





-
20世纪50年代以来,土壤有机碳含量和储量一直是研究焦点之一[1]。1982年,POST等[2]通过建立全球土壤碳密度的地理分布与气候因子和植被因子间的关系,估算全球1 m厚度土壤有机碳库存量约1 395 Pg。此后,国内外学者采取不同估算方法,推算出全球1 m土层中土壤有机碳库为1 143~1 576 Gt[3-5];另有国内学者依据2次全国土壤普查所得的土壤属性数据,估算中国有机碳库为50.0~80.7 Gt[6]。为了掌握土壤有机碳累积变化特点,更多研究者在小尺度范围探讨土壤有机碳的相关影响因子,发现植被类型[7-8]、气候条件[9]、林龄[10-11]、造林[12]和抚育间伐措施[13-14]以及土壤理化性质[15]等都可能影响土壤有机碳累积。油松Pinus tabulaeformis是中国北方主要造林树种之一,分布面积约161×104 hm2,其土壤有机碳储量及动态是影响土壤碳循环的重要因素。油松林地主要土壤类型为山地棕壤和褐土,其土层相对较薄[10],土壤有机碳的相关研究大多集中于60 cm以上土层,而其下土层数据多为估算值,实测数据极少。1996年,马钦彦等[16]以中国山西太岳山和北京西山油松天然纯林为研究对象,估算中国油松林0~130 cm土层深度的平均土壤有机碳含量为90 t·hm−2,但存在点位过少,研究尺度小等问题。此后相关研究多以小区域为基础,探讨不同因子对油松林土壤有机碳的影响[7, 10-11],而大尺度范围的相关研究鲜有报道。因此,从宏观角度量化油松林土壤有机碳变化及其影响因素,对今后探讨其在调节碳平衡中的作用和贡献具有重要意义。本研究通过收集1980−2017年油松林土壤有机碳和相关影响因子文献中实测数据,分析0~60 cm土层土壤有机碳质量分数和储量的垂直变化和时间变化特征及主要影响因子,旨在为今后估算及预测油松林有机碳质量分数和储量变化提供参考,也为揭示油松林土壤对碳循环的贡献提供理论支持。
-
利用Web of Science和中国知网、万方数据库等,分别以主题词“Pinus tabulaeformis & Soil organic carbon”和“油松&土壤有机碳”检索1980−2017年CSCD-E库及以上公开发表学术期刊,并设置筛选标准如下:①数据基于实测值;②土壤类型为棕壤和褐土。文献采集数据包括各层土壤有机碳质量分数、土壤理化性质(土壤容重、全氮、碳氮比和pH)、林木特征(凋落物、林分密度、郁闭度和林龄)以及气候因子(年均气温和年均降水量)共计10个因子的相关数据,当研究点没有年均气温和年均降水量信息时,根据研究点的经纬度及研究时间,通过使用中国气象数据网网站进行补充。最后共收集165篇有效文献。
-
采集的数据首先按照1980−1989、1990−1999、2000−2009和2010−2017年4个时段以及0~20、20~40和40~60 cm的3个土层深度分类汇总(图1),其后按照下述方法对数据进行整理和补充。
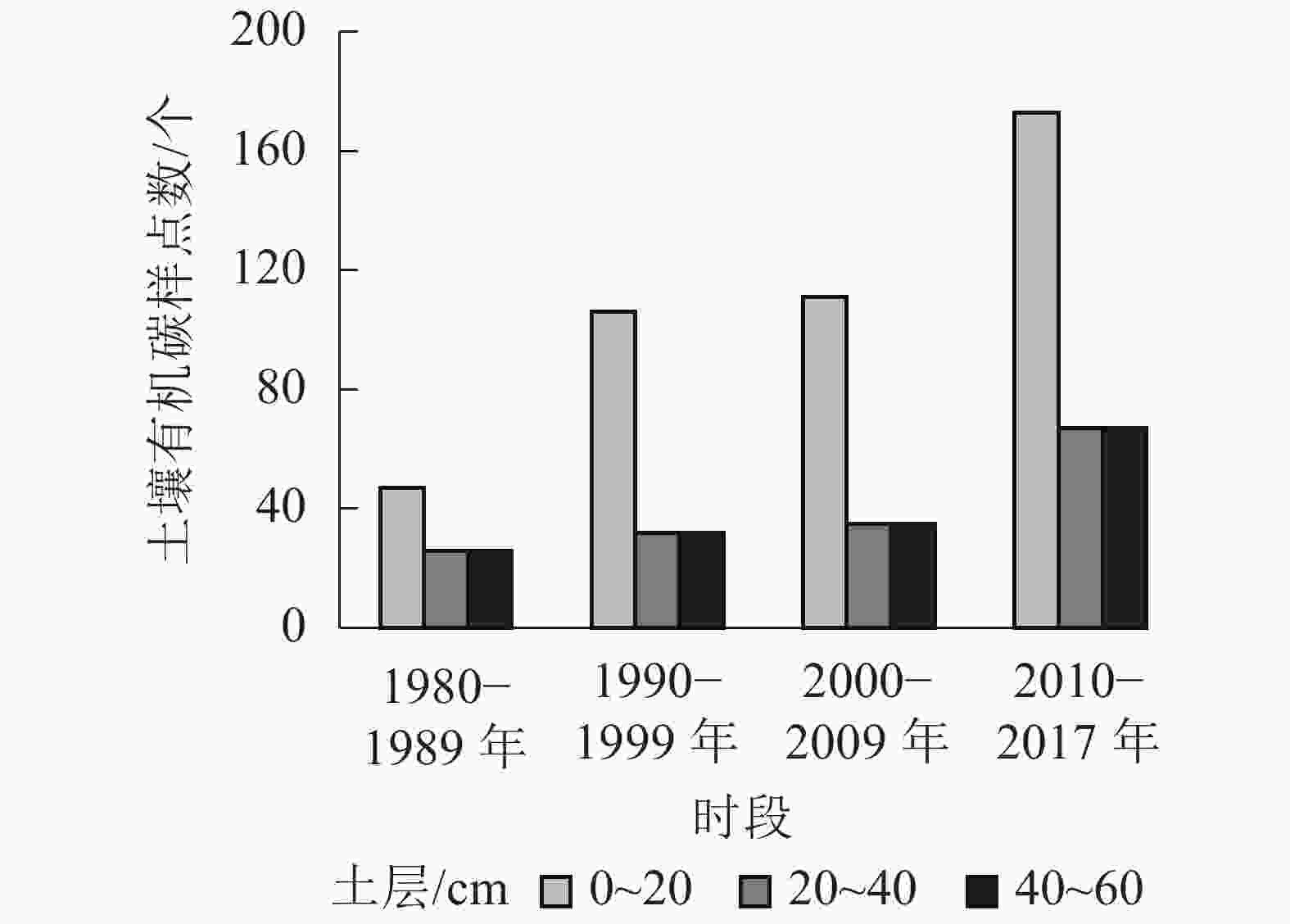
图 1 土壤有机碳样点数量
Figure 1. Quantitative distribution of soil organic carbon
-
由于部分文献土层深度划分标准不符合本研究划分要求,为此利用加权平均法对土层深度和对应的土壤性质数据进行等间隔深度转换[17]。
-
将文献中土壤全氮和土壤有机碳质量分数单位统一转化为g·kg−1,并根据式(1)将部分文献中土壤有机质质量分数转化为土壤有机碳质量分数:
$$ {C_{{\rm{SOC}}}} = {C_{{\rm{SOM}}}}/1.724{\text{。}} $$ (1) 式(1)中:CSOC为土壤有机碳质量分数(g·kg−1);CSOM为土壤有机质质量分数(g·kg−1)。
-
对于部分文献中缺失的土壤容重数据采用下述方法进行补充。筛选同时具有土壤有机质和土壤容重的文献,将各点的土壤有机质值代入式(2)[18]获取该点的土壤容重估算值:
$$ {B_{\rm{D}}} = \dfrac{{100}}{{\dfrac{{{C_{{\rm{SOM}}}}}}{{0.244}} + \dfrac{{100 - {C_{{\rm{SOM}}}}}}{{1.64}}}}{\text{。}} $$ (2) 式(2)中:BD为土壤容重(g·cm−3);CSOM为土壤有机质质量分数(g·kg−1)。
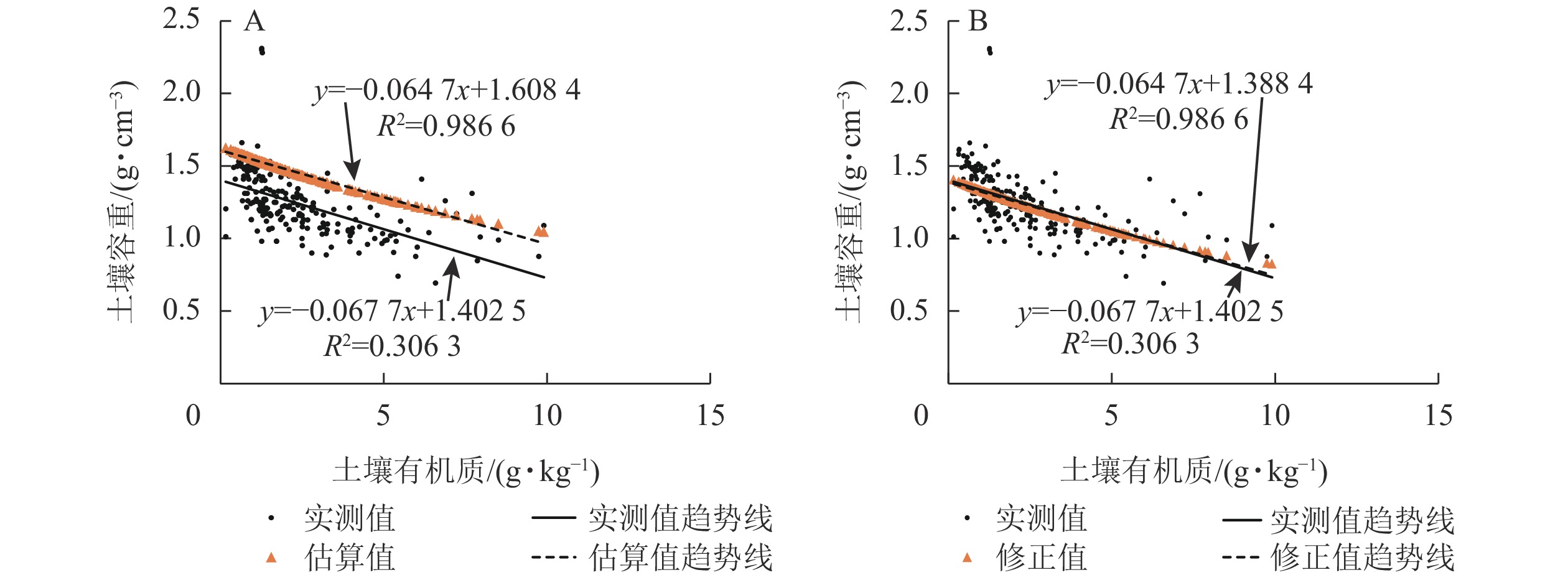
以各点的实测值和估算值为成对变量进行t检验。结果表明各点的估算值比实测值平均高0.22 g·cm−3(图2A),存在明显差异。据此,对式(2)修正后获得:

图 2 土壤容重实测值与估算值比较
Figure 2. Comparison of measured and calculated soil bulk density
$$ {B_{\rm{D}}} = \dfrac{{100}}{{\dfrac{{{C_{{\rm{SOM}}}}}}{{0.244}} + \dfrac{{100 - {C_{{\rm{SOM}}}}}}{{1.64}}}} - 0.22{\text{。}} $$ (3) 再次利用t检验对估算值和实测值进行显著性分析。结果表明:式(3)获得的土壤容重估算值和实测值之间差异不显著,可以更好地反映油松林土壤有机质质量分数和土壤容重间的关系(图2B)。为此,利用修正后的式(3)对缺失的容重数据进行补充。
-
油松林各层土壤有机碳储量计算公式[19]为:
$$ {C_{{\rm{SOCs}}}} = \sum\limits_{{{i}} = 1}^{{n}} \left[{{C_{{\rm{SOC}}i}}} \times {B_{{\rm{D}}i}} \times {T_i} \times \left( {1 - {G_i}} \right)\right] \times S \times {10^{ - 1}}{\text{。}} $$ (4) 式(4)中:CSOCs为某特定深度土壤有机碳储量(Tg);CSOCi为第i层土壤有机碳质量分数(g·kg−1);BDi为第i层土壤容重(g·cm−3);Ti为第i层土层厚度(cm);Gi为直径大于2 mm的石砾所占的体积百分比(%);n为参与计算的土壤层次总数;S为4个时段油松林分布总面积(hm2)。油松林不同时段各林龄面积及占比见表1。
表 1 不同时段油松林各龄组面积及占比
Table 1. Statistics of P. tabulaeformis forest area at different stages
龄组 1980−1989年 1990−1999年 2000−2009年 2010−2017年 面积/(×104 hm2) 占比/% 面积/(×104 hm2) 占比/% 面积/(×104 hm2) 占比/% 面积/(×104 hm2) 占比/% 幼龄林 132.9 64 102.2 42 74.6 32 55.0 22 中龄林 59.4 29 91.4 38 103.9 45 103.1 41 近熟林 13.8 7 49.5 20 54.7 23 93.1 37 总计 206.1 100 243.1 100 233.2 100 251.2 100 说明:林龄划分按照《中国森林(第1卷)》[20]标准,其中,近熟林为划分标准中的近熟林及以上林龄的油松林;不同时期幼龄林、中 龄林和近熟林的面积根据文献[21-27]统计数据计算获取 -
采集数据经Excel 2016初步整理后,利用SPSS 23.0进行单因素方差分析(one-way ANOVA),检验相同土层不同时段以及相同时段不同土层土壤有机碳质量分数差异,多重比较采用最小显著差异(LSD)法,相关性分析采用Pearson检验,并利用多元逐步回归和通径分析相结合的方法分析土壤性质和林分特征对土壤有机碳质量分数的影响。
-
从表2可知:各时段土壤有机碳质量分数表聚现象明显,0~20 cm土层土壤有机碳质量分数占整个剖面的47%~54%,表现出随土层深度的增加而减少的剖面垂直变化特征。各层土壤有机碳质量分数表现出不同程度的时间变化特点,其中0~20 cm土层土壤变化最为强烈,在1980−1989年为19.50 g·kg−1,此后经历了20 a的持续降低,2000−2009年达到最低点,仅14.57 g·kg−1,2010−2017年又显著增加至20.76 g·kg−1。20~60 cm土壤有机碳质量分数时间变化趋势与0~20 cm相同,同样表现出高—低—高的变化特点,但各个时段间不存在层间显著差异,土壤碳质量分数相对稳定。各层土壤有机碳质量分数变异系数为0.55~0.96,属于中等变异。
表 2 不同时段土壤有机碳质量分数统计分析
Table 2. Variation of soil organic carbon content at different stages
时段 0~20 cm有机碳/(g·kg−1) 20~40 cm有机碳/(g·kg−1) 40~60 cm有机碳/(g·kg−1) 0~60 cm有机碳平
均值/(g·kg−1)最大值 最小值 平均值 变异系数 最大值 最小值 平均值 变异系数 最大值 最小值 平均值 变异系数 1980−1989年 45.00 4.63 19.50 Aa 0.57 27.50 4.26 11.48 Ab 0.56 30.62 2.70 8.37 Abc 0.79 13.11 A 1990−1999年 41.93 4.11 15.00 Ba 0.56 44.66 1.34 10.09 Ab 0.86 22.33 0.68 6.15 Ac 0.67 10.41 B 2000−2009年 56.55 1.20 14.57 Ba 0.66 45.62 1.00 8.49 Ab 0.96 34.68 0.68 6.33 Abc 0.96 9.80 B 2010−2017年 57.45 0.50 20.76 Aa 0.55 35.28 0.74 11.36 Ab 0.62 30.20 0.65 7.94 Ac 0.68 13.35 A 说明:不同大写字母表示相同土层不同时段有机碳质量分数差异显著(P<0.05);不同小写字母表示相同时段不同土层土壤有机碳质 量分数差异显著(P<0.05) -
由表3可以看出:0~20 cm土层是油松林土壤有机碳的主要碳库,1980−1989、1990−1999、2000−2009和2010−2017年4个时期的碳储量分别为97.18、90.20、86.27和124.92 Tg,占0~60 cm土层土壤总碳储量的45.41%~50.57%。随着土层加深,有机碳储量降低,20~40和40~60 cm土层土壤有机碳储量占比分别为28.58%~33.76%和20.83%~22.78%。在1980−2017年,各土层土壤有机碳储量表现出与土壤有机碳质量分数一致的时间变化特征。与1980−1989年相比,1990−1999年土壤有机碳储量呈降低趋势,但总体变化不大,0~60 cm土层土壤有机碳储量减少了6.81 Tg;进入2000−2009年后,有机碳储量降低幅度较大,共减少14.81 Tg;在2010−2017年,土壤有机碳储量快速增加,达247.02 Tg,为40 a来的最高水平,特别是0~20 cm土层土壤有机碳储量增幅达38.65 Tg,占增幅总量的61%。综上可见,近40 a间0~60 cm土层土壤有机碳库变动主要集中在0~20 cm土层,而20~60 cm土层土壤有机碳储量始终保持相对稳定状态。
表 3 不同时段土壤有机碳储量及占比
Table 3. Estimation of soil organic carbon storage in P. tabulaeformis forest at different stages
土层/cm 1980−1989年 1990−1999年 2000−2009年 2010−2017年 储量/Tg 占比/% 储量/Tg 占比/% 储量/Tg 占比/% 储量/Tg 占比/% 0~20 97.18 47.30 90.20 45.41 86.27 46.93 124.92 50.57 20~40 61.85 30.10 67.06 33.76 55.68 30.29 70.60 28.58 40~60 46.43 22.60 41.38 20.83 41.88 22.78 51.49 20.85 0~60 205.45 100.00 198.64 100.00 183.83 100.00 247.02 100.00 -
表4表明:土壤有机碳质量分数与土壤容重呈极显著负相关(P<0.01),与土壤全氮、pH、郁闭度以及林龄呈极显著正相关(P<0.01),而与年均气温、年均降水量和林分密度相关不显著。
表 4 0~20 cm土层土壤有机碳质量分数与各因子间的Pearson相关性分析
Table 4. Pearson correlation analysis between 0−20 cm soil organic carbon and each factors
因子 有机碳 容重 全氮 碳氮比 pH 年均气温 年均降水量 凋落物量 林分密度 郁闭度 林龄 有机碳 1 容重 −0.774** 1 全氮 0.627** 0.424** 1 碳氮比 0.609** −0.425** 0.446** 1 pH 0.140** −0.084 0.212* 0.209* 1 年均气温 −0.052 0.037 −0.016** 0.259** 0.023 1 年均降水量 0.158 −0.126 0.059 0.031 −0.300* 0.339** 1 凋落物量 0.042 0.131 −0.031 0.144 −0.113 0.273* 0.228 1 林分密度 0.055 −0.147* −0.094 −0.232* 0.017 0.161 0.149 0.169 1 郁闭度 0.336** −0.159 0.246 0.221 0.048 0.047 0.090 0.220 −0.128 1 林龄 0.376** −0.297* 0.469** 0.454** 0.021 0.088 0.064 0.211 −0.455** 0.269** 1 说明:*表示显著相关(P<0.05);**表示极显著相关(P<0.01) -
将土壤有机碳质量分数(y)与土壤容重(x1)、全氮(x2)、郁闭度(x3)进行逐步多元回归分析,得到标准多元回归方程:
$y=-0.061\;2 x_{1}+0.037\;6 x_{2}+0.310\;0x_{3}$ 。其中,回归方程中的系数为直接通径系数。 -
多元回归结果表明:土壤容重、全氮、郁闭度3个因子对0~20 cm土壤有机碳质量分数影响显著,但各个影响因子间的相关性导致信息重叠,仅根据土壤有机碳质量分数与影响因子间的系数,仍然无法明确各个因子对有机碳质量分数变化的影响程度。为此,将逐步多元线性回归分析中的变量均纳入通径分析,通径分析[28-29]是研究多个变量与因变量之间线性关系的多元统计方法,当多个解释变量交互作用对响应变量产生影响时,通径分析将极具优势。根据计算获得间接通径系数和决定系数,分析3个因子对土壤有机碳质量分数的影响作用。
表5及图3结果表明:土壤容重对有机碳表现出强烈的直接负效应,显著掩盖了间接正效应的影响,导致其降低土壤有机碳的作用较强。同理,全氮和郁闭度表现出增加土壤有机碳的作用。决定系数(R2)是通径分析中的综合性决策指标,按绝对值大小对有机碳的影响从大到小依次为容重(R2=0.375)、容重与全氮相互作用(R2=0.195)、全氮(R2=0.141)、郁闭度(R2=0.096),表明土壤容重、全氮和郁闭度以及三者间的相互作用共同控制着土壤有机碳的变化强度和方向。
表 5 通径分析结果
Table 5. Results of path analysis of factors
因子 直接通径系数 间接通径系数 决定系数 容重 全氮 郁闭度 合计 容重 全氮 郁闭度 容重 −0.612 −0.259 0.097 −0.162 0.375 0.195 0.060 全氮 0.376 0.159 0.092 0.251 0.141 0.057 郁闭度 0.310 −0.049 0.076 0.027 0.096 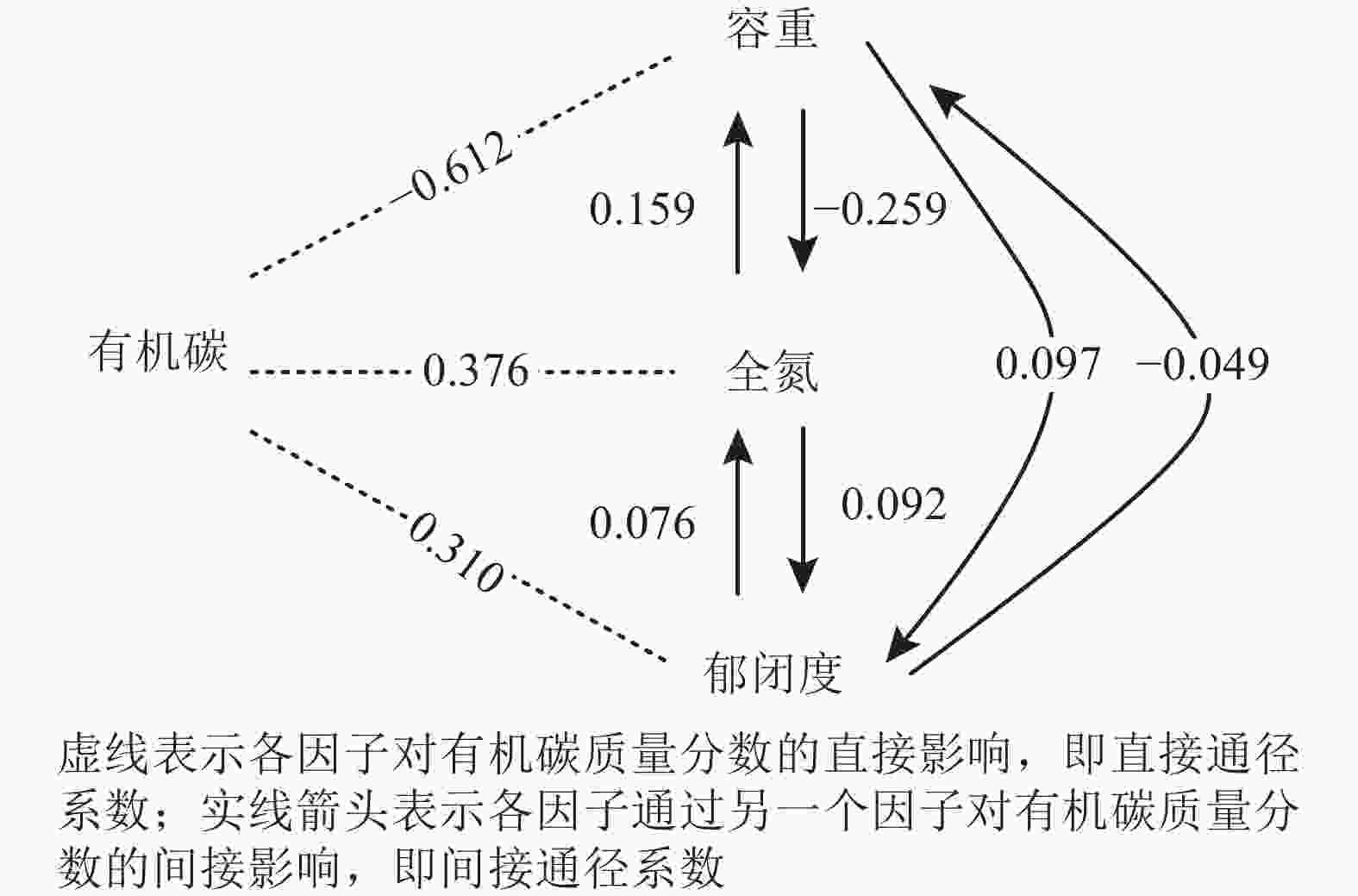
图 3 土壤有机碳质量分数对各因子的响应
Figure 3. Soil organic carbon content response to factors
-
土壤容重对土壤有机碳表现出明显的直接负效应,主要由于容重通过改变土壤孔隙结构特点影响土壤水气环境,改变土壤微生物的生长和活动特征,对土壤有机碳转化产生强烈影响[30];此外,逯军峰等[31]研究发现:油松林土壤容重与凋落物总量、凋落物现存量、分解率表现出显著负相关。土壤全氮和碳氮比对土壤有机碳的转化方向和强度有重要作用。王棣等[8]研究表明:容重与土壤全氮、碳氮比呈显著负相关,这与本研究结果一致。全氮对土壤有机碳呈显著正效应,氮素作为土壤微生物的重要营养元素,能够对土壤微生物活动产生直接影响。还有研究发现:较高的全氮能够降低凋落物叶中碳氮比,避免微生物与植物的“争氮”现象,利于凋落物矿化分解[32-33]。郁闭度是影响林下植被群落结构、改变凋落物组成和林地土壤特征的重要因素。油松人工林中,乔木层植物种类单一,结构简单,养分分解归还速率慢[34],林下植被作用更为重要[35]。低郁闭度条件下可以丰富林下植被群落,减少凋落物中的木质素和粗纤维等难溶性物质含量以及改善林地土壤性质,从而提高土壤微生物对油松凋落物的分解速度[36],促进土壤有机碳变化。
有研究指出:降水[37]、气温[38-39]、pH[39]和种植密度[40]是油松林土壤有机碳变化的可能影响因素,这与本研究结果不同。本研究表明:近40 a间研究区年均降水量、年均气温和土壤pH均未出现明显变化,年均降水量最大相差仅54.7 mm,年均气温维持在7.7~8.8 ℃,土壤均为中性(pH 6.99~7.58)。此外,中国油松林以人工林为主,不同地区对种植密度要求虽有不同,但整体差异不大[41]。由于年均气温、年均降水量和土壤pH的稳定性以及人工种植密度的相对一致性,使上述因素没有对油松分布区土壤有机碳产生强烈影响。
-
棕壤和褐土在形成过程中存在强烈的黏化过程,从而在淋溶层以下形成厚度40~80 cm的黏化层,表现出黏粒含量高(20%~30%)、土壤紧实致密、渗透性差的土层特征[42],使凋落物分解补充的有机碳源主要集中在表层土壤,土壤有机碳呈现出表聚特征,但其下层土壤有机碳始终相对稳定且保持在较低水平。近40 a间研究区年均降水量为560~610 mm,变幅较小,最大相差仅50 mm,各时期上层土壤中可溶性有机碳淋溶量相对稳定。由于黏化层对淋溶强度的制约以及每年死亡根系数量有限,导致下层土壤有机碳输入量远低于上层土壤,加之黏化层的影响造成土壤水气状况不良,微生物繁殖和活动被抑制,导致下层土壤有机质分解转换缓慢。
-
油松林土壤有机碳变化与林木生长特征密切相关,这已在诸多研究中得以证实[10-11,43]。由于油松造林地土壤相对肥沃,立地条件较好,育种初期幼龄林有机碳处于较高水平。造林前的整地、蓄水等营林措施改善土壤容重,营造良好水、气条件,土壤微生物繁殖和活动旺盛,利于有机质的分解转化[44],为幼林生长提供充足养分。随着林木生长郁闭度增加,林分透光率降低,林下植被开始逐渐淘汰,有机质来源减少。油松人工林一般在造林15 a后开始进行抚育间伐作业[45],受抚育间伐影响,郁闭度降低,透光率提高,土壤微生物和土壤酶对间伐响应迅速,土壤生物活性提高[46]。进入中龄阶段(20 a)后,油松林进入快速生长期,林木间的竞争分化已缓和或基本停止,凋落物主要为难分解的油松凋落物,养分归还率慢,土壤微生物更多通过分解土壤原有有机物以满足林木生长对养分的需求,土壤容重和全氮出现了明显的负极化趋势[47],导致土壤有机碳降低至近40 a最低点。近熟林(>35 a)阶段,油松林土壤微生物及土壤酶酶活性明显改善,森林生态系统稳定性更强[43],林下植被得以恢复,种类更为丰富的凋落物及根系逐年积累,为土壤微生物新陈代谢和自身合成提供丰富的碳、氮以及能量来源[48],加之土壤容重改善以及土壤氮素供应能力提高,促进微生物对凋落物的分解转化,使土壤中有机碳逐步恢复和提高。
改革开放以来,中国林业政策不断演变[49],不同时期森林经营措施深刻影响着油松生长过程,导致各林龄油松林面积不断发生变化。1978−1983年,中国开展了全国范围的植树造林活动,但1984−1991年,在经济利益驱使下,过量采伐和大面积皆伐成为中国森林主要经营方式,导致该时期中龄林及近熟林面积占比较低,油松幼龄林面积达132.9×104 hm2,占总面积的64%。由于良好的土壤种植条件和整地措施,1980−1989年油松林0~60 cm土层土壤有机碳储量处于较高水平,为205.45 Tg。1992年起,中国开始实行一系列林业可持续发展战略,乱砍滥伐、毁林开荒和占用林地现象得到遏制,使得1990−1999年油松林总面积增加至243.1×104 hm2。但由于该时期幼龄林+近熟林面积占比降低,而中龄林面积增加至91.4×104 hm2,占比提高38%,抵消了总面积增加对土壤有机碳的增加效果,使土壤有机碳储量和前期相比,降低了6.81 Tg左右。2000年后中国进一步启动“天然林资源保护工程”,严禁采伐或大幅削减森林采伐量等更严格的森林保护措施,最大程度地保证油松生长,使2000−2009年中龄林面积进一步提高至103.9×104 hm2,占比达45%,加之该时期总面积出现小幅降低,上述双重因素的叠加作用导致该时期0~60 cm土层土壤有机碳储量出现较大幅度降低,达到近40 a的最低点,仅183.83 Tg。2010−2017年,虽然幼龄林面积持续降低,但经过近30 a自然生长,20世纪80年代种植的油松大部分进入近熟林阶段,近熟林面积达93.1×104 hm2,两者占比达59%,加之总面积增加至251.2×104 hm2,不仅抵消了该时期中龄林对土壤有机碳的降低作用,而且使0~60 cm土层土壤有机碳储量出现较大幅度增加,达247.02 Tg的最高值。
-
近40 a来,油松林土壤有机碳质量分数和储量变化表聚现象明显,0~20 cm土层是土壤有机碳质量分数或储量变化的主要贡献者,土壤有机碳质量分数和储量的占比分别达47%~54%和45%~50%。由于年均降水量和土壤黏化层对上层土壤可溶性有机碳输入和微生物活动制约以及死亡根系提供碳源有限,20~60 cm土层土壤有机碳质量分数和储量在各阶段始终处于相对稳定和较低的水平。油松林0~60 cm土层土壤有机碳质量分数和储量均呈先减少后增加的时间变化特征,其中,土壤有机碳储量从1980−1989年的205.45 Tg降低至2000−2009年最低点,为183.83 Tg,之后2010−2017年间出现较大幅度的增加,达到最高,为247.02 Tg。油松林地土壤性质、林分条件和各龄组面积比例受不同时期森林经营和保护措施深刻影响,土壤容重、土壤全氮和林分郁闭度是导致油松林土壤有机碳质量分数及储量变化的主要因素,随着近熟林面积比例的增加,预计未来仍会继续保持增长趋势。
Changes of soil organic carbon storage in Pinus tabulaeformis forest and its influencing factors
-
摘要:
目的 通过探究油松林Pinus tabulaeformis土壤有机碳质量分数和储量的垂直变化和时间变化特征,为油松林土壤碳储量预测和碳汇管理提供理论依据。 方法 基于1980−2017年文献数据,综合运用单因素方差分析、多重比较、相关性分析和通径分析等方法,探讨棕壤和褐土2种土壤类型下油松林土壤有机碳质量分数及储量变化特征,并结合不同时期中国森林经营措施和油松生长特征分析其驱动因素。 结果 油松林土壤有机碳质量分数和储量变化随土层深度增加而显著降低(P<0.05),0~20 cm土层是碳库的主要贡献层,占0~60 cm土层土壤有机碳储量的45%~50%;近40 a间土壤有机碳质量分数和储量呈先减少后增加的时间变化特征,其中,2000−2009年为最低点,而后出现较大幅度增加,在2017年达到储量最高点,为247.02 Tg。 结论 土壤容重、土壤全氮和林分郁闭度是油松林土壤有机碳质量分数(储量)变化的主要因素,不同时期森林经营和保护措施对三者的深刻影响是油松林土壤有机碳质量分数和储量呈现明显时间变化的重要原因。图3表5参49 Abstract:Objective The objective is to explore the vertical and temporal variation characteristics of soil organic carbon(SOC) content and storage in Pinus tabulaeformis forest, so as to provide a theoretical basis for soil carbon storage prediction and carbon sink management in P. tabulaeformis forest. Method Based on the literature data from 1980 to 2017, the characteristics of spatial and temporal variation for SOC content and storage in P. tabulaeformis forest under brown soil and cinnamon soil types were discussed using the methods of one-way ANOVA, multiple comparison, correlation analysis and path analysis, and the driving factors were analyzed combined with China’s forest management measures and growth characteristics of P. tabulaeformis in different periods. Result The change of SOC content and storage in P. tabulaeformis forest decreased significantly with the increase of soil depth (P<0.05), and 0−20 cm soil layer was the main contribution layer of carbon pool, accounting for 45%−50% of SOC storage in 0−60 cm soil layer. In the past 40 years, the content and storage of SOC showed the temporal variation characteristics of first decreasing and then increasing, among which 2000−2009 was the lowest, and then increased significantly, reaching the highest storage point of 247.02 Tg in 2017. Conclusion Soil bulk density, soil total nitrogen and stand canopy density are the main factors affecting the change of SOC content, and the profound effects of forest management and protection measures on the three factors are the important reasons for the significant temporal changes of SOC content and storage in P. tabulaeformis forest. [Ch, 3 fig. 5 tab. 49 ref.] -
表 1 不同时段油松林各龄组面积及占比
Table 1. Statistics of P. tabulaeformis forest area at different stages
龄组 1980−1989年 1990−1999年 2000−2009年 2010−2017年 面积/(×104 hm2) 占比/% 面积/(×104 hm2) 占比/% 面积/(×104 hm2) 占比/% 面积/(×104 hm2) 占比/% 幼龄林 132.9 64 102.2 42 74.6 32 55.0 22 中龄林 59.4 29 91.4 38 103.9 45 103.1 41 近熟林 13.8 7 49.5 20 54.7 23 93.1 37 总计 206.1 100 243.1 100 233.2 100 251.2 100 说明:林龄划分按照《中国森林(第1卷)》[20]标准,其中,近熟林为划分标准中的近熟林及以上林龄的油松林;不同时期幼龄林、中 龄林和近熟林的面积根据文献[21-27]统计数据计算获取  下载: 导出CSV
下载: 导出CSV
表 2 不同时段土壤有机碳质量分数统计分析
Table 2. Variation of soil organic carbon content at different stages
时段 0~20 cm有机碳/(g·kg−1) 20~40 cm有机碳/(g·kg−1) 40~60 cm有机碳/(g·kg−1) 0~60 cm有机碳平
均值/(g·kg−1)最大值 最小值 平均值 变异系数 最大值 最小值 平均值 变异系数 最大值 最小值 平均值 变异系数 1980−1989年 45.00 4.63 19.50 Aa 0.57 27.50 4.26 11.48 Ab 0.56 30.62 2.70 8.37 Abc 0.79 13.11 A 1990−1999年 41.93 4.11 15.00 Ba 0.56 44.66 1.34 10.09 Ab 0.86 22.33 0.68 6.15 Ac 0.67 10.41 B 2000−2009年 56.55 1.20 14.57 Ba 0.66 45.62 1.00 8.49 Ab 0.96 34.68 0.68 6.33 Abc 0.96 9.80 B 2010−2017年 57.45 0.50 20.76 Aa 0.55 35.28 0.74 11.36 Ab 0.62 30.20 0.65 7.94 Ac 0.68 13.35 A 说明:不同大写字母表示相同土层不同时段有机碳质量分数差异显著(P<0.05);不同小写字母表示相同时段不同土层土壤有机碳质 量分数差异显著(P<0.05)
下载: 导出CSV
表 3 不同时段土壤有机碳储量及占比
Table 3. Estimation of soil organic carbon storage in P. tabulaeformis forest at different stages
土层/cm 1980−1989年 1990−1999年 2000−2009年 2010−2017年 储量/Tg 占比/% 储量/Tg 占比/% 储量/Tg 占比/% 储量/Tg 占比/% 0~20 97.18 47.30 90.20 45.41 86.27 46.93 124.92 50.57 20~40 61.85 30.10 67.06 33.76 55.68 30.29 70.60 28.58 40~60 46.43 22.60 41.38 20.83 41.88 22.78 51.49 20.85 0~60 205.45 100.00 198.64 100.00 183.83 100.00 247.02 100.00
下载: 导出CSV
表 4 0~20 cm土层土壤有机碳质量分数与各因子间的Pearson相关性分析
Table 4. Pearson correlation analysis between 0−20 cm soil organic carbon and each factors
因子 有机碳 容重 全氮 碳氮比 pH 年均气温 年均降水量 凋落物量 林分密度 郁闭度 林龄 有机碳 1 容重 −0.774** 1 全氮 0.627** 0.424** 1 碳氮比 0.609** −0.425** 0.446** 1 pH 0.140** −0.084 0.212* 0.209* 1 年均气温 −0.052 0.037 −0.016** 0.259** 0.023 1 年均降水量 0.158 −0.126 0.059 0.031 −0.300* 0.339** 1 凋落物量 0.042 0.131 −0.031 0.144 −0.113 0.273* 0.228 1 林分密度 0.055 −0.147* −0.094 −0.232* 0.017 0.161 0.149 0.169 1 郁闭度 0.336** −0.159 0.246 0.221 0.048 0.047 0.090 0.220 −0.128 1 林龄 0.376** −0.297* 0.469** 0.454** 0.021 0.088 0.064 0.211 −0.455** 0.269** 1 说明:*表示显著相关(P<0.05);**表示极显著相关(P<0.01)
下载: 导出CSV
表 5 通径分析结果
Table 5. Results of path analysis of factors
因子 直接通径系数 间接通径系数 决定系数 容重 全氮 郁闭度 合计 容重 全氮 郁闭度 容重 −0.612 −0.259 0.097 −0.162 0.375 0.195 0.060 全氮 0.376 0.159 0.092 0.251 0.141 0.057 郁闭度 0.310 −0.049 0.076 0.027 0.096
下载: 导出CSV
-
[1] 李甜甜, 季宏兵, 孙媛媛, 等. 我国土壤有机碳储量及影响因素研究进展[J]. 首都师范大学学报(自然科学版), 2007, 28(1): 93 − 97. LI Tiantian, JI Hongbing, SUN Yuanyuan, et al. Advances in researches on soil organic carbon storages and affecting factors in China [J]. J Cap Norm Univ Nat Sci Ed, 2007, 28(1): 93 − 97. [2] POST W M, EMANUEL W R, ZINKE P J, et al. Soil carbon pools and world life zones [J]. Nature, 1982, 298(8): 156 − 159. [3] SCHLESINGER W H. Evidence from chronosequence studies for a low carbon-storage potential of soils [J]. Nature, 1990, 348(6298): 232 − 234. [4] PRENTICE K C, FUNG I Y. The sensitivity of terrestrial carbon storage to climate change [J]. Nature, 1990, 346(6279): 48 − 51. [5] ESWARAN H, van den BERG E, REICH P. Organic carbon in soils of the World [J]. Soil Sci Soc Am J, 1993, 57(1): 192 − 194. [6] 岳曼, 常庆瑞, 王飞, 等. 土壤有机碳储量研究进展[J]. 土壤通报, 2008, 39(5): 1173 − 1178. YUE Man, CHANG Qingrui, WANG Fei, et al. Research progress in soil organic carbon storage [J]. Chin J Soil Sci, 2008, 39(5): 1173 − 1178. [7] WANG Dan, WANG Bing, NIU Xiang. Effects of natural forest types on soil carbon fractions in northeast China [J]. J Trop For Sci, 2014, 26(3): 362 − 370. [8] 王棣, 耿增超, 佘雕, 等. 秦岭典型林分土壤有机碳储量及碳氮垂直分布[J]. 生态学报, 2015, 35(16): 5421 − 5429. WANG Di, GENG Zengchao, SHE Diao, et al. Soil organic carbon storage and vertical distribution of carbon and nitrogen across different forest types in the Qinling Mountains [J]. Acta Ecol Sin, 2015, 35(16): 5421 − 5429. [9] ZHOU Guoyi, XU Shan, CIAIS P, et al. Climate and litter C/N ratio constrain soil organic carbon accumulation [J]. Natl Sci Rev, 2019, 6(4): 746 − 757. [10] 黄晓强, 信忠保, 赵云杰, 等. 林龄和立地条件对北京山区油松人工林碳储量的影响[J]. 水土保持学报, 2015, 29(6): 184 − 190. HUANG Xiaoqiang, XIN Zhongbao, ZHAO Yunjie, et al. Effect of stand ages and site conditions on carbon stock of Pinus tabulaeformis plantations in Beijing mountainous area [J]. J Soil Water Conserv, 2015, 29(6): 184 − 190. [11] CHENG Xiaoqin, HAN Hairong, KANG Fengfeng, et al. Variation in biomass and carbon storage by stand age in pine (Pinus tabulaeformis) planted ecosystem in Mt. Taiyue, Shanxi, China [J]. J Plant Interactions, 2014, 9(1): 521 − 528. [12] DENG Lei, SHANGGUAN Zhouping. Afforestation drives soil carbon and nitrogen changes in China [J]. Land Degradation Dev, 2017, 28(1): 151 − 165. [13] 张文雯, 韩海荣, 程小琴, 等. 间伐对华北落叶松人工林土壤活性有机碳含量及酶活性的影响[J]. 应用生态学报, 2019, 30(10): 3347 − 3355. ZHANG Wenwen, HAN Hairong, CHENG Xiaoqin, et al. Effects of thinning on soil active organic carbon contents and enzyme activities in Larix principis-rupprechtii plantation [J]. Chin J Appl Ecol, 2019, 30(10): 3347 − 3355. [14] MA Junyong, KANG Fengfeng, CHENG Xiaoqin, et al. Moderate thinning increases soil organic carbon in Larix principis-rupprechtii (Pinaceae) plantations [J]. Geoderma, 2018, 329: 118 − 128. [15] 曹小玉, 李际平, 张彩彩, 等. 不同龄组杉木林土壤有机碳和理化性质的变化特征及其通径分析[J]. 水土保持学报, 2014, 28(4): 200 − 205. CAO Xiaoyu, LI Jiping, ZHANG Caicai, et al. Variation of contents of organic carbon and physic-chemical properties of soil and path analysis for their relations in different age-group chinese fir plantations [J]. J Soil Water Conserv, 2014, 28(4): 200 − 205. [16] 马钦彦, 谢征鸣. 中国油松林储碳量基本估计[J]. 北京林业大学学报, 1996, 18(3): 31 − 34. MA Qinyan, XIE Zhengming. Estomation of the carbon stored in Chinese pine forest [J]. J Beijing For Univ, 1996, 18(3): 31 − 34. [17] 孙维侠, 史学正, 于东升. 土壤有机碳的剖面分布特征及其密度的估算方法研究——以我国东北地区为例[J]. 土壤, 2003, 35(3): 236 − 241. SUN Weixia, SHI Xuezheng, YU Dongsheng. Distribution pattern and density calculation of soil organic carbon in profile [J]. Soils, 2003, 35(3): 236 − 241. [18] ADAMS W A. The effect of organic matter on the bulk and true densities of some uncultivated podzolic soils [J]. J Soil Sci, 1973, 24(1): 10 − 17. [19] 解宪丽, 孙波, 周慧珍, 等. 中国土壤有机碳密度和储量的估算与空间分布分析[J]. 土壤学报, 2004, 41(1): 35 − 43. XIE Xianli, SUN Bo, ZHOU Huizhen, et al. Organic carbon density and storage in soils of China and spatial analysis [J]. Acta Pedol Sin, 2004, 41(1): 35 − 43. [20] 中国森林编辑委员会. 中国森林: 第1卷[M]. 北京: 中国林业出版社, 1997. [21] 中华人民共和国林业部. 全国森林资源统计(1984−1988)[M]. 北京: 中国林业出版社, 1989. [22] 国家林业和草原局. 第四次全国森林资源清查主要结果(1989−1993年)[EB/OL]. 2000-12-30[2021-03-20]. http://www.forestry.gov.cn/main/65/content-555100.html. [23] 国家林业和草原局. 第五次全国森林资源清查主要结果(1994−1998年)[EB/OL]. 2000-06-13[2021-03-20]. http://www.forestry.gov.cn/main/65/20000613/554361.html. [24] 国家林业和草原局. 第六次全国森林资源清查主要结果(1999−2003年)[EB/OL]. 2006-09-28[2021-03-20]. http://www.forestry.gov.cn/main/65/20060928/90.html. [25] 国家林业和草原局. 第七次全国森林资源清查主要结果(2004−2008年)[EB/OL]. 2010-01-28[2021-03-20]. http://www.forestry.gov.cn/main/65/20100128/326341.html. [26] 国家林业局. 中国森林资源报告(2009−2013)[M]. 北京: 中国林业出版社, 2014. [27] 国家林业和草原局. 中国森林资源报告(2014−2018)[M]. 北京: 中国林业出版社, 2019. [28] 刘广深, 徐冬梅, 许中坚, 等. 用通径分析法研究土壤水解酶活性与土壤性质的关系[J]. 土壤学报, 2003, 40(5): 756 − 762. LIU Guangshen, XU Dongmei, XU Zhongjian, et al. Relationship between hydrolase activity in soils and soil properties in Zhejiang Province [J]. Acta Pedol Sin, 2003, 40(5): 756 − 762. [29] 王丹, 王兵, 戴伟, 等. 杉木人工林土壤系统有机碳相关变量的通径分析[J]. 土壤通报, 2011, 42(4): 822 − 827. WANG Dan, WANG Bing, DAI Wei, et al. Path analysis on the correlative variables of organic carbon in soil system of Chinese fir plantations [J]. Chin J Soil Sci, 2011, 42(4): 822 − 827. [30] 张维俊, 李双异, 徐英德, 等. 土壤孔隙结构与土壤微环境和有机碳周转关系的研究进展[J]. 水土保持学报, 2019, 33(4): 1 − 9. ZHANG Weijun, LI Shuangyi, XU Yingde, et al. Advances in research on relationships between soil pore structure and soil miocroenvironment and organic carbon turnover [J]. J Soil Water Conserv, 2019, 33(4): 1 − 9. [31] 逯军峰, 王辉, 曹靖, 等. 油松人工林凋落物对土壤理化性质的影响[J]. 西北林学院学报, 2007, 22(3): 25 − 28. LU Junfeng, WANG Hui, CAO Jing, et al. Effect of forest litiers on soil physical and chemical properties in Pinus tabulaeformis plantation [J]. J Northwest For Univ, 2007, 22(3): 25 − 28. [32] BENGTSSON G, BENGTSON P, MANSSON K F. Gross nitrogen mineralization, immobilization, and nitrification rates as a function of soil C/N ratio and microbial activity[J]. Soil Biol Biochem, 2003, 35(1): 143. doi: 10.1016/S0038-0717(02)00248-1. [33] 徐波, 朱忠福, 李金洋, 等. 九寨沟国家自然保护区4个典型树种叶片凋落物在林下及高山湖泊中的分解及养分释放特征[J]. 植物生态学报, 2016, 40(9): 883 − 892. XU Bo, ZHU Zhongfu, LI Jinyang, et al. Leaf decomposition and nutrient release of dominant species in the forest and lake in the Jiuzhaigou National Nature Reserve, China [J]. Chin J Plant Ecol, 2016, 40(9): 883 − 892. [34] 刘勇, 李国雷. 不同林龄油松人工林叶凋落物分解特性[J]. 林业科学研究, 2008, 21(4): 500 − 505. LIU Yong, LI Guolei. Decomposition characteristics affected by forest age in Pinus tabulaeformis plantations [J]. For Res, 2008, 21(4): 500 − 505. [35] 何姗, 刘娟, 姜培坤, 等. 经营管理对森林土壤有机碳库影响的研究进展[J]. 浙江农林大学学报, 2019, 36(4): 818 − 827. HE Shan, LIU Juan, JIANG Peikun, et al. Effects of forest management on soil organic carbon pool: a review [J]. J Zhejiang A&F Univ, 2019, 36(4): 818 − 827. [36] 李国雷, 刘勇, 李瑞生, 等. 油松叶凋落物分解速率、养分归还及组分对间伐强度的响应[J]. 北京林业大学学报, 2008, 30(5): 52 − 57. LI Guolei, LIU Yong, LI Ruisheng, et al. Responses of decomposition rate, nutrient return and composition of leaf litter to thinning intensities in Pinus tabulaeformis plantation [J]. J Beijing For Univ, 2008, 30(5): 52 − 57. [37] 杨丽丽, 王彦辉, 杜敏, 等. 六盘山典型森林伴随降水的总有机碳(TOC)通量变化特征[J]. 生态学报, 2014, 34(21): 6297 − 6308. YANG Lili, WANG Yanhui, DU Min, et al. The variation of total organic carbon (TOC) flux with rain water in regional forests of Liupan Mountains, China [J]. Acta Ecol Sin, 2014, 34(21): 6297 − 6308. [38] ZHOU Zhiyong, ZHANG Zhiqiang, ZHA Tonggang, et al. Predicting soil respiration using carbon stock in roots, litter and soil organic matter in forests of Loess Plateau in China [J]. Soil Biol Biochem, 2013, 57: 135 − 143. [39] QI Li, WANG Shuai, ZHUANG Qianlai, et al. Spatial-temporal changes in soil organic carbon and pH in the Liaoning Province of China: a modeling analysis based on observational data [J]. Sustainability, 2019, 11(13): 3569. [40] 任丽娜, 王海燕, 丁国栋, 等. 林分密度对华北土石山区油松人工林土壤有机碳及养分特征的影响[J]. 干旱区地理, 2012, 35(3): 456 − 464. REN Lina, WANG Haiyan, DING Guodong, et al. Effects of Pinus tabulaeformis Carr. plantation density on soil organic carbon and nutrients characteristics in rocky mountain area of northern China [J]. Arid Land Geogr, 2012, 35(3): 456 − 464. [41] 国家林业局. 油松人工林经营技术规程: LY/T 2971−2018[S]. 北京: 中国标准出版社, 2018. [42] 中国科学院南京土壤研究所. 中国土壤[M]. 北京: 科学出版社, 1978. [43] 范媛媛, 李懿, 李启迪. 不同林龄油松土壤微生物、酶活性和养分特征[J]. 水土保持研究, 2019, 26(6): 58 − 64. FAN Yuanyaun, LI Yi, LI Qidi. Microbe, enzymatic activity and nutrient contents of soils in different stand ages of Pinus tabuliformis [J]. Res Soil Water Conserv, 2019, 26(6): 58 − 64. [44] 庞学勇, 刘庆, 刘世全, 等. 川西亚高山云杉人工林土壤质量性状演变[J]. 生态学报, 2004, 24(2): 261 − 267. PANG Xueyong, LIU Qing, LIU Shiquan, et al. Changes of soil fertility quality properties under subalpine spruce plantation in Western Sichuan [J]. Acta Ecol Sin, 2004, 24(2): 261 − 267. [45] 中国树木志编委会. 中国主要树种造林技术[M]. 北京: 中国林业出版社, 1981: 103 − 106. [46] 李国雷, 刘勇, 李俊清, 等. 油松飞播林土壤质量评判及其调控[J]. 南京林业大学学报(自然科学版), 2008, 32(3): 19 − 24. LI Guolei, LIU Yong, LI Junqing, et al. Evaluation and regulation of soil quality of aerial seeding Pinus tabulaeformis stands [J]. J Nanjing For Univ Nat Sci Ed, 2008, 32(3): 19 − 24. [47] 冯顺煜, 刘增文, 段而军, 等. 陕北半湿润黄土丘陵区人工纯林土壤中微量元素含量及其极化现状[J]. 西北林学院学报, 2009, 24(2): 32 − 36. FENG Shunyu, LIU Zengwen, DUAN Erjun, et al. Contents of trace-elements and polarizabilities of the soil of manmade pure forest soil semihumid loessial hilly area, Northern Shaanxi [J]. J Northwest For Univ, 2009, 24(2): 32 − 36. [48] 高晶, 韩海荣, 康峰峰, 等. 冀北辽河源不同林龄油松天然次生林土壤微生物生物量及酶活性[J]. 东北林业大学学报, 2015, 43(9): 78 − 83. GAO Jing, HAN Hairong, KANG Fengfeng, et al. Soil microbial biomass and enzymes activity of Pinus tabulaeformis forest with different ages in Liaohe River of Northern Hebei [J]. J Northeast For Univ, 2015, 43(9): 78 − 83. [49] 潘丹, 陈寰, 孔凡斌. 1949年以来中国林业政策的演进特征及其规律研究——基于283个涉林规范性文件文本的量化分析[J]. 中国农村经济, 2019(7): 89 − 108. PAN Dan, CHEN Huan, KONG Fanbin. The evolution of forestry policies since 1949: an quantitative analysis based on 283 texts of forest-related normative policy document [J]. Chin Rural Econ, 2019(7): 89 − 108. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210390
 点击查看大图
点击查看大图
计量
- 文章访问数: 1798
- HTML全文浏览量: 438
- PDF下载量: 44
- 被引次数: 0





