

 下载:
下载:
-
森林是陆地生态系统中具最大生物量和生物生产力的生态系统,是地球生物圈过程的重要参与者[1]。森林生态系统为人类社会提供了丰富的物质产品和文化产品[2],更为维护生态系统平衡发挥了水源涵养、土壤保育等多种调节服务功能[3]。这些由森林生态系统与生态过程所形成及维持的人类赖以生存的自然环境条件与效用即为森林生态系统服务功能[4]。COSTANZA等[5]最早对全球16类生态系统服务价值进行了估算,其中将森林生态系统分为热带和温带进行价值估算。此后,国内外学者聚焦于森林生态系统服务功能概念内涵[6]和方法应用[7]进行了研究,国内获得了森林生态连清技术体系[8]、生态系统生产总值(GEP)核算[9]、国标GB/T 3858—2020《森林生态系统服务功能评估规范》[10]等研究成果。现阶段,国内许多学者基于此国标,采用森林资源规划设计调查(“二类”调查)数据,对省[11]、市[12]、县[13],以及自然保护地[14]、典型森林生态系统[15]等各层级的森林生态系统开展了服务功能核算。鉴于采用“二类”调查数据能够将核算结果落实到山头地块,从生态区位[16]、空间格局[17]等角度切入,细致探究森林生态系统服务功能机制也逐渐成为学界关注的焦点。
森林资源数据是整项核算工作的最基本数据。近些年,基于“二类”调查数据对各层级森林生态系统服务功能核算研究越来越多,但“二类”调查数据没有调查精度控制[18],累加后形成的上级行政区森林资源结果亦无精度控制[19]。如何对基于“二类”调查数据的森林生态系统服务功能核算结果进行精度控制,并没有在现有研究中得到重视。使用森林资源连续清查(“一类”清查)数据,通过系统抽样统计方法可使核算结果具有很高的精度保证和较好可信度[20]。鉴于物质量核算能够比较客观地反映森林生态系统的生态过程[15],本研究选取浙江省湖州市为研究地,使用“一类”清查样地数据和“二类”调查小班数据,将样地宏观核算和小班微观核算紧密结合,构建森林生态系统服务功能物质量市县联动核算体系,以期为解决上述问题提供参考。另外,也能够在把握市、县森林生态系统服务功能总体趋势的同时,为多层级核算结果的精度保证提供参考和借鉴。
-
湖州市位于浙江省最北端,辖吴兴区、南浔区、长兴县、德清县和安吉县,土地总面积5 824 km2。境内地形地貌以低山丘陵、湖积和冲积平原为主,地势大致呈现由西向东北倾斜的走势。全境为亚热带季风性气候,年均气温为12.2~17.3 ℃,多年平均降水量为761~1 780 mm。根据2018年森林资源监测结果(表1),湖州市林地面积为2 891.7 km2,其中森林面积为2 802.1 km2 (含一般灌木林面积)。森林蓄积量为759.1万 m3,森林覆盖率为48.1%。
表 1 湖州市2018年森林面积、蓄积量及其占比
Table 1. Forest areas, inventory and percentages of Huzhou City in 2018
县(区) 森林面
积/km2森林面积
占比/%森林蓄积量/
(×104 m3)森林蓄积量
占比/%吴兴区 329.4 11.7 127.4 16.8 南浔区 69.8 2.5 8.0 1.1 德清县 400.0 14.3 77.7 10.2 长兴县 678.9 24.2 212.7 28.0 安吉县 1 324.0 47.3 333.3 43.9 总计 2 802.1 100.0 759.1 100.0 -
森林生态系统服务功能包括供给服务、调节服务、文化服务和支持服务[21]。根据国标GB/T 38582—2020《森林生态系统服务功能评估规范》,本研究对湖州市森林生态系统的水源涵养、土壤保育、固碳、林木养分固持和提供负离子等5项服务功能物质量进行市县联动核算与精度控制研究,指标选取说明见表2。
表 2 森林生态系统服务功能指标体系及本研究指标选取说明
Table 2. Index system of forest ecosystem services function and explanation of index selection
服务类别 功能类别 指标类别 说明 供给服务 生物多样性 物种资源保育 主要通过野生动植物资源调查成果核算,故未纳入 林木产品供给 木材产品 此2项指标的产量一般是统计数据,与本研究目的不一致,故未纳入 非木材产品 调节服务 水源涵养 调节水量 此2项指标物质量核算方法与结果均一致,仅在评估价值量时,分别选取
不同单价。在实际应用中,水源涵养物质量也仅指调节水量。因此,本
研究选择调节水量一项指标,且后文中的水源涵养表述仅指代调节水量净化水质 固碳释氧 固碳 此2项指标核算方法差异仅体现在转换参数不同,而且,释氧量可通过固
碳量直接乘以氧气(O2)与碳(C)的相对原子质量比得到。因此,本研究
仅选择固碳一项指标释氧 净化大气环境 提供负离子 由于森林单位面积吸收气体污染物及滞纳TSP等的实测参数不足,本研究
仅选择提供负离子一项指标吸收气体污染物 滞纳TSP、PM10、PM2.5 森林防护 防风固沙 研究区不属于风力侵蚀区域。农田防护功能仅针对农田境界外100 m范围
内的森林。且2类调查数据未能界定森林是否为农田防护林,因此未选
取该2项指标农田防护 文化服务 森林康养 森林康养 森林康养价值一般是统计数据,或通过条件价值法等方法核算。与本研
究目的不一致,未选取该指标支持服务 保育土壤 固土 考虑到保肥量仅是固土量乘以土壤营养元素百分比含量(为常数)得到,固
土量核算精度决定了保肥量核算精度。因此,本研究仅选择固土指标。
且后文中的保育土壤表述仅指代固土保肥 林木养分固持 氮、磷、钾固持 该指标反映了林木吸收氮、磷、钾等营养元素并储存在体内的功能,因
此,本研究选择此3项指标,并以此3项指标物质量之和作为林木养分
固持物质量 -
“一类”清查数据源于湖州市961个样地的现场调查。“二类”调查数据源于2018年湖州市县级森林资源监测,该数据由2016年全面调查完成的森林资源数据年度更新形成。
数字高程模型(DEM)来源于地理空间数据云,采用ArcGIS 10.2重分类至分辨率10 m。年降水量和蒸发量数据来源于中国气象数据网,通过反距离权重法插值形成栅格数据,分辨率为10 m。地表径流系数参考文献[22]。土壤数据来源于浙江省土壤普查成果。植被净初级生产力采用2018年浙江省4 319个公益林固定样地监测成果。林分负离子量采用2018年浙江省62个空气负离子监测点测定成果。林木的氮、磷、钾含量参考文献[23]。
-
市县联动核算体系由1个市级总体和5个县级副总体组成。市级总体为湖州市,县级副总体以县(区)为单位进行划分。
-
①样地核算体系(YD)。该体系分为市级总体样地核算(YD_S)和县级副总体样地核算(YD_X)2个子体系,采用“一类”清查数据,按系统抽样核算方法[24],将单个样地的森林生态系统服务功能物质量推算到市、县级尺度并提供精度控制。2016—2018年,湖州市在477个省级样地(该477个样地是湖州市域范围内的浙江省级样地,以4 km×6 km间距布设)基础上,再按95%的可靠度,全市森林面积估计精度达到90%以上、活立木蓄积量估计精度达到85%以上的设计要求,以2 km×3 km间距加密布设484个市级样地,共计布设样地961个。其中,吴兴区134个、南浔区121个、德清县158个、长兴县236个、安吉县312个。单个样地面积为0.08 hm2,形状为正方形。②小班核算体系(XB)。该体系分为市级总体小班汇总(XB_S)和县级副总体小班核算(XB_X)2个子体系,采用“二类”调查数据,将各小班的森林生态系统服务功能物质量累加得到县级副总体核算结果,将各县级副总体核算结果累加得到市级总体汇总结果。
-
①同步核算机制。第一,市对县森林面积控制检验。森林面积对森林生态系统服务功能有着较高影响[25],按照张国江等[18]提出的方法进行市对县森林面积控制检验。市级森林面积核算结果估计精度92.99%,置信区间为250 772~288 524 hm2,县级副总体森林面积之和落在置信区间内,表明其森林面积有精度保证。第二,核算体系出数。针对YD_S和YD_X、XB_S和XB_X子体系,根据“2.3.2”分别核算森林生态系统服务功能物质量。
②相容机制。通过县对自身、市对县的小班核算体系(XB)结果是否落在样地核算体系(YD)置信区间内的双控制,来确认各级核算结果是否相容,降低核算误差。XB_X核算结果必须受YD_X核算置信区间控制[式(1)],满足该条件后,XB_S结果必须受YD_S的置信区间控制[式(2)],任意一项不满足,通过修正机制对XB_X核算结果进行修正。
$$ {U_{bj}} \in \left[ {{G_{bj}} - {A_b}{\mu _a}{S_{{{\overline y }_{bj}}}},{G_{bj}} + {A_b}{\mu _a}{S_{{{\overline y }_{bj}}}}} \right] \text{;} $$ (1) $$ \sum\limits_{b = 1}^5 {{U_{bj}}} \in \left[ {{G_j} - A{\mu _a}{S_{{{\overline y }_j}}},{G_j} + A{\mu _a}{S_{{{\overline y }_j}}}} \right] 。 $$ (2) 式(1)中:Ubj和Gbj分别为基于“二类”调查、“一类”清查数据的县级副总体b的第j类森林生态系统服务功能物质量;Ab为县级副总体b的土地总面积;μa为显著性水平a=0.05时的值,即1.96;
$S_{\overline y_{bj}} $ 为基于“一类”清查数据的县级副总体b的第j类森林生态系统服务功能物质量平均数标准误;Gj为基于“一类”清查数据的市级总体的第j类森林生态系统服务功能物质量;A为市级总体的土地总面积;$S_{\overline y_{j} }$ 为基于“一类”清查数据的市级总体的第j类森林生态系统服务功能物质量平均数标准误。③修正机制。第一,核算结果单位面积法修正。出现相容机制中所述情况的,则基于YD_S,采用[式(3)]对XB_X核算结果进行修正。
$$ {U_{bj({\rm{R}})}} = \displaystyle\sum\limits_{a = 1}^3 {\frac{{\displaystyle\sum\limits_{i = 1}^n {{y_{aij}}} }}{{\displaystyle\sum\limits_{i = 1}^n {{S_{ai}}} }}{S_{bai({\rm{XB}})}}} 。$$ (3) 式(3)中:Ubj(R)为修正后县级副总体b的第j类森林生态系统服务功能物质量;yaij为YD_S中,第a种森林类型(a=1为乔木林,a=2为竹林,a=3为灌木林)的第i个样地的第j类森林生态系统服务功能物质量;Sai为YD_S中,第a种森林类型的第i个样地面积;Sbaj(XB)为县级副总体b的第a种森林类型的第i个小班面积。
第二,产出核算结果。本研究所述的精度控制是指样地核算体系(YD)置信区间可以包含小班核算体系(XB)结果,小班核算结果即有精度保证。在此种情况下,产出基于小班核算体系(XB)的结果。因为在有精度保证的同时,小班核算结果可以落到山头地块,反之,则产出基于样地核算体系(YD)的结果。
-
由表3可见:县级副总体中,仅长兴县和安吉县的提供负离子物质量“二类”核算值未能落入置信区间;从估计精度看,安吉县和长兴县估计精度较高,南浔区估计精度最低。主要是961个样地基于市级总体森林资源空间变动系数和相对误差布设,县级副总体范围内的市级样地估计精度还受到县内森林破碎化程度带来的影响,南浔区地处杭嘉湖平原,森林空间分布零散,变异系数大。安吉县和长兴县以山地丘陵为主,森林面积占比较高,变异系数较小。
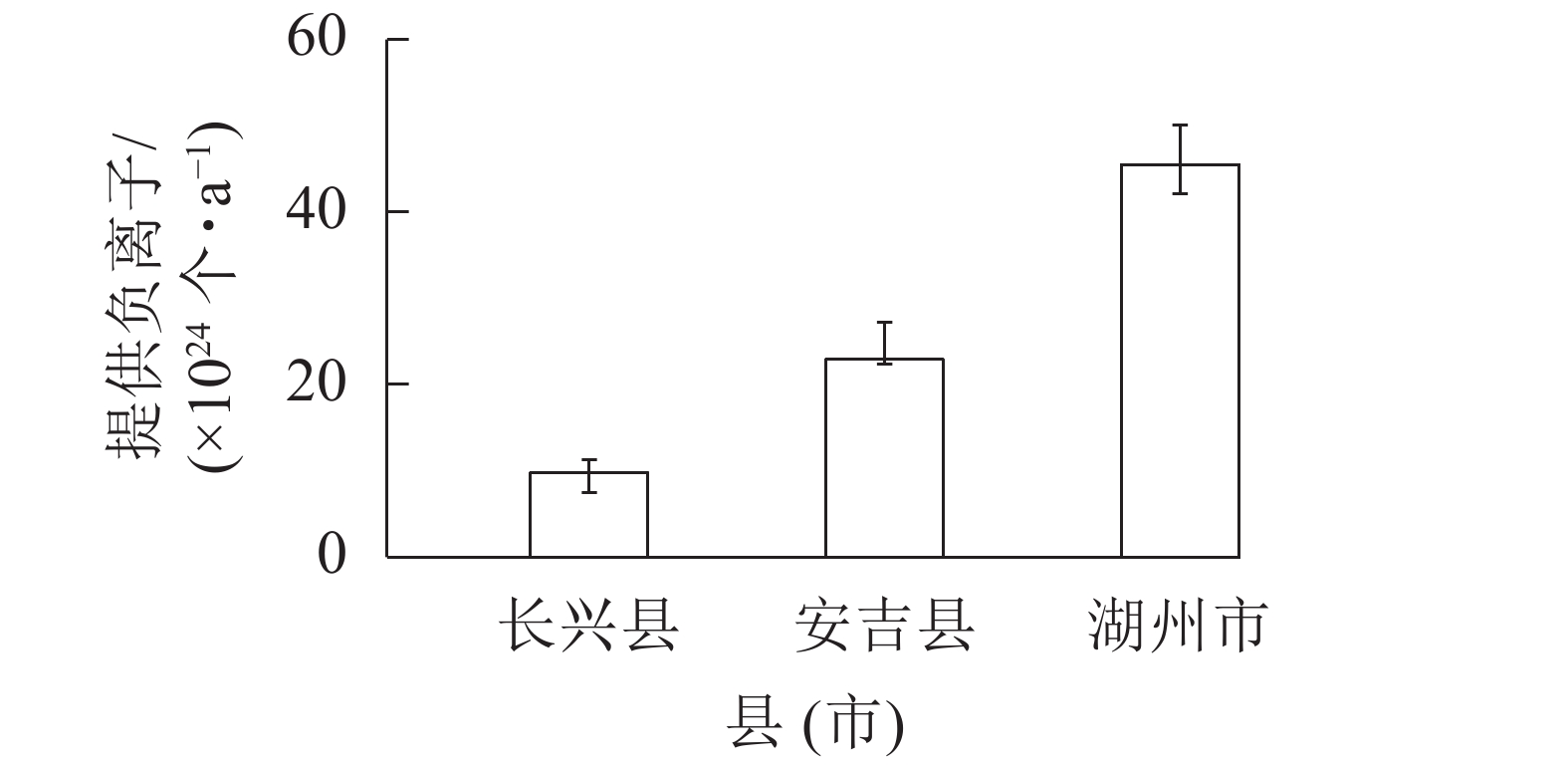
图 1 样地核算(a=0.05)森林提供负离子与修正后的县级副总体、市级总体结果比较
Figure 1. Comparion of CSLM and CLM volume of NAI between plots assessing results (a=0.05) and revised results
表 3 县级副总体森林生态系统服务功能物质量核算结果
Table 3. Accounting results of forest ecosystem services amount of CLSM
县级副总体 项目 水源涵养/
(×104 t·a−1)保育土壤/
(×104 t·a−1)固碳/
(×104 t·a−1)林木养分固持/
(×104 t·a−1)提供负离子/
(×1024个·a−1)吴兴区 估计中值 16 872.07 76.68 11.43 0.42 4.76 置信区间 13 852.14~19 892.00 63.34~90.03 7.78~15.08 0.29~0.55 3.30~6.23 估计精度/% 82.10 82.60 68.03 68.38 69.29 “二类”核算值 18 874.43 84.05 12.58 0.48 5.93 南浔区 估计中值 4 809.83 30.52 2.26 0.04 0.29 置信区间 3 466.21~6 153.45 22.01~39.04 1.33~3.18 0.02~0.06 0.14~0.45 估计精度/% 72.07 72.10 59.17 55.68 46.52 “二类”核算值 3 505.14 18.94 1.81 0.05 0.35 德清县 估计中值 20 895.11 100.76 16.59 0.56 6.93 置信区间 17 552.31~24 237.91 85.74~115.78 12.11~21.07 0.41~0.71 5.37~8.50 估计精度/% 84.00 85.10 73.01 73.08 77.44 “二类”核算值 23 458.00 100.25 17.42 0.54 6.42 长兴县 估计中值 39 992.91 195.04 28.07 1.09 9.40 置信区间 35 102.35~44 883.47 171.93~218.16 23.39~32.76 0.90~1.28 7.52~11.27 估计精度/% 87.77 88.15 83.30 82.77 80.04 “二类”核算值 39 843.56 172.78 27.98 1.15 6.85 安吉县 估计中值 76 780.87 357.00 58.43 2.06 24.79 置信区间 70 454.48~83 107.27 330.41~383.60 51.90~64.97 1.80~2.31 22.37~27.22 估计精度/% 91.76 92.55 88.81 87.71 90.23 “二类”核算值 79 087.77 345.53 52.62 1.82 19.49 -
各森林生态系统服务功能物质量的估计精度都超过90%,除提供负离子,其他核算结果均落入市级样地核算置信区间(表4)。水源涵养的市级总体小班汇总结果大于估计中值,比较2类调查所得的不同优势树种面积和蒸散率发现,“一类”清查数据的优势树种平均蒸散率为54.01%,“二类”调查为53.41%,而林分蒸散率越高,其水源涵养能力越低。保育土壤、固碳、林木养分固持的市级总体小班汇总结果均小于估计中值,主要原因是保育土壤物质量采用ULSE模型,由于“一类”清查得到的全市森林平均郁闭度(为0.71)比“二类”调查(为0.68)高,故所用的植被覆盖因子(C)更小。森林年蓄积量变化一定程度反映了森林固碳和林木养分固持的功能,“一类”清查得到全市森林单位面积蓄积69.04 m3·hm−2、蓄积年增率6.02%,均高于“二类”调查(63.45 m3·hm−2,5.95%)。平均树高是提供负离子物质量核算时的重要参数。“一类”清查得到的全市平均树高为8.5 m,相较高1.2倍,使得市级总体小班汇总结果未能落在置信区间内。
表 4 市级总体森林生态系统服务功能物质量核算结果
Table 4. Accounting results of forest ecosystem services amount of CLM
服务功能 水源涵养/
(×104 t·a−1)保育土壤/
(×104 t·a−1)固碳/
(×104 t·a−1)林木养分固持/
(×104 t·a−1)提供负离子/
(×1024个·a−1)估计中值 159 031.95 759.24 116.61 4.16 46.08 置信区间 148 728.91~169 334.98 712.62~805.86 106.16~127.06 3.77~4.55 42.06~50.10 估计精度/% 93.52 93.86 91.04 90.59 91.28 市级总体小班汇总结果 164 768.90 721.55 112.41 4.04 39.04 -
根据市县联动核算机制和式(3),对长兴县和安吉县的提供负离子核算结果进行修正(表5)。修正后,2个县提供负离子均落入YD_X的置信区间。市级总体森提供负离子汇总结果为45.46×1024个·a−1,也落入YD_S的置信区间(图1)。
表 5 县级副总体(长兴县和安吉县)森林提供负离子物质量核算结果修正
Table 5. Revision of assessing results of negative air ion supply of CLSM (Changxing County and Anji County)
项目 提供负离子/(×1024个·a−1) 样地(小班)面积/hm2 合计 乔木林 竹林 灌木林 乔木林 竹林 灌木林 市级样地 0.006 087 0.001 873 0.000 353 0.000 261 14.96 15.60 9.04 长兴县修正结果 9.80 5.04 4.47 0.29 40 277.67 17 629.53 9 984.00 安吉县修正结果 22.96 5.59 16.75 0.63 44 634.53 66 082.60 21 686.20 说明:修正结果为(各森林类型小班面积×对应森林类型市级样地的合计提供负离子物质量)/对应森林类型市级样地合计面积 -
本研究以湖州市为研究地,基于森林资源“一类”清查样地数据和“二类”调查小班数据,以样地和小班为纽带,构建了森林生态系统服务功能物质量市县联动核算体系。核算体系包括1个市级总体和5个县级副总体的核算布局;样地和小班2个核算体系;市县2级的同步核算机制,县对县、市对县核算结果双控制的相容机制,市对县核算结果修正的纠错机制3方面构成的联动核算机制。通过市县联动核算体系,为各级核算结果提供精度控制。从核算结果看,2018年,湖州市森林生态系统实现水源涵养量164 768.91万 t、保育土壤量721.54万 t、固碳量112.40万 t、林木养分固持量4.05万 t、提供负离子量45.46×1024个。从市级总体核算精度看,在显著性水平a=0.05下,森林生态系统服务功能物质量核算结果的估计精度均超过90%。
“一类”清查和“二类”调查属不同森林资源调查体系,故所得森林面积、林分测树因子不同,而森林生态系统服务功能量与森林面积、质量、气候环境有关[25],因此2类核算体系的核算结果也不一致。水源涵养、保育土壤、固碳和林木养分固持的小班汇总结果都能落入样地核算置信区间内,说明基于“二类”调查数据的核算结果是有精度保证的,其精度可直接采用估计精度数值。2类调查体系树高的调查差异使小班核算的提供负离子物质量不能落入样地核算置信区间,这主要是由于树高与空气负离子浓度差值有显著正相关[26]。不同县级副总体估计精度差异较大,地貌带来的森林破碎化程度是估计精度差异较大的客观原因,山地丘陵地区的土地利用类型以林地为主,变异系数较小,估计精度较高。而平原地区土地利用类型多样,森林资源的空间零散分布导致变异系数大,降低了估计精度。有研究性[19]表明:“一类”清查有精度保证,其抽样置信区间控制能够确保基于小班的森林资源结果可靠。本研究也证明了利用样地核算所得置信区间控制小班核算结果的方法可行性。本研究基于市级总体构建样地核算体系(YD),县级副总体的森林破碎化程度会对YD_X的估计精度带来影响。鉴于分层抽样[27]、不等概抽样[28]等能够提高森林资源监测结果精度,未来可探究上述抽样方案在森林生态系统服务功能核算时的精度控制效果。
综上所述,森林生态系统服务功能物质量市县联动核算体系能够为各级核算结果提供精度控制。小班核算体系(XB)结果落入样地核算体系(YD)置信区间,可使用样地核算体系(YD)估计精度,并采用前者核算结果确认区域森林生态系统服务功能物质量。反之,对小班核算结果进行修正,修正后仍不能落入置信区间的,应按样地核算结果进行确认。
City-county synchronized assessing and accuracy control of forest ecosystem services
-
摘要:
目的 探讨以市县联动核算体系为手段进行森林生态系统服务功能物质量精度控制的应用方法。 方法 选取浙江省湖州市为研究地,通过设定1个市级总体和5个县级副总体,并依托森林资源连续清查和森林资源规划设计调查体系建立样地核算和小班核算2个体系,形成由同步核算机制、相容机制和修正机制构成的市县联动核算机制,完成森林生态系统水源涵养、土壤保育、固碳、林木养分固持和提供负离子5项指标的物质量核算和精度控制。 结果 在显著性水平a=0.05下,市级总体各指标物质量的样地核算结果估计精度均超过90%;除提供负离子,其他指标物质量的小班核算结果均落入样地核算置信区间。县级副总体中,除长兴县和安吉县的提供负离子,其他指标的小班核算结果均落入样地核算置信区间。采用修正机制,对提供负离子结果予以修正,最后得到2018年湖州市森林生态系统5项服务功能物质量与精度。 结论 市县联动核算体系能够对各层级森林生态系统功能物质量核算结果进行精度控制,可根据小班核算结果是否落入样地核算置信区间来确定区域森林生态系统服务功能物质量最终结果。图1表5参28 Abstract:Objective This study is designed to explore the application method of accuracy control of volume of forest ecosystem services (FES) by means of city-county synchronized assessing system. Method Huzhou City of Zhejiang Province was selected as the research site. By setting up 1 city-level module (CLM) and 5 county-level sub-modules (CLSM), 2 systems of plot data assessing (PDA) and sub-compartment data assessing (SDA) were established based on the continuous forest inventory and forest management inventory to form a city-county synchronized assessing mechanism composed of synchronous accessing mechanism, compatibility mechanism and correction mechanism, so as to complete the volume calculation and assessing accuracy control of multiple FES indicators (water conservation, soil conservation, carbon sequestration, nutrient retention and negative ions provision). Result Under the significance level a=0.05, the accuracy of PDA results of each FES indicators at the city level exceeded 90%. Except for negative ions provision, SDA results of other indicators fell into PDA confidence interval. In county-level sub-modules (a=0.05), except for the negative ions provision in Changxing County and Anji County, SDA results of all other indicators fell into PDA confidence interval. Using the correction mechanism, the results of negative ions provision were corrected, and finally the volume and assessing accuracy of 5 FES in Huzhou City in 2018 were obtained. Conclusion The city-county synchronized assessing system can control the accuracy of the quality accounting results of FES at all levels, and the final results of regional FES quality can be determined according to whether SDA results fall into PDA confidence interval. [Ch, 1 fig.5 tab. 28 ref.] -
棉纤维是由外珠被表皮层的单细胞分化发育而成,分为长绒(lint)和短绒(fuzz)2种,长绒是高级棉纱纺织品的主要原材料,短绒主要用做制作纤维素、絮棉、纸张及纺织品的原料。在已有的四倍体棉种中,陆地棉Gossypium hirsutum和海岛棉Gossypium barbadense已经被驯化为栽培种[1-3]。目前世界上97%的棉纤维都产自陆地棉,产量高且适应性广,但是纤维品质中等;海岛棉产量低,适应性差,栽培范围不广泛,但是其纤维更长且品质高。如何获得优质高产的棉种,一直是遗传育种学家关注的焦点。而随着遗传学、细胞学和分子生物学等学科的交叉融合,棉纤维生长发育分子机制已成为国内外研究的热点。探明棉花种子表皮细胞生长发育的分子基础,对于提高棉花产量及改良纤维品质至关重要。早期有关棉纤维发育研究大多集中于遗传定位。大量与纤维品质和产量相关的数量性状位点(QTL)通过图位克隆的方法被发现于各个染色体[4-5],而光子显性基因Li1,Li2,N1和Fbl以及光子隐性基因n2,sma-4(fz)和sma-4(ha)[6]等一直备受关注。近年来,深度测序技术的兴起,对棉纤维发育的分子机制的研究起了有效的推动作用。随着深度测序技术的不断革新,棉花全基因组测序不断完善[7],全基因组微卫星序列得以注释[8],单核苷酸多态性(SNP)芯片的开发成为可能[9],使得遗传定位工作更加便捷[9-10]。转录组学、蛋白组学及表观遗传学领域三方位的深度测序有效构建了核糖核酸(RNA)水平和蛋白质水平、编码区域和非编码序列之间的联系,并发现一系列的转录因子、编码转脂蛋白的基因、钙信号转导相关基因、多糖合成相关蛋白、大量的微核糖核酸(miRNA)以及脱氧核糖核酸(DNA)甲基化作用等共同参与棉纤维发育过程[11-16]。本文将从棉纤维发育各时期的形态结构变化及特征,经典遗传学研究,深度测序技术在转录组学、蛋白组学及表观遗传学领域的运用,以及棉纤维发育各个时期所涉及的相关调控基因等4个方面对棉纤维发育机制的研究进展进行综述。
1. 棉纤维发育各时期的形态结构变化特征
棉纤维细胞发育进程是一个多基因调控的有序的系统发生过程,整个细胞分化过程可被分为棉纤维起始、伸长、次生壁合成与增厚、脱水成熟等4个时期[17-20]。
1.1 棉纤维起始期
长绒纤维细胞一般在开花前或开花当天就开始突起,而短绒纤维的突起要稍迟几天,两者的分化过程基本相似[21]。RAMSEY等[22]通过观察开花前16 d到开花当天胚珠的亚显微结构,发现开花前16 d到前3 d表皮细胞无差异,说明纤维原始细胞的分化与突起晚于开花前3 d。而在开花前2~3 d纤维原始细胞受生长素(IAA)和赤霉素(GA3)的刺激开始产生纤维[11],开花当天,纤维原始细胞的分化与突起已基本完成。棉花纤维原始细胞分化与突起多少决定种子表面纤维数量,从而决定了棉纤维的产量。
1.2 棉纤维伸长期
研究发现一般只有25%~30%的棉花种子表皮细胞(约2万个)能正常突起伸长,形成成熟的纤维[23-24]。棉纤维的伸长几乎和突起同时进行,从开花当天开始,发生在细胞壁膨胀过程,纤维的最终长度取决于纤维伸长速率和持续时间2个方面[25],一般持续20~30 d,该过程通过一种扩散生长机制实现并指导纤维细胞的极化生长[26-27]。纤维伸长分为非极性膨胀和极性伸长2个阶段:非极性膨胀期决定了纤维的细度,纤维细胞向四周扩展直至形成纤维的最终直径[28];极性伸长期可使纤维长度达到最终长度的80%[29],这一时期生化反应最为活跃,主要决定纤维的长度,是影响纤维品质的关键时期[24]。
1.3 棉纤维次生壁合成与增厚期
棉纤维次生壁合成与增厚期和纤维伸长期存在一段时期的重叠[17],在开花后16 d开始,持续到开花后40 d,在这段时期,纤维素大量沉积,次生壁不断加厚[30]。伴随着纤维素沉积的加速,纤维伸长逐渐减弱,该过程是影响纤维强度和韧性的关键时期。
1.4 棉纤维脱水成熟期
在次生壁合成与增厚期后就进入了纤维脱水成熟期,发生在开花后40~50 d,棉铃开裂至充分吐絮,纤维失水,形成转曲[31]。成熟的棉纤维由外向内依次为初生壁、次生壁和中腔。
2. 棉花纤维生长发育的遗传学研究
遗传规律研究和基因遗传定位是经典遗传学中2项重要的基础工作。在棉纤维遗传规律研究中发现,相同性状的材料基因型不同,其遗传模式也不同,而棉纤维发育相关基因的遗传定位又可被分成质量性状和数量性状(QTL)的定位。
2.1 棉纤维遗传规律研究
CARVER[32]和KEARNEY等[33-34]研究发现棉花光子性状主要由2对独立的位点控制,显性光子基因(N1)和隐性光子基因(n2),宋丽等[35]证实这2种光子基因均符合单基因遗传模型。既无长绒也无短绒的L40突变体的光子性状为不完全显性[36]。既无长绒也无短绒的突变体Xu142 fl的短绒的发育受N1和n2 2对基因控制,长绒的发育受Li3基因位点控制[37]。陆地棉短绒突变体Li1和Li2均为单基因显性遗传[38-40]。孙亚莉等[41]选取大量的陆地棉和海岛棉的光子材料对棉花光子性状进行了遗传分析,其研究发现棉花短绒多少与生态环境有关系,且不同品种光子材料的遗传模式也不同,不论海岛棉还是陆地棉材料均存在显性、部分显性和隐性遗传。对3个陆地棉隐性性状的材料进一步研究表明:这3个材料的遗传规律均不同,‘库光子’的光子性状由2对隐性等位基因控制,并且有互补效应;‘陆无絮’的光子性状由2对隐性等位基因控制,基因间呈积加作用;SA65的光子性状由单隐性基因控制。
2.2 棉纤维基因的QTL定位
纤维品质性状包括长度、整齐度、伸长率、强度、细度、颜色和马克隆值等多个方面。随着分子标记的不断开发与应用,在棉花染色体A组和D组染色体上都有大量棉纤维品质和产量相关的QTL被发现(表 1)。从表 1可知:纤维品质和产量性状的QTL几乎遍布了每一条染色体,且不同实验室使用不同的群体所得到的结果也有很大差异。同时,研究也发现这些性状受环境的影响很大,某些QTL在不同环境条件下有变化,甚至检测不到,导致已定位的QTL间重复性差[10, 42],这也说明纤维品质及产量性状的遗传非常复杂。研究也发现了一些稳定的主效QTL,如第10号染色体的棉纤维强度主效QTL(FS1),解释了超过30.00%的表型变异[43];第19号染色体影响衣分的QTL(qLI17),解释24.30%的表型变异[34];第8号染色体上颜色相关QTL(Ge6_Rd_8_3_10.60_[+]),解释48.00%的表型变异[5];以及第14号染色体上与长度相关的QTL(qFL-Chr14-3),解释15.05%的表型变异[10],等等。此外,有些QTL虽然微效,但在不同环境下都能稳定存在,比如WANG等[42]在8,11,12和21号染色上发现的6个QTL:qFL-A8-1(长度相关),qFS-A8-1(强度相关),qFS-A12-1(强度相关),qFS-A12-2(强度相关),qFS-D11-1(强度相关)和qFM-A11-1(马克隆值相关)。这些稳定存在的QTL都值得科研工作者进一步关注和研究。
表 1 不同群体中与棉纤维品质和产量相关的QTL分布Table 1. QTL related to cotton fiber quality and yield in different populations性状 QTL所在染色体或连锁群 检出限(LOD) 变异率1% 群体 出处 长度 Chr04,Chrl8,Chr22 2.00~2.74 7.80~12.60 陆地棉TM-1×海岛棉3-79的F2群体 KOHEL等[44] Chr20,LGA02(Chr08),LGA03 (Chr11),LGA05 2.63~5.40 2.90~13.70 陆地棉Siv’ on×海岛棉F-177的F2和F3群体 PATERSON等[4] Chr04 3.50 24.00 陆地棉Acala 44×海岛棉Pima S-7的F2群体 MEI等[45] Chr01,Chr03,Chr04,Chr06,Chr09,Chr13,Chr14,Chr18,Chr19,Chr20,Chr21,Chr23,Chr24,Chr26 3.30~9.50 6.00~40.00 陆地棉Guazuncho-2×海岛棉VH8-4602RIL LACAPE等[5] Chr05,Chr07,Chr08,Chr11,Chr12,Chr19,Chr21,Chr23,Chr26 4.57~6.05 2.47~8.49 陆地棉TM-1×海岛棉Hai7124的CSILs群体 WANG等[42] Chr10,Chr14,Chr15 2.50~7.71 6.21~15.05 陆地棉HS46 ×陆地棉MAR CABU-CAG8US-1-88 RIL LI等[10] 整齐度 Chr04,Chr14,Chr15,Chr22,LGA03 (Chr11),LGA05 1.65~3.79 2.10~13.30 陆地棉Siv’on×海岛棉F-177的F2和F3群0体 PATERSON等[4] Chr05,Chr09,Chr12,Chr15,Chr16,Chr18,Chr19,Chr20,Chr23,Chr26 3.50~7.80 9.00~32.00 陆地棉Guazuncho-2×海岛棉VH8-4602 RIL LACAPE等[5] Chr09 2.68~4.17 5.58~10.94 陆地棉HS46 ×陆地棉MARCABU- CAG8US-1-88的RIL LI等[10] 伸长率 Chr05,Chr10,Chr15,Chr23,LGA02 (Chr8),LGA03(Chr11),LGD07 2.32~5.77 3.40~8.90 陆地棉Siv’on×海岛棉F-l77的F2和F3群体 PATERSON等[4] Chr09 5.16 42.00 陆地棉Acala 44×海岛棉Pima S-7的F2群体 MEI等[45] Chr02,Chr06,Chr09,Chr10,Chr12,Chr13,Chr15,Chr19,Chr20,Chr21,Chr23,Chr26 3.40~6.70 6.00~21.00 陆地棉Guazuncho-2×海岛棉VH8-4602RIL LACAPE等[5] Chr14,Chr20,Chr24 2.49~7.80 5.35~32.28 陆地棉HS46 ×陆地棉MARCABU-CAG8US-1-88 RIL LI等[10] 强度 Chr03,Chr14,Chr15,Chr25 2.08~2.69 10.40~23.10 陆地棉TM-1×海岛棉3-79的F2群体 KOHEL等[10] Chr10 4.79~5.80 53.00~53.80 异质棉7235×陆地棉TM-1的F2群体 ZHANG等[43] Chr01,Chr04,Chr14,Chr17,Chr18,Chr20,Chr22,Chr23,Chr25,LGA01 (Chr13),LGA02(Chr08),LGA03(Chr11),LGA05,LGD02(Chr21),LGD03(Chr24),LGD04,LGD07 0.21~6.22 2.50~17.40 陆地棉Siv’on×海岛棉F-177的F2和F3群体 PATERSON等[4] Chr03,Chr04,Chr05,Chr07,Chr09,Chr12,Chr14,Chr15,Chr16,Chr18,Chr19,Chr21,Chr23,Chr26 3.30~8.50 7.00~31.00 陆地棉Guazuncho-2×海岛棉VH8-4602RIL LACAPE等[5] Chr05,Chr07,Chr08,Chr09,Chr11,Chr12,Chr13,Chr14,Chr15,Chr16,Chr17,Chr18,Chr21,Chr23 7.32~22.54 5.07~15.82 陆地棉TM-1×海岛棉Hai7124的CSILs群体 WANG等[42] 细度 Chr01,Chr02,Chr03,Chr12,Chr16,LGD01 2.16~4.04 16.70~43.90 陆地棉TM-1×海岛棉3-79的F2群体 KOHEL等[44] Chr02,Chr04,Chr05,Chr06,Chr09,Chr14,Chr15,Chr17,Chr20,Chr23,Chr25,LGA01 (Chr13),LGA05,LGA06,LGD01,LGD02(Chr2l),LGD03(Chr24),LGD04,LGD05,LGD07 2.21~9.78 2.20~30.30 陆地棉Siv’on×海岛棉F-l77的F2和F3群体 PATERSON等[4] Not determined 5.11 43.20 陆地棉Acala 44×海岛棉Pima S-7的F2群体 MEI等[45] Chr01,Chr02,Chr03,Chr04,Chr05,Chr06,Chr08,Chr09,Chr10,Chr12,Chr15,Chr16,Chr17,Chr18,Chr19,Chr20,Chr21,Chr22,Chr23,Chr24,Chr25,Chr26 3.30~8.90 6.00~41.00 陆地棉Guazuncho-2×海岛棉VH8-4602RIL LACAPE等[5] 颜色 Chr06,Chr09,Chr14,Chr17,Chr18,Chr22,Chr25,LGA01,LGA02,LGA03,LGD02(Chr21) 2.66~11.67 2.50~14.90 陆地棉Siv’on×海岛棉F-177的F2和F3群体 PATERSON等[4] Chr01,Chr02,Chr06,Chr07,Chr08,Chr09,Chr11,Chr14,Chr15,Chr17,Chr18,Chr19,Chr21,Chr22,Chr25 3.30~10.60 6.00~48.00 陆地棉Guazuncho-2×海岛棉VH8-4602RIL LACAPE等[5] 马克隆值 Chr05,Chr06,Chr09,Chr11,Chr12,Chr15,Chr16,Chr19,Chr21,Chr22 4.56~9.09 0.80~8.03 陆地棉TM-1×海岛棉Hai7124的CSILs群体 WANG等[42] Chr14,Chr16 2.51~4.23 5.52~9.20 陆地棉HS46 ×陆地棉MARCABU- CAG8US-1-88 RIL LI等[10] 产量 A02(Chr08),A03(Chr11),Chr14,Chr23,Chr25,LG5 3.00~5.28 13.01~28.35 陆地棉Handan 208×海岛棉Pima 90的F2群体 HE等[46] 衣分 D08(Chr19) 3.45 24.34 陆地棉Handan 208×海岛棉Pima 90的F2群体 HE等[46] 然而,棉纤维相关性状QTL的分离和克隆仍然很少。有研究发现在第12条染色体上(A12/D12)与棉纤维品质相关的QTL附近的GhHOX3基因对棉纤维长度起重要调控作用[47],以及定位于同源染色体A8(chr08)和D8(chr24)上的GhSusA1基因过表达可以增强纤维长度和强度[48]。
2.3 棉纤维基因的质量性状定位
在光子显性基因定位中发现,N1和Fbl基因位于chr12上[6],其中N1基因被鉴定为转录因子MYB25-like[49]。光子隐性基因定位研究表明:n2定位在chr26上,sma-4(fz)位于L.G.A3的端部,sma-4(ha)位于L.G.A3中部[6]。超短纤维突变体Li1基因位于chr22上,并已通过精细定位被克隆到,是一个肌动蛋白家族基因[50-53];而Li2基因则位于chr18上[6, 53]。
3. 棉纤维发育组学研究
棉纤维发育过程涉及到大量的基因和通路调控。利用高通量测序技术,对棉纤维发育的转录组和蛋白组分析及表观遗传研究,可以短时间内获得大量信息,捕获到许多参与不同发育阶段的特异性基因及信号通路,为下游单独研究重要基因的功能奠定良好的基础。
3.1 转录组学研究
利用棉花胚珠体外培养技术结合转录组数据分析比较,KIM等[11]在纤维起始分化时期发现了许多在野生型和无毛突变体间表达有差异的基因,包括MYB25,MYB109,PDF1,MYB25-like,HD1等转录调节因子,表明在棉纤维起始分化过程中存在复杂的信号网络调节机制。通过比较短绒突变体(Li1)与野生型之间在开花后1,3和8 d的转录组数据,LIANG等[49]在胚珠中共检测到7 852个差异表达基因,主要参与次生代谢物和脂质代谢途径,其中涉及非长链脂肪酸生物合成的37个基因在Li1突变体纤维的快速伸长发育过程中被显著抑制,这说明脂质代谢途径与纤维伸长密切相关。HOVAV等[54]评估了从开花后初级次生壁到次级次生壁合成过程中棉纤维发育的转录组变化发现,棉纤维发育过程中的基因转录水平很高,在每个阶段占到所有基因的75%~94%,并且半数以上的基因在纤维发育的至少一个阶段中上调。BOLTON等[55]利用基因芯片技术和实时荧光定量PCR技术在Li1突变体中发现超过100个基因在次生壁的生物合成过程中差异表达,其中的3个候选基因:伸展蛋白(extensin),蔗糖合成酶(sucrose synthase)和微管蛋白(actin)的表达量明显偏离野生型的表达水平。通过陆地棉TM-1背景下的海岛棉染色体导入系与亲本纤维的转录组差异比较,FANG等[56]在CSIL-35431和CSIL-31010等2个导入系的次生壁合成过程中发现了大量与TM-1有表达差异的基因,功能富集分析表明这些基因主要富集于次生细胞壁的生物合成、葡糖醛酸合成、纤维素合成等生物途径。
3.2 蛋白质组学研究
利用蛋白组学研究,许多棉纤维发育过程中的重要蛋白被不断发掘,且此技术很好地互补了转录组只能在mRNA水平上研究纤维相关基因的劣势。HU等[12]应用相对和绝对定量(iTRAQ)LC-MS/MS分析技术研究了1 317个纤维特异性表达蛋白,其中205个蛋白在发育阶段中差异表达,190个蛋白在野生和栽培棉之间差异表达。结合转录组、iTRAQ蛋白质组和遗传图谱定位的综合分析方法,MA等[13]发现徐州142野生型与其无绒毛突变体(fl)的胚珠之间存在大量差异表达的基因和蛋白,这些差异基因和蛋白主要存在于氨基酸、核苷酸、脂肪酸和叶酸代谢以及黄酮生物合成中,说明这些代谢途径在纤维发育过程中具有重要作用。
3.3 表观遗传学研究
近年来,棉纤维发育的表观遗传学研究也取得了巨大进展。基于pre-miRNAs和已发现的miRNA靶基因数据,CHEN等[14]对83个miRNA前体及其目标调控基因进行了研究,并构建了miRNAs及其靶位点调控网络,并揭示了这些miRNA及靶基因在纤维不同发育阶段的表达模式。SONG等[15]对纤维和胚珠进行了甲基化组、转录组和小RNA组学分析,发现在胚珠和纤维发育过程中CHH甲基化变化显著。该研究发现,在胚珠中,启动子中的CHH甲基化与可诱导RNA依赖的DNA甲基化(RdDM)和胚珠偏好基因上调的siRNA呈正相关;在纤维细胞中,胚珠衍生细胞产生独立的RdDM的异染色质CHH超甲基化,抑制转座子及附近纤维相关基因的活性。使用甲基化抑制剂5-氮-2′-脱氧胞苷对胚珠进行体外培养,可以减少纤维细胞的数量和长度,这表明DNA甲基化在纤维发育中具有潜在作用。这些研究表明:启动子和转座子及附近基因中的RdDM依赖的甲基化可作为基因和转座子表达的双保险反馈机制。ZOU等[16]对参与纤维起始和伸长过程的长链非编码RNA(lncRNA)进行了系统分析,共鉴定到5 996条lncRNAs,其中长链非编码RNA(lincRNA)3 510条,天然反义转录RNA(lncNAT)2 486条,表明lncRNA对棉纤维的发育至关重要。
4. 棉纤维发育不同时期相关调控基因研究
4.1 棉纤维起始期相关基因
在棉花胚珠EST数据库中,约10%的基因与转录因子密切相关,包括56个转录因子家族成员[57],如MYB类转录因子家族的GL1及其同源基因MYB2[58],MYB109[59],TTG1[60]和GL2[61]等均被发现在棉纤维发育的早期阶段高效表达。棉花GaMYB2基因可以诱导种子表皮毛的产生,且转化拟南芥可以弥补GL1突变对表皮毛起始分化造成的影响[58]。通过干扰GhMYB109的表达,PU等[62]发现棉纤维细胞分化延迟且起始数量减少,说明GhMYB109在棉花纤维分化阶段起重要作用。LOGUERICO等[63]发现GhMYB4和GhMYB5基因在纤维分化期的胚珠中特异表达。WALFORD等[64]发现GhHD1可以介导棉花表皮细胞的分化。辛婧[65]的研究表明转录因子GbSPB8可能调控棉花纤维起始发育。
4.2 棉纤维伸长期相关基因
转脂蛋白和钙信号转导相关蛋白在棉纤维伸长过程中起到极其重要的作用。MA等[66]研究发现:转脂蛋白GhLTP3和GhLTP6的表达量在纤维快速伸长期达到最高水平,此外李锡花等[67]发现GhLTP3的表达量从开花后0~15 d中表达量不断升高,在第15天达到顶峰,之后逐渐下降。赵存鹏等[68]和GAPPER等[69]研究发现:钙调蛋白CaM在低温逆境的条件下能使活性氧(ROS)、超氧游离基、过氧化氢和羟自由基等物质提高,进而使纤维细胞壁松弛,从而影响细胞伸长。CHENG等[70]发现GhCaM7-like基因在纤维快速伸长期显著表达。HUANG等[71]发现钙依赖性蛋白激酶GhCPK1在开花第10天的胚珠中高表达。这些研究都说明钙信号转导在纤维伸长过程中发挥重要作用。除了转脂蛋白和钙信号转导相关蛋白,转录调节因子也参与其中,如ZHANG等[72]发现GbMYB25与GbML1相互作用并通过调节ROS信号调节纤维伸长。HSU等[73]发现GhMYB7可以调控LTP3等脂转移蛋白编码基因。
4.3 棉纤维次生壁合成与加厚期相关基因
DELMER等[74]发现Rac9和Rac13可以控制棉花纤维素沉积方向。ZHAO等[75]发现GhRGP1在纤维发育的初生壁伸长及次生壁加厚后期优势表达,参与植物细胞壁非纤维素类的多糖合成。杨郁文等[76]发现一种Ser/Thr激酶和Try激酶的双受体蛋白GhRLK1与激活和维持次级细胞壁形成的细胞信号传导过程有关。此外,GhRDL1(RD22-Like1)与GhEXPA1互作会影响纤维细胞壁发育,GhRDL1基因过表达会产生长且质量较好的棉纤维[77]。
4.4 棉纤维脱水成熟期相关基因
目前,关于棉纤维细胞脱水成熟期的相关研究较少,对于纤维在脱水成熟过程中细胞与分子水平的变化还不清楚,只是猜想可能涉及到棉纤维细胞的程序性死亡[78]。
5. 研究展望
随着技术的革新,大量棉纤维发育相关基因和涉及的调控网络被不断发现,但是基因间的相互作用及其潜在的调控机制还有待进一步探索。近年来,棉花转基因技术日益成熟,新兴的CRISPR/Cas9基因编辑系统也于2017年3月在棉花基因组靶基因敲除中得到首次应用。JANGA等[79]利用CRISPR/Cas9系统成功将转绿色荧光蛋白(GFP)基因棉花系的GFP基因敲除;LI等[80]以棉花內源GhMYB25为目标基因,使用2种单导向RNA对该基因进行定点突变,突变率分别为100%和98.8%。这些研究表明:CRISPR/Cas9可以在棉花基因组上进行高效和高特异性地突变。随着新技术在棉花中的应用与成熟,棉纤维发育相关基因及其调控机制也有望有更多的发现。
-
图 1 样地核算(a=0.05)森林提供负离子与修正后的县级副总体、市级总体结果比较
Figure 1 Comparion of CSLM and CLM volume of NAI between plots assessing results (a=0.05) and revised results
表 1 湖州市2018年森林面积、蓄积量及其占比
Table 1. Forest areas, inventory and percentages of Huzhou City in 2018
县(区) 森林面
积/km2森林面积
占比/%森林蓄积量/
(×104 m3)森林蓄积量
占比/%吴兴区 329.4 11.7 127.4 16.8 南浔区 69.8 2.5 8.0 1.1 德清县 400.0 14.3 77.7 10.2 长兴县 678.9 24.2 212.7 28.0 安吉县 1 324.0 47.3 333.3 43.9 总计 2 802.1 100.0 759.1 100.0  下载: 导出CSV
下载: 导出CSV
表 2 森林生态系统服务功能指标体系及本研究指标选取说明
Table 2. Index system of forest ecosystem services function and explanation of index selection
服务类别 功能类别 指标类别 说明 供给服务 生物多样性 物种资源保育 主要通过野生动植物资源调查成果核算,故未纳入 林木产品供给 木材产品 此2项指标的产量一般是统计数据,与本研究目的不一致,故未纳入 非木材产品 调节服务 水源涵养 调节水量 此2项指标物质量核算方法与结果均一致,仅在评估价值量时,分别选取
不同单价。在实际应用中,水源涵养物质量也仅指调节水量。因此,本
研究选择调节水量一项指标,且后文中的水源涵养表述仅指代调节水量净化水质 固碳释氧 固碳 此2项指标核算方法差异仅体现在转换参数不同,而且,释氧量可通过固
碳量直接乘以氧气(O2)与碳(C)的相对原子质量比得到。因此,本研究
仅选择固碳一项指标释氧 净化大气环境 提供负离子 由于森林单位面积吸收气体污染物及滞纳TSP等的实测参数不足,本研究
仅选择提供负离子一项指标吸收气体污染物 滞纳TSP、PM10、PM2.5 森林防护 防风固沙 研究区不属于风力侵蚀区域。农田防护功能仅针对农田境界外100 m范围
内的森林。且2类调查数据未能界定森林是否为农田防护林,因此未选
取该2项指标农田防护 文化服务 森林康养 森林康养 森林康养价值一般是统计数据,或通过条件价值法等方法核算。与本研
究目的不一致,未选取该指标支持服务 保育土壤 固土 考虑到保肥量仅是固土量乘以土壤营养元素百分比含量(为常数)得到,固
土量核算精度决定了保肥量核算精度。因此,本研究仅选择固土指标。
且后文中的保育土壤表述仅指代固土保肥 林木养分固持 氮、磷、钾固持 该指标反映了林木吸收氮、磷、钾等营养元素并储存在体内的功能,因
此,本研究选择此3项指标,并以此3项指标物质量之和作为林木养分
固持物质量
下载: 导出CSV
表 3 县级副总体森林生态系统服务功能物质量核算结果
Table 3. Accounting results of forest ecosystem services amount of CLSM
县级副总体 项目 水源涵养/
(×104 t·a−1)保育土壤/
(×104 t·a−1)固碳/
(×104 t·a−1)林木养分固持/
(×104 t·a−1)提供负离子/
(×1024个·a−1)吴兴区 估计中值 16 872.07 76.68 11.43 0.42 4.76 置信区间 13 852.14~19 892.00 63.34~90.03 7.78~15.08 0.29~0.55 3.30~6.23 估计精度/% 82.10 82.60 68.03 68.38 69.29 “二类”核算值 18 874.43 84.05 12.58 0.48 5.93 南浔区 估计中值 4 809.83 30.52 2.26 0.04 0.29 置信区间 3 466.21~6 153.45 22.01~39.04 1.33~3.18 0.02~0.06 0.14~0.45 估计精度/% 72.07 72.10 59.17 55.68 46.52 “二类”核算值 3 505.14 18.94 1.81 0.05 0.35 德清县 估计中值 20 895.11 100.76 16.59 0.56 6.93 置信区间 17 552.31~24 237.91 85.74~115.78 12.11~21.07 0.41~0.71 5.37~8.50 估计精度/% 84.00 85.10 73.01 73.08 77.44 “二类”核算值 23 458.00 100.25 17.42 0.54 6.42 长兴县 估计中值 39 992.91 195.04 28.07 1.09 9.40 置信区间 35 102.35~44 883.47 171.93~218.16 23.39~32.76 0.90~1.28 7.52~11.27 估计精度/% 87.77 88.15 83.30 82.77 80.04 “二类”核算值 39 843.56 172.78 27.98 1.15 6.85 安吉县 估计中值 76 780.87 357.00 58.43 2.06 24.79 置信区间 70 454.48~83 107.27 330.41~383.60 51.90~64.97 1.80~2.31 22.37~27.22 估计精度/% 91.76 92.55 88.81 87.71 90.23 “二类”核算值 79 087.77 345.53 52.62 1.82 19.49
下载: 导出CSV
表 4 市级总体森林生态系统服务功能物质量核算结果
Table 4. Accounting results of forest ecosystem services amount of CLM
服务功能 水源涵养/
(×104 t·a−1)保育土壤/
(×104 t·a−1)固碳/
(×104 t·a−1)林木养分固持/
(×104 t·a−1)提供负离子/
(×1024个·a−1)估计中值 159 031.95 759.24 116.61 4.16 46.08 置信区间 148 728.91~169 334.98 712.62~805.86 106.16~127.06 3.77~4.55 42.06~50.10 估计精度/% 93.52 93.86 91.04 90.59 91.28 市级总体小班汇总结果 164 768.90 721.55 112.41 4.04 39.04
下载: 导出CSV
表 5 县级副总体(长兴县和安吉县)森林提供负离子物质量核算结果修正
Table 5. Revision of assessing results of negative air ion supply of CLSM (Changxing County and Anji County)
项目 提供负离子/(×1024个·a−1) 样地(小班)面积/hm2 合计 乔木林 竹林 灌木林 乔木林 竹林 灌木林 市级样地 0.006 087 0.001 873 0.000 353 0.000 261 14.96 15.60 9.04 长兴县修正结果 9.80 5.04 4.47 0.29 40 277.67 17 629.53 9 984.00 安吉县修正结果 22.96 5.59 16.75 0.63 44 634.53 66 082.60 21 686.20 说明:修正结果为(各森林类型小班面积×对应森林类型市级样地的合计提供负离子物质量)/对应森林类型市级样地合计面积
下载: 导出CSV
-
[1] 蒋有绪. 世界森林生态系统结构与功能的研究综述[J]. 林业科学研究, 1995, 8(3): 314 − 320. JIANG Youxu. A review of the structure and function of the world’s forest ecosystems [J]. For Res, 1995, 8(3): 314 − 320. [2] 靳芳, 鲁绍伟, 余新晓, 等. 中国森林生态系统服务功能及其价值评价[J]. 应用生态学报, 2005, 16(8): 1531 − 1536. JIN Fang, LU Shaowei, YU Xinxiao, et al. Evaluation of forest ecosystem service functions and their values in China [J]. Chin J Appl Ecol, 2005, 16(8): 1531 − 1536. [3] GYMER R. Man’s Impact on the Global Environment: Assessment and Recommendation for Action[M]. Massachusetts: The MIT Press, 1970. [4] DAILY G C. Nature’s Services: Societal Dependence on Natural Ecosystems[M]. Washington DC: Island Press, 1997. [5] COSTANZA R, D’ARGE R, DE GROOT R, et al. The value of the world’s ecosystem services and natural capital [J]. Nature, 1997, 387(6630): 253 − 260. [6] FISHER B, TURNER R, ZYLSTRA M, et al. Ecosystem services and economic theory: integration for policy research [J]. Ecol Appl, 2008(18): 2050 − 2067. [7] 赵金龙, 王泺鑫, 韩海荣, 等. 森林生态系统服务功能价值评估研究进展与趋势[J]. 生态学杂志, 2013, 32(8): 2229 − 2237. ZHAO Jinlong, WANG Luoxin, HAN Hairong, et al. Research advances and trends in forest ecosystem services value evaluation [J]. Chin J Ecol, 2013, 32(8): 2229 − 2237. [8] 王兵. 森林生态连清技术体系构建与应用[J]. 北京林业大学学报, 2015, 37(1): 1 − 8. WANG Bing. Construction and utilization of continuous inventory system for forest ecology [J]. J Beijing For Univ, 2015, 37(1): 1 − 8. [9] 欧阳志云, 朱春全, 杨广斌, 等. 生态系统生产总值核算: 概念、核算方法与案例研究[J]. 生态学报, 2013, 33(21): 6747 − 6761. OUYANG Zhiyun, ZHU Chunquan, YANG Guangbin, et al. Gross ecosystem product: concept, accounting framework and case study [J]. Acta Ecol Sin, 2013, 33(21): 6747 − 6761. [10] 国家林业和草原局. 森林生态系统服务功能评估规范: GB/T 38582—2020 [S]. 北京: 中国标准出版社, 2020. National Forestry and Grassland Administration. Specification for Assesment of Forest Ecosystem Service: GB/T 38582−2020 [S]. Beijing: China Standard Press, 2020. [11] 崔亚琴, 樊兰英, 刘随存, 等. 山西省森林生态系统服务功能评估[J]. 生态学报, 2019, 39(13): 4732 − 4740. CUI Yaqin, FAN Lanying, LIU Suicun, et al. Evaluation of forest ecosystem services value in Shanxi Province [J]. Acta Ecol Sin, 2019, 39(13): 4732 − 4740. [12] 黄龙生, 王兵, 牛香, 等. 济南市森林生态系统服务功能的维持机制[J]. 生态学报, 2018, 38(23): 8544 − 8554. HUANG Longsheng, WANG Bing, NIU Xiang, et al. Research on maintenance mechanism for ecosystem services function of forests in Ji’nan City [J]. Acta Ecol Sin, 2018, 38(23): 8544 − 8554. [13] 薛沛沛, 王兵, 牛香, 等. 武宁县、江山市和邵武市森林生态系统服务功能及其价值评估[J]. 水土保持学报, 2013, 27(5): 249 − 254. XUE Peipei, WANG Bing, NIU Xiang, et al. Forest ecosystem service and its evaluation in Wuning County, Jiangshan City and Shaowu City [J]. J Soil Water Conserv, 2013, 27(5): 249 − 254. [14] 李左玉, 董红先, 刘雷雷, 等. 浙江乌岩岭国家级自然保护区森林生态系统服务价值评估[J]. 浙江农林大学学报, 2020, 37(5): 891 − 897. LI Zuoyu, DONG Hongxian, LIU Leilei, et al. Evaluation of forest ecosystem service value in Wuyanling national nature reserve of Zhejiang Province [J]. J Zhejiang A&F Univ, 2020, 37(5): 891 − 897. [15] 吴钢, 肖寒, 赵景柱, 等. 长白山森林生态系统服务功能[J]. 中国科学(C辑: 生命科学), 2001, 31(5): 471 − 480. WU Gang, XIAO Han, ZHAO Jingzhu, et al. Ecosystem service function of forests in Changbai Mountain [J]. Sci China Ser C Life Sci, 2001, 31(5): 471 − 480. [16] 牛香. 森林生态效益分布式测算及其定量化补偿研究——以广东省和辽宁省为例[D]. 北京: 北京林业大学, 2012. NIU Xiang. Research on Distributed Assessment and Quantitative Compensation of Forest Ecological Benefits: Case Studies with Guangdong and Liaoning Provinces[D]. Beijing: Beijing Forestry University, 2012. [17] 黄龙生, 王兵, 牛香, 等. 济南市森林生态系统服务功能空间格局研究[J]. 生态学报, 2019, 39(17): 6477 − 6486. HUANG Longsheng, WANG Bing, NIU Xiang, et al. Spatial pattern of the ecosystem service function of forests in Ji’nan City [J]. Acta Ecol Sin, 2019, 39(17): 6477 − 6486. [18] 张国江, 季碧勇, 王文武, 等. 设区市森林资源市县联动监测体系研究[J]. 浙江农林大学学报, 2011, 28(1): 46 − 51. ZHANG Guojiang, JI Biyong, WANG Wenwu, et al. City-county synchronized monitoring system of forest resources in the city with districts under its jurisdiction [J]. J Zhejiang A&F Univ, 2011, 28(1): 46 − 51. [19] 陶吉兴, 张国江, 季碧勇, 等. 杭州市森林资源市县联动年度化监测的探索与实践[J]. 林业资源管理, 2014(4): 14 − 18. TAO Jixing, ZHANG Guojiang, JI Biyong, et al. Exploration and practice of integrated annual forest resources monitoring at both municipal and county levels in Hangzhou [J]. For Resour Manage, 2014(4): 14 − 18. [20] 季碧勇, 陶吉兴, 张国江, 等. 高精度保证下的浙江省森林植被生物量评估[J]. 浙江农林大学学报, 2012, 29(3): 328 − 334. JI Biyong, TAO Jixing, ZHANG Guojiang, et al. Zhejiang province’s forest vegetation biomass assessment for guaranteed accuracy [J]. J Zhejiang A&F Univ, 2012, 29(3): 328 − 334. [21] UK National Ecosystem Assessment. The UK National Ecosystem Assessment Technical Report (Technical Report: Introduction to the UK National Ecosystem Assessment)[M]. Cambridge: UNEP-WCMC, 2011. [22] 龚诗涵, 肖洋, 方瑜, 等. 中国森林生态系统地表径流调节特征[J]. 生态学报, 2016, 36(22): 7472 − 7478. GONG Shihan, XIAO Yang, FANG Yu, et al. Forest ecosystem surface run-off regulation characteristics in China [J]. Acta Ecol Sin, 2016, 36(22): 7472 − 7478. [23] 张永利, 杨锋伟, 王兵, 等. 中国森林生态系统服务功能研究[M]. 北京: 科学出版社, 2010. ZHANG Yongli, YANG Fengwei, WANG Bing, et al. Research on Forest Ecosystem Service Function in China [M]. Beijing: Science Press, 2010. [24] 全国森林资源标准化技术委员会. 国家森林资源连续清查数据处理统计规范: LY/T 1957—2011[S]. 北京: 中国标准出版社, 2011. National Forest Resources Standardization Technical Committee. Specification for Data Processing and Statistic in National Forest Inventory: LY/T 1957−2011 [S]. Beijing: China Standard Press, 2011. [25] 宋庆丰. 中国近40年森林资源变迁动态对生态功能的影响研究[D]. 北京: 中国林业科学研究院, 2015. SONG Qingfeng. Study on Impact of Forest Resource Dynamic Change on Forest Ecological Function in Recent 40 Years in China[D]. Beijing: Chinese Academy of Forestry, 2015. [26] 张志永. 杭州市典型城市森林生态保健功能动态变化分析[D]. 北京: 北京林业大学, 2014. ZHANG Zhiyong. Monitoring Dynamic Changes of Ecological Health Functions of Typical Urban Forests of Hangzhou[D]. Beijing: Beijing Forestry University, 2014. [27] STEHMAN S V, HANSEN M C, BROICH M, et al. Adapting a global stratified random sample for regional estimation of forest cover change derived from satellite imagery [J]. Remote Sensing Environ, 2011, 115(2): 650 − 658. [28] 林国忠. 森林资源二类调查方法的改进及监测体系研究[D]. 南京: 南京林业大学, 2010. LIN Guozhong. Improvement of Forest Inventory Method for Management Plan and Study on Monitoring System[D]. Nanjing: Nanjing Forestry University, 2010. 期刊类型引用(6)
1. 冯常辉,李林,张友昌,王琼珊,张教海,王孝刚,夏松波. 棉花纤维品质SNP标记研究进展. 湖北农业科学. 2024(S1): 5-9+13 .  百度学术
百度学术2. 龙遗磊,郑凯,齐静潇,蔡永生,邓晓娟,曲延英,陈全家. 海岛棉GbPIN1a基因的克隆与表达特性分析. 农业生物技术学报. 2022(11): 2086-2098 . 百度学术3. 孟超敏,耿翡翡,卿桂霞,周佳敏,张富厚,刘逢举. 陆地棉磷高效基因GhMGD3的克隆与表达分析. 浙江农林大学学报. 2022(06): 1203-1211 . 本站查看4. 闵凯丽,晁祥保,滕露,蔡永生,雷慧辰,严中建,郑凯,陈全家. 海岛棉GbHCT10基因的克隆与表达分析. 新疆农业科学. 2021(02): 206-215 . 百度学术5. 郭宝生,刘素恩,赵存鹏,王兆晓,王凯辉,李丹,刘旭,杜海英,耿军义. 转FBP7::iaaM基因陆地棉种质冀资139纤维品质性状杂种优势分析. 植物学报. 2021(02): 166-174 . 百度学术6. 赵柯柯,曲延英,段雅洁,石颖颖,范蓉,刘亚丽,陈全家. 海岛棉GbMYB5基因的克隆及表达分析. 分子植物育种. 2021(08): 2512-2520 . 百度学术其他类型引用(11)
-

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210328
 点击查看大图
点击查看大图
计量
- 文章访问数: 867
- HTML全文浏览量: 208
- PDF下载量: 46
- 被引次数: 17





