料的疏水性更好,表面能更低,材料的耐久性和抗污性更加优良。同时脲醛树脂与秸秆纤维的相容性得以改善,F5和F10的抗弯强度和冲击韧性相比于F0有大幅提高,但因为生物改性处理后秸秆纤维结构被破坏,纤维自身的力学性能下降导致F10的冲击韧性低于F5。综合分析秸秆纤维的结晶度、微观形貌以及复合材料的表面性能和力学性能等,生物改性处理5 d为制备秸秆/脲醛树脂复合材料的适宜改性时长。
Properties of bio-pretreated straw fiber and its composite materials
-
摘要:
目的 探究生物预处理对秸秆纤维及其与脲醛树脂制备的复合材料性能的影响,为秸秆基复合材料的制备及发展提供理论依据。 方法 接种微生物菌剂(秸秆腐熟剂)对水稻Oryza sativa秸秆进行好氧发酵处理,测定不同处理时间下水稻秸秆中半纤维素、纤维素、木质素等的变化,测试并对比未经生物改性处理秸秆纤维(S0)、经生物改性处理5 (S5)和10 d (S10)秸秆纤维的结晶度和微观形貌,制备秸秆纤维/脲醛树脂复合材料,分别标记为F0、F5、F10,比较不同生物预处理时间下秸秆基复合材料的表面性能和力学性能。 结果 改性处理后秸秆表面的硅和蜡等物质被去除,但较长的生物改性处理时间(10 d)会破坏秸秆纤维自身结构。相比于S0和S10,S5的纤维素相对含量最高,为37.99%,结晶度也最好,为47.8%。3种秸秆基复合材料中F5疏水性最好,表面能最低,冲击韧性最大(7 665.64 J·m−2);F10抗弯性能更好,静曲强度和弹性模量分别为27.73和20 354 MPa,相比F0分别提高了59.00%和50.17%。 结论 生物改性处理可以改善秸秆纤维的表面性质,提高秸秆纤维/脲醛树脂复合材料的性能,生物改性处理5 d的秸秆纤维更好,制备的复合材料性能更优良。图4表1参28 Abstract:Objective This study aims to explore the influence of biological pretreatment on the properties of straw fiber and its composites prepared with urea formaldehyde resin, so as to provide theoretical basis for the preparation and development of straw based composites. Method Rice (Oryza sativa) straw was treated with microbial agent for aerobic fermentation. The changes of hemicellulose, cellulose and lignin in rice straw under different treatment time were measured. The crystallinity and microscopic morphology of straw fiber without biological pretreatment (S0), straw fiber bio-pretreated for 5 days (S5) and 10 days (S10) were tested and compared. Straw fiber/urea formaldehyde resin composites (F0, F5, F10) were prepared. Then the surface properties and mechanical properties of straw based composites under different biological pretreatment time were compared. Result Substances such as silicon and wax on the surface of straw fiber were removed after biological pretreatment, but the longer biological pretreatment time (10 d) could destroy the structure of straw fiber itself. Compared with S0 and S10, S5 had the highest relative content of cellulose (37.99%) and best crystallinity (47.8%). In contrast, F5 had the best hydrophobicity, lowest surface energy, and highest impact toughness (7 665.64 J·m−2). F10 had the best flexural performance. The static flexural strength and flexural modulus were 27.73 and 20 354 MPa, respectively, which were 59.00% and 50.17% higher than the composites prepared by S0, respectively. Conclusion Biological pretreatment can improve the surface properties of straw fiber and the properties of straw fiber/urea formaldehyde resin composites. The straw fiber bio-pretreated for 5 days is better, and the properties of the composites are superior. [Ch, 4 fig. 1 tab. 28 ref.] -
Key words:
- straw fiber /
- biological pretreatment /
- composites /
- surface properties /
- mechanical properties
-
图 1 生物预处理秸秆组分相对含量变化(A)及X射线衍射图谱(B)
Figure 1 Relative content changes of the components (A) and X-ray diffraction pattern (B) of biologically pretreated straw fiber
图 2 不同秸秆纤维S0(A)、S5(B)和S10(C)的表面微观形貌及其宏观对比(D)
Figure 2 Surface morphology of biologically pretreated straw fiber S0 (A), S5 (B) and S10 (C) and their macroscopic comparison (D)
图 3 复合材料的接触角(A)及表面能(B)
Figure 3 Contact angle (A) and surface energy (B) of composites
图 4 复合材料的抗弯强度(A)及冲击韧性(B)
Figure 4 Flexural strength (A) and impact toughness (B) of composites
表 1 脲醛树脂制备过程中物料添加情况
Table 1. Adding quality of materials during the preparation of urea-formaldehyde resin
阶段 37%(质量分数)
甲醛溶液/g尿素/g 聚乙烯
醇-124/g三聚氰
胺/gPEGDGE/g 1 1 000.0 371.0 5.3 2 106.0 10.6 3 53.0 106.0  下载: 导出CSV
下载: 导出CSV
-
[1] 王红梅, 屠焰, 张乃锋, 等. 中国农作物秸秆资源量及其“五料化”利用现状[J]. 科技导报, 2017, 35(21): 81 − 88. WANG Hongmei, TU Yan, ZHANG Naifeng, et al. Chinese crop straw resource and its utilization status [J]. Sci Technol Rev, 2017, 35(21): 81 − 88. [2] 霍丽丽, 赵立欣, 孟海波, 等. 中国农作物秸秆综合利用潜力研究[J]. 农业工程学报, 2019, 35(13): 218 − 224. HUO Lili, ZHAO Lixin, MENG Haibo, et al. Study on straw multi-use potential in China [J]. Trans Chin Soc Agric Eng, 2019, 35(13): 218 − 224. [3] DONG Chensong. Review of natural fibre-reinforced hybrid composites [J]. J Reinf Plast Compos, 2018, 37(5): 331 − 348. [4] 杨雪慧, 汤丽娟, 章蓉, 等. 农作物秸秆表面改性处理的研究进展[J]. 南京林业大学学报(自然科学版), 2013, 37(3): 157 − 162. YANG Xuehui, TANG Lijuan, ZHANG Rong, et al. Review on progress of crop straws surface modification [J]. J Nanjing For Univ Nat Sci Ed, 2013, 37(3): 157 − 162. [5] BARAMEE S, SIRIATCHARANON A K, KETBOT P, et al. Biological pretreatment of rice straw with cellulase-free xylanolytic enzyme-producing Bacillus firmus K-1: structural modification and biomass digestibility [J]. Renew Energy, 2020, 160: 555 − 563. [6] MA Yingqun, SHEN Yanqing, LIU Yu. 秸秆处理技术的最新进展: 挑战与解决方案[J]. 中国农机化学报, 2020, 41(11): 152 − 161. MA Yingqun, SHEN Yanqing, LIU Yu. State of the art of straw treatment technology: challenges and solutions forward [J]. J Chin Agric Mech, 2020, 41(11): 152 − 161. [7] TIAN Jianghao, POURCHER A M, BIZE A, et al. Impact of wet aerobic pretreatments on cellulose accessibility and bacterial communities in rape straw [J]. Bioresour Technol, 2017, 237: 31 − 38. [8] QUÉMÉNEUR M, BITTEL M, TRABLY E, et al. Effect of enzyme addition on fermentative hydrogen production from wheat straw [J]. Int J Hydrog Energy, 2012, 37(14): 10639 − 10647. [9] QU Ping, HUANG Hongying, ZHAO Yongfu, et al. Physicochemical changes in rice straw after composting and its effect on rice-straw-based composites [J/OL]. J Appl Polym Sci, 2017, 134(22): 44878[2021-09-20]. doi: 10.1002/APP. 44878. [10] LI Jia, ZHANG Xiaolin, NIE Sunjian, et al. Effects of surface treatment on the properties of wheat straw fiber-reinforced rHDPE composites [J/OL]. Mater Res Express, 2019, 6(12): 125103[2021-09-20]. doi: 10.1008/2053-1591/ab4636. [11] XU Chao, LI Jun, YUAN Qiaoxia, et al. Effects of different fermentation assisted enzyme treatments on the composition, microstructure and physicochemical properties of wheat straw used as a substitute for peat in nursery substrates [J/OL]. Bioresour Technol, 2021, 341: 125815[2021-09-20]. doi: 10.1016/j.biortech.2021.125815. [12] GHAFFAR S H, FAN Mizi, MCVICAR B. Bioengineering for utilisation and bioconversion of straw biomass into bio-products [J]. Ind Crops Prod, 2015, 77: 262 − 274. [13] MA Yingqun, SHEN Yanqing, LIU Yu. State of the art of straw treatment technology: challenges and solutions forward [J/OL]. Bioresour Technol, 2020, 313: 123656[2021-09-20]. doi: 10.1016/j.biortech.2020.123656. [14] 王士强, 顾春梅, 赵海红. 木质纤维素生物降解机理及其降解菌筛选方法研究进展[J]. 华北农学报, 2010, 25(增刊 1): 313 − 317. WANG Shiqiang, GU Chunmei, ZHAO Haihong. The research progress on the mechanisms of lignocellulose biological degrading and the screening method on the degrading bacteria [J]. Acta Agric Boreali-Sin, 2010, 25(suppl 1): 313 − 317. [15] 李思蓓, 解玉红, 罗晶, 等. 秸秆预处理中木质纤维物质含量测定方法的研究进展[J]. 安徽农业科学, 2011, 39(3): 1620 − 1622, 1626. LI Sibei, XIE Yuhong, LUO Jing, et al. Research progress of content determination method of xylem fiber material in straw pretreatment [J]. J Anhui Agric Sci, 2011, 39(3): 1620 − 1622, 1626. [16] 刘永明, 施建宇, 鹿芹芹, 等. 基于杨氏方程的固体表面能计算研究进展[J]. 材料导报, 2013, 27(11): 123 − 129. LIU Yongming, SHI Jianyu, LU Qinqin, et al. Research progress on calculation of solid surface tension based on Young’s equation [J]. Mater Rev, 2013, 27(11): 123 − 129. [17] MATHEWS S L, PAWLAK J, GRUNDEN A M. Bacterial biodegradation and bioconversion of industrial lignocellulosic streams [J]. Appl Microbiol Biotechnol, 2015, 99(7): 2939 − 2954. [18] 付春霞, 付云霞, 邱忠平, 等. 木质素生物降解的研究进展[J]. 浙江农业学报, 2014, 26(4): 1139 − 1144. FU Chunxia, FU Yunxia, QIU Zhongping, et al. Research progresses of lignin biodegradation [J]. Acta Agric Zhejiang, 2014, 26(4): 1139 − 1144. [19] 张晖. 木质素的降色及其在木质素基化学品中的应用[D]. 广州: 华南理工大学, 2018. ZHANG Hui. Color Reduction of Ligin and Its Applications in the Ligin based Chemicals [D]. Guangzhou: South China University of Technology, 2018. [20] BOQUILLON N, ELBEZ G R, SCHÖNFELD U. Properties of wheat straw particleboards bonded with different types of resin [J]. J Wood Sci, 2004, 50(3): 230 − 235. [21] GUAN Mingjie, YONG Cheng, WANG Lu. Shear strain and microscopic characterization of a bamboo bonding interface with poly (vinyl alcohol) modified phenol-formaldehyde resin [J]. J Appl Polym Sci, 2013, 130(2): 1345 − 1350. [22] GUAN Mingjie, HUANG Zhiwei, ZENG Dan. Shear strength and microscopic characterization of a bamboo bonding interface with phenol formaldehyde resins modified with larch thanaka and urea [J]. Bioresources, 2016, 11(1): 492 − 502. [23] ZHAO Hongping, ZHU Jintang, FU Zhongyu, et al. Plasma surface graft of acrylic acid and biodegradation of poly (butylene succinate) films [J]. Thin Solid Films, 2008, 516(16): 5659 − 5663. [24] WANG Jun, BIAN Jianjun, WANG Gangfeng. Calculation of surface energy density of rough surface by atomic simulations [J]. Appl Surf Sci, 2019, 484: 184 − 188. [25] 王春红, 刘胜凯. 碱处理对竹纤维及竹纤维增强聚丙烯复合材料性能的影响[J]. 复合材料学报, 2015, 32(3): 683 − 690. WANG Chunhong, LIU Shengkai. Effects of alkali treatment on properties of bamboo fiber and bamboo fiber reinforced polypropylene composite [J]. Acta Mater Compos Sin, 2015, 32(3): 683 − 690. [26] 赵晓非, 杨明全, 章磊, 等. 仿生超疏水表面的制备与应用的研究进展[J]. 化工进展, 2016, 35(9): 2818 − 2829. ZHAO Xiaofei, YANG Mingquan, ZHANG Lei, et al. Research progress in fabrication and application of bioinspired super-hydrophobic surface [J]. Chem Ind End Prog, 2016, 35(9): 2818 − 2829. [27] SWOLFS Y, GEBOES Y, GORBATIKH L, et al. The importance of translaminar fracture toughness for the penetration impact behaviour of woven carbon/glass hybrid composites [J]. Compos Part A Appl Sci Manuf, 2017, 103: 1 − 8. [28] AL-MAHARMA A Y, SENDUR P. Review of the main factors controlling the fracture toughness and impact strength properties of natural composites [J/OL]. Mater Res Express, 2019, 6(2): 022001[2021-09-20]. doi: 10.1088/2053-1591/aaaec28. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210647
 点击查看大图
点击查看大图
计量
- 文章访问数: 1977
- HTML全文浏览量: 1272
- PDF下载量: 60
- 被引次数: 0
