





-
高等植物是由根、枝、叶、花、果等基本构件单位不断重复增加而形成的构件生物[1-2],其外部形态具有高度可塑性,这种可塑性与植物对环境的适应性密切联系,是植物适应异质环境的重要手段[3-6]。通过观察植物构件特征变化,可以了解植物个体构件结构的复杂程度和其对空间的占据及利用潜力。枝是植物重要的构件单位,为植物生长输导水分和养分,其伸长和分枝有利于叶和花扩大生存空间,并达到生长的最佳位置,同时,枝长、枝径粗、分枝角度、分枝数等的不同组合,将直接影响植株对空间、光等资源的利用和适应策略[7]。叶是植物进行光合作用的重要器官,叶片数能直接或间接影响植物枝构件生长,叶面积大小影响着植物对光能的截获和利用,影响植物生产力[8-9]。目前,关于植物构件特征与环境关系的研究不断增多,研究对象包括乔木、灌木、草本。如闫宝龙等[10]对大兴安岭南段白桦Betula platyphylla种群构件研究,程伟等[11]对石山和土山2种生境条件下西南桦Betula alnoides的生殖构件及其空间格局进行研究,于应文等[12]分析了主要气候因子(气温、日照和降水)对混播草地中黑麦草Lolaum perenne,白三叶Trifolaum repens和白苞蒿Artemisia lactiflora等构件生长的影响,皆是从构件水平研究植物与环境的关系,揭示植物对环境的适应机制。但当前涉及水麻Debregeasia orientalis的研究较少,仅有水麻扦插实验、野生果树资源研究、水麻化学成分及抗菌作用分析等[13-15],缺乏对水麻在不同生境的生长发育和适应策略以及水麻在环境中所起功能作用的研究。水麻,为荨麻科Urticaceae水麻属Debregeasia植物,性喜湿耐阴,树势健壮,易繁殖,丰产性强,主要分布在中国亚热带中部及北部、秦岭南坡,对坡面的负荷小,能防止浅层土壤崩塌,是水土保持优良树种,具有重要的生态功能[16-17],在解决20世纪50年代四川农业大学老板山水土流失问题中发挥了重要作用。当前,水麻已成为雅安市老板山优势植被类型。本研究比较和分析不同生境下水麻枝叶构件特征的变化,旨在探究环境对水麻生长的影响以及水麻枝叶构件对环境的适应对策和进化机制,为进一步增强水土保持功能提供依据。
HTML
-
老板山(28°30'~30°33'N,101°13'~101°33'E)位于雅安市雨城区四川农业大学内,是四川农业大学的人文地标,山体成东西走向,东西全长约1 300 m,南北最宽约430 m,海拔为592~735 m。该区属亚热带湿润季风气候,降水集中于夏季,多夜雨,年平均气温为16.2 ℃,1月平均气温为6.1 ℃,7月平均气温为25.4 ℃,年降水量达2 000.0 mm,年平均蒸发量为838.8 mm,平均湿度为79%,土壤为紫色土,呈微酸性。研究区内植被为亚热带常绿阔叶林,覆盖率95%以上,乔木层主要是以慈竹Neosinocalamus affinis,水杉Metasequoia glyptostroboides,柳杉Cryptomeria fortunei,樟树Cinnamomum camphora等为主要树种,所研究的灌木层以水麻为主,伴生植物主要有毛竹Phyllostachys edulis,灯台树Bothrocaryum controversum等,草本层包括鸭跖草Commelina communis,土牛膝Achyranthes aspera和蕨类等植物。历史上,老板山水土流失问题严重,乔木在坡度较大、土层覆盖率较低的地方难以生长。以水麻为主的灌木在水土保持,固土护坡方面发挥了重要作用。
-
通过实地调查,根据水麻生长的地理位置以及造成个体构件生存策略转变的自身生长类型的不同,在雅安市四川农业大学老板山实践基地选取林内单株、林缘丛生、沟边丛生、林内丛生等4个不同生境类型样地(表 1),每个生境选取生长良好且具有代表性的水麻各12株进行定株调查,测定其地径(距地面3.00 cm处直径)、株高、冠幅。枝级划分:运用Strahler法[18]确定枝级。枝叶构件指标的测定:测量其枝条数、枝条长度、枝径粗、分枝角度;统计各级枝的叶片数,同时在各级枝上随机选取30片大小不等的叶片,利用Photoshop CS5软件测定叶面积。
生境类型 海拔/m 生境郁闭度 土壤类型 平均高度/cm 平均地径/cm 伴生植物 林内单株 663 0.7 紫色土 295 3.09 水杉,柳杉,构树 Browssonetia papyrifera 林缘丛生 625 0.4 紫色土 346 3.78 柳杉,灯台树 沟边丛生 627 0.1 紫色土 386 2.46 鸭妬草,棠叶悬钩子 Rubus malifolius 林内丛生 658 0.7 紫色土 306 3.47 水杉,柳杉,朴树 Celtis sinensis Table 1. Condition of habitats and growth of Debregeasia orientalis
-
依据吕志伟等[19]对植物枝构件特征的运算方法计算枝径比,采用WHITNEY[20]提出并经STEINGRAEBER等[21]修改的方法计算逐级分枝率。枝径比是指植物枝与枝之间枝条直径之比;逐级分枝率是植物某一级枝与其下一个高级枝的枝数比。
其中:RBD为枝径比;Di为第i级枝直径;Di+1为第i+1级枝直径;SBR为逐级分枝率;Ni+1为第i+1级枝数目;Ni为第i级枝数目。运用Excel 2010和DPS 15.10进行数据统计和处理,用Origin 9.0制图。
2.1. 样地设置与调查
2.2. 数据处理
-
分枝数与枝条发展潜力密切相关,体现着植物对空间资源的获取和利用潜力。由表 2可知:林缘丛生的水麻一级枝、二级枝、三级枝、四级枝分枝数均大于其他生境,生长潜力大。沟边丛生水麻无三级枝、四级枝,一级枝、二级枝分枝数与其他生境水麻同级分枝数存在显著差异(P<0.05),其他生境水麻各级枝数均表现为一级枝>二级枝>三级枝>四级枝,且一级枝到二级枝分枝数变幅较大,表明水麻以一级枝为主。
枝构件 生境类型 一级枝 二级枝 三级枝 四级枝 分枝数 林内单株 134.08 ± 13.71 a 26.08 ± 2.77 a 4.08 ± 0.76 ab 2.00 ± 1.00 a 林缘丛生 148.08 ± 20.32 a 29.75 ± 4.25 a 6.55 ± 2.02 a 2.00 ± 1.50 a 沟边丛生 47.75 ± 6.15 b 6.58 ± 0.88 b 林内丛生 142.58 ± 29.72 a 24.42 ± 2.64 a 3.64 ± 0.80 ab 1.50 ± 0.50 a 分枝长度/cm 林内单株 65.85 ± 11.53 a 50.91 ± 3.99 a 38.87 ± 2.21 b 17.50 ± 0.87 c 林缘丛生 52.03 ± 7.49 a 50.89 ± 3.07 a 44.14 ± 1.50 ab 56.00 ± 5.49 b 沟边丛生 65.94 ± 13.72 a 58.43 ± 3.10 a 林内丛生 61.40 ± 8.78 a 57.12 ± 4.90 a 50.67 ± 2.40 a 86.00 ± 4.14 a 分枝角度/(°) 林内单株 59.59 ± 0.95 a 59.05 ± 1.75 a 62.03 ± 1.47 a 45.00 ± 1.12 a 林缘丛生 56.20 ± 1.14 ab 56.43 ± 2.04 a 54.13 ± 4.03 a 58.33 ± 1.20 a 沟边丛生 54.73 ± 1.54 b 51.46 ± 1.83 a 林内丛生 56.97 ± 1.07 ab 54.60 ± 2.23 a 59.74 ± 3.08 a 52.67 ± 1.20 a 说明:数据为平均值±标准误。不同小写字母表示不同生境同一枝级差异显著(P<0.05) Table 2. Branching module of Debregeasia orientalis in different habitats
-
分枝长度是测定枝质量的重要指标,影响着叶和芽的空间分布,能间接表明植物枝系吸收利用光能和二氧化碳的能力,反映植物生长活力的强弱和对环境的适应对策。表 2表明:不同生境水麻四级枝分枝长度差异显著(P<0.05),其他枝级差异不明显。一级枝分枝长度变化为沟边丛生>林内单株>林内丛生>林缘丛生,二级枝为沟边丛生>林内丛生>林内单株>林缘丛生,三四级枝分枝角度变化均为林内丛生>林缘丛生>林内单株。沟边丛生的水麻无三四级分枝,一二级枝有充足时间进行生长,分枝长度为65.94和58.43 cm,优于其他生境同级枝条;在同一生境中,林内单株水麻分枝长度随着枝级的增加逐渐减小,林缘和林内丛生水麻四级枝较上级枝长,这可能与萌生枝所处位置以及获取资源能力有关。
-
分枝角度指植株相邻枝级间的夹角,是枝条向空间拓展的形式之一,影响枝条的空间分布格局,反映植株对光照这一生境因子的利用。树木枝条的夹角主要受遗传基因控制,但不同生境中空间、光资源的不同以及各级枝为争夺资源等有利环境所产生竞争干扰程度不同,分枝角度也会发生相应的变化。由表 2可知:在各生境中所调查的水麻分枝角度整体变化范围为45.00°~63.00°,为中等分枝角度类型,变化幅度较小。不同生境下同一枝级水麻分枝角度差异不显著(P>0.05)。一级分枝角度变化趋势为林内单株>林内丛生>林缘丛生>沟边丛生,二级枝分枝角度为林内单株>林缘丛生>林内丛生>沟边丛生,三级枝分枝角度呈现出林内单株>林内丛生>林缘丛生的变化,四级枝则为林缘丛生>林内丛生>林内单株;一般而言,分枝角度越大,表明植物的空间扩展和利用能力越强。同一生境内,林内单株水麻三级枝分枝角度为62.03°,林缘丛生水麻四级枝条分枝角度为58.33°,沟边丛生水麻一级枝分枝角度为54.73°,林内丛生水麻三级枝分枝角度为59.74°。沟边丛生水麻各级枝条分枝角度相对于其他生境较小,分枝角度表现为收敛型。
-
枝径比能够反映植株不同级别枝条之间的承载能力以及相邻枝条水分和养分的运输情况。一般而言,枝径比与相邻枝间运输效率(水分、养分)呈正相关,枝径比越大,水分、养分运输效率越高,枝条生长发育得越好。由图 1可知:在不同生境中,沟边丛生水麻一二级枝枝径比(RBD2:1)为2.08,枝条承载力最强,其他生境RBD2:1差异不明显。林内丛生水麻二三级枝枝径比(RBD3:2)大于林内单株和林缘丛生水麻,林缘丛生水麻三四级枝枝径比(RBD4:3)最大,这与不同生境中光、热、水资源分布不同,植株所采取的生存策略存在差异有关。在同一生境中,枝径比均为RBD3:2>RBD2:1>RBD4:3,二三级枝之间水分、养分运输效率高,生长优势好。

Figure 1. Ratio of branching diameter of D. orientalis in different habitats
-
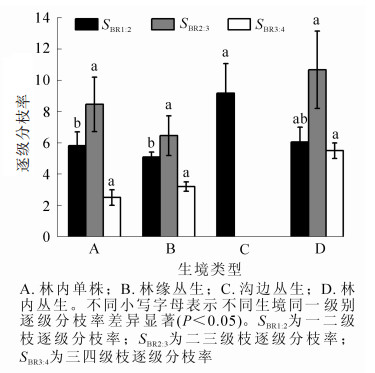
逐级分枝率是构型研究的重要指标,能反映枝条分枝能力以及各枝级间枝条数量配置状况和资源的分配状况。植株的逐级分枝率越小,枝条的分枝数就越多,分枝能力也越强,占据空间、获取资源的能力就越大。由图 2可知:沟边丛生水麻一二级枝逐级分枝率(SBR1:2)为9.17,且与其他生境差异显著(P<0.05),不同生境水麻二三级枝逐级分枝率(SBR2:3)表现为林缘丛生<林内单株<林内丛生,三四级枝逐级分枝率(SBR3:4)则为林内单株<林缘丛生<林内丛生。表明在不同生境中,水麻各级枝条对空间异质性的响应不同;在同一生境中,各级枝条逐级分枝率变化趋势一致,SBR3:4<SBR1:2<SBR2:3,表明水麻在二三级枝间分枝能力较强。

Figure 2. Branching ratios of D. orientalis in different habitats
-
植物能进行光合作用主要靠叶。作为光合器官,叶片数量、叶面积大小以及其配置等都会对叶片接受光量、光强等产生影响,从而决定植物光能利用效率,影响植物生长。叶在进行光合作用时,叶片数能够直接或间接影响植物的植株高度、基径、枝和其上叶质量。如果叶的数量和面积处于劣势,那么必然导致植物在种内、种间竞争时生长发育受阻。
从叶片数来看(图 3),不同生境各级枝叶片数差异不显著(P>0.05)。在不同生境中,一级枝叶片数,林缘丛生>林内单株>林内丛生>沟边丛生,二级枝叶片数,林内丛生>林内单株>沟边丛生>林缘丛生,三级枝上叶片数变化较小;在同一生境中,各级枝叶片数变化趋势一致,为一级枝>二级枝>三级枝。

Figure 3. Leaves module characteristics of Debregeasia orientalis in different habitats
叶面积的大小与植株利用光能的多少呈正比,但也有人认为,在光照不充足时,叶片会通过增大叶面积来获取更多的光环境资源,进行光合作用,促进植物光合产物的积累。不同生境下,一级枝上叶面积呈现出沟边丛生>林缘丛生>林内丛生>林内单株,二级枝叶面积林缘丛生>林内单株>林内丛生>沟边丛生(图 3);在同一生境下,不同枝级水麻总体上表现出二级枝叶面积较一级枝和三级枝叶面积大,这既是构件种群内部竞争引起的结果,亦是植物在不同生育时期,不断调整各构件个体间生长和营养物质的分配,以满足其他构件个体生长的结果。
3.1. 不同生境水麻枝构件特征
3.1.1. 分枝数
3.1.2. 分枝长度
3.1.3. 分枝角度
3.1.4. 枝径比
3.1.5. 逐级分枝率
3.2. 不同生境水麻叶构件特征

-
植物的构件特征是植物在特定环境下的形态反映,与植物内部生理生化性质关系密切,其枝系构件主要由分枝角度、分枝数、分枝长度及分枝率等组成,是连接其他构件(叶、芽)的中介,它决定着植物叶空间分布,影响植物光合作用、水分和养分的运输与吸收,对植物适应环境变化至关重要[22]。本研究在对老板山水麻枝叶构件调查分析中发现,不同生境以及不同植物生长类型(单生、丛生)都会对水麻枝系构件产生影响。林缘丛生生境土壤肥沃,光热水资源充足,水麻分枝数量大,枝条发育良好,生长旺盛。这与赵友华等[23]发现适度遮光可以使栲树Castanopsis fargesii分枝数增多的结论相悖,三四级枝分枝数最少,但其叶、芽密度最大,最有发展潜力[24];沟边丛生生境土壤肥力较低,土层较薄,无三四级枝生长,但其一二级枝分枝长度明显超过其他生境,原因可能是水麻作为沟边生境绝对的优势种,竞争资源,获取养分的优势明显且自身养分仅供一二级枝生长,无三四级枝对资源进行竞争;分枝角度受光照、水分及空间资源的影响,在一定的空间内,植物通过改变自身角度将枝伸展到最大利用空间资源的位置来适应环境[25]。林缘丛生和沟边丛生生境水麻分枝角度较小,主要在于这2种生境皆位于林地边缘,空间范围广,光照资源充足,水麻只有通过减小分枝角度,来最大限度地获取光照,促进植株生长;林内单株和丛生的水麻,因水杉、柳杉等乔木对光照的截获,林内光资源匮乏,水麻必须通过扩大分枝角度,占据空间资源,促进植株生长,这与贾程等[8]在构件研究中得出的结论一致。同一生境内,水麻二三级枝枝径比(RBD3:2)最大,各级别逐级分枝率SBR3:4<SBR1:2<SBR2:3,二三级枝水分、养分运输效率高,分枝能力强,生长优势好。实地调查显示,水麻生长的沟边生境,土壤固结,土层紧实,主要原因可能是水麻根系固土护坡作用强且树冠在林内形成二次降水,降低雨水对坡面的直接冲刷小,水土流失较少。
叶片通过光合作用,可为植物生长发育提供营养物质和储存能量,与植物对环境的利用有紧密关系[26],植物叶片数和叶面积是其在环境中竞争能力的表现。本研究中林缘丛生水麻叶片数最多,与分枝数变化规律一致。谢元贵等[25]在研究桃叶杜鹃Rhododendron annae种群构件特征时也得出相似结论,即分枝数的增加导致构件种群内部竞争更加激烈,为更多地弥补因竞争而损失的物质能量积累,植物又通过增加叶片数量以充分的利用资源环境[27];叶片面积的大小不仅与外界环境密切相关,还与植株内部构造及叶的发育期有关。沟边和林缘丛生水麻叶面积较大,这主要与植株利用光能的多少呈正比。沟边和林缘生境皆不受乔木层影响,光照充足,光热条件较好,利于叶片进行充足的光合利用,物质积累和代谢增加,叶面积增大;林内丛生水麻叶面积比林内单株大,这也说明适当的遮光可使叶面积增大,以最大限度利用光能,进行光合作用。另外,在同一生境中,叶面积二级枝>一级枝>三级枝,这是不同叶片发育期,光合速率存在差异,老枝上的叶所占据的营养物质减少,促进新叶叶面积增长,避免叶自身竞争,这与刺五加Acanthopanax senticosus种群的生长状况相似。同时,分枝数的多少关系着植物内部的竞争,对叶面积的大小也存在一定影响[27]。
水麻易繁殖,丛生比单株生长良好。在乔木无法生长、易滑坡的坡面,水麻护坡、固土作用强。通过分析比较不同生长类型水麻对不同生境光照、空间等资源的利用和适应策略,对水麻进行科学管理,以最大限度获取生境资源,实现生态效益的最优化。
 DownLoad:
DownLoad: