




-
南方集体林区是中国重要林区之一。2000年以来,是中国经济高速发展的时期,也是退耕还林和集体林权改革等政策开始推行和实施的阶段。在经济和政策的双重作用下,南方集体林区的土地利用产生了较为明显的变化,并引起了学界的关注。鲁亚楠等[1]研究了1980−2015年南方10省1市集体林的土地利用动态特征,发现耕地、草地面积减少,城乡建设用地显著增加,林地面积少量增加,而且2000年后土地利用类型变化速率明显加快。李明诗等[2]对浙江余杭地区的集体林进行研究后发现:虽然该地区的森林面积总体变化不大,但是森林破碎化程度一直加剧,可能造成近交衰败、能量传递和物质流动失衡、外来物种入侵加剧等生态后果[3]。可见对于南方集体林,不仅需要关注其森林面积的变化,还需要重视其景观格局的变化,以及这些变化引起的生态功能变化。浙江天目山地区森林覆盖率在80%以上,以集体林为主。随着市场需求的变化,树种类型发生了较大的变化,因此本研究以天目山保护区的国有林为对照,对集体林区近20 a的景观格局变化特征进行研究,以期为集体林社区的经济发展和生态环境建设提供思路。
-
研究区为浙江省杭州市临安区天目山镇(30°17′23″~30°24′39″N,119°23′31″~119°30′31″E)境内的保护区国有林(面积约984 hm2)和周边社区的集体林(面积约7 801 hm2),该区属中亚热带季风气候,年均降水量为1 822 mm,年均气温为8.9~15.8 ℃。保护区内拥有常绿阔叶林和落叶阔叶林等6个保护良好的原生植被类型;社区森林以常绿阔叶林、常绿落叶阔叶混交林、杉木Cunninghamia lanceolata林、竹林、山核桃Carya cathayensis林等次生林和人工林为主[4]。
-
航空和遥感影像数据分别来源于SPOT 5.0(2000年2月,分辨率2.5 m×2.5 m)、QuickBird(2010年12月,分辨率0.6 m×0.6 m)和WorldView(2017年11月,分辨率0.5 m×0.5 m)。森林资源二类调查数据来源于临安区林业局。行政界线及道路分布图来源于杭州市规划和自然资源局临安分局。2018年5−6月通过研究区居民走访和实地考察,对解译结果进行实地验证,并获取其他研究相关资料。
-
根据天目山的实际状况以及研究目的,并参照土地利用现状分类国家标准(GB/T 2010−2017),将研究区划分为10种景观类型。具体分类情况见表1。使用ArcGIS 10.1软件将遥感影像、森林资源二类调查数据及行政边界统一投影到CGCS2000坐标系,再根据遥感影像进行人工目视解译,最后根据实地调查数据对解译结果进行修正。
表 1 景观类型划分
Table 1. Standard for classification of landscape types
景观类型 描述与说明 生态林 经济林以外的,以发挥生态效益为主的各类天然或人工起源的乔木林 竹林 毛竹Phyllostachys edulis、早竹Phyllostachys praecox等 山核桃林 以山核桃Carya cathayensis等为主的经济林 香榧林 以香榧Torreya grandis ‘Merrillii’ 等为主的经济林 茶园 以茶树Camellia sinensis等 为主的经济林 建筑用地 居民房屋、生活基础设施、保护区和景区相关用地等 道路 宽度在3 m以上可以行车的道路 耕地 农业用地 水域 河流、湖泊、水库、坑塘、沟渠等 裸地 没有植被或者建筑覆盖的土地 -
在ArcGIS 10.1软件中将相邻时段的遥感解译结果进行空间叠加和统计分析,即可得到2000−2010年和2010−2017年这2个时段研究区景观类型面积转移状况[5]。景观格局分析选取景观边缘密度、景观形状指数、蔓延度、香农多样性指数、香农均匀度指数、斑块密度、最大斑块指数、平均边缘面积比和斑块结合度等指标,使用Fragstats 4.0软件进行计算。
-
在景观形成过程中,人类活动具有很大的影响。本研究主要从政策、市场和道路3方面对研究区的土地利用和景观格局的变化进行分析。
-
生态林是保护区的主体景观,竹林次之,山核桃林是面积最小的森林类型。其他景观类型面积占比均非常低。2000−2010年,保护区生态林面积减少了10.95 hm2,道路面积增加了5.12 hm2,建筑用地面积增长了3.44 hm2。2010−2017年,生态林面积增加了83.36 hm2,竹林和山核桃林面积分别减少了34.87和50.35 hm2,建筑用地等面积变化较小(图1和表2)。社区生态林面积比例比保护区面积低12%~20%;竹林面积比例约20%;山核桃林排名第3位,但是面积逐年增加。2000−2010年,生态林面积减少了242.98 hm2,竹林、山核桃林面积分别增加了47.37和275.67 hm2,建筑用地和道路面积分别增加了24.86和36.50 hm2。2010−2017年,生态林面积减少了127.89 hm2,竹林和茶园面积分别减少了125.74和38.05 hm2,山核桃林和香榧林面积分别增加了189.88和16.63 hm2,建筑用地和道路面积分别增加了50.55和9.03 hm2。
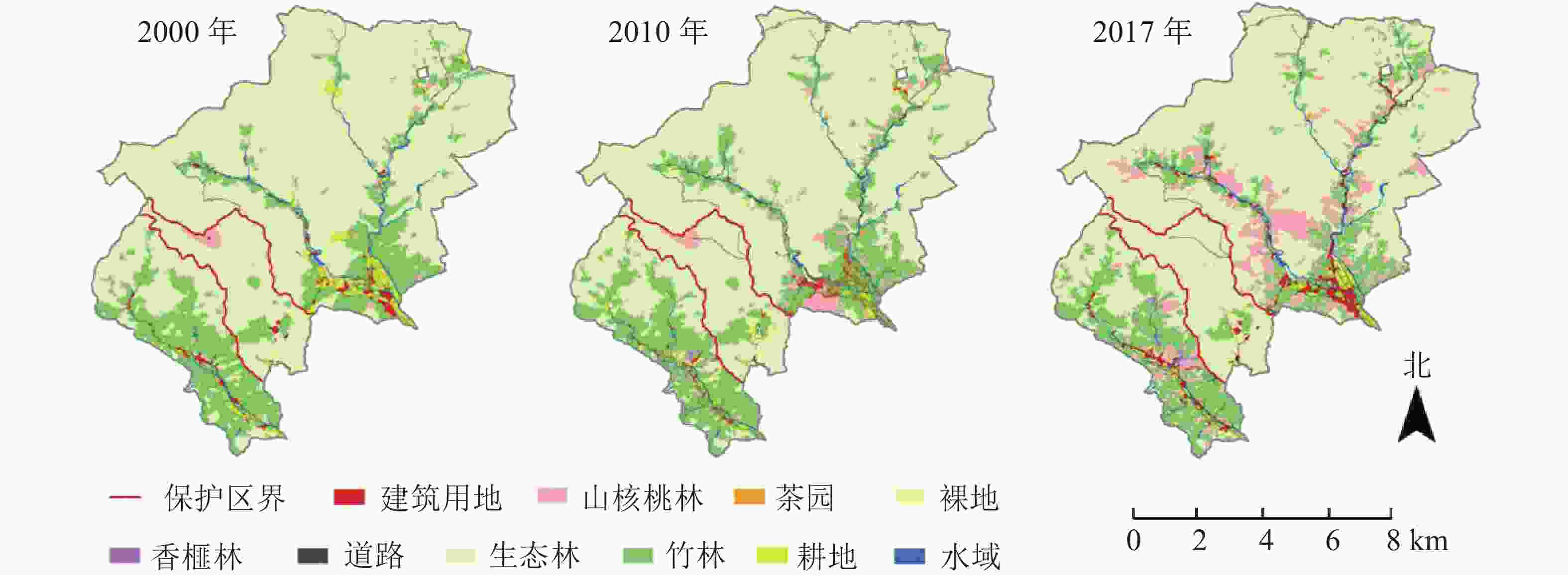
图 1 2000−2017年研究区景观格局时空变化示意图
Figure 1. Spatial and temporal changes of landscape pattern from 2000 to 2017
表 2 保护区和社区景观类型面积比例变化
Table 2. Area propration change of landscape types in reserve and communities
景观类型 保护区面积比例/% 社区面积比例/% 2000年 2010年 2017年 2000年 2010年 2017年 生态林 85.42 84.31 92.77 73.00 69.88 68.24 竹林 8.83 9.02 5.48 20.39 21.00 19.39 山核桃林 5.09 5.11 0.00 1.60 5.14 7.57 香榧林 − − − 0.04 0.10 0.31 茶园 − − − 0.54 0.57 0.08 建筑用地 0.50 0.85 0.78 0.87 1.08 1.73 道路 0.03 0.55 0.52 0.44 0.60 0.71 耕地 0.11 0.14 0.42 2.16 0.70 1.05 水域 0.03 0.02 0.03 0.82 0.83 0.82 裸地 − − − 0.14 0.12 0.10 说明:“−”表示研究区没有该景观类型 -
生态林占据了保护区的绝大部分区域,竹林主要集中在海拔350~900 m的游步道附近,建筑等主要分布在低海拔的景区服务中心一带。社区中竹林等经济林以各种形状和组合方式镶嵌在生态林中。竹林大部分集中分布于研究区的南部,呈块状分布,结合度非常高。山核桃林也主要分布于道路两侧。建筑和耕地主要沿着道路呈线状分布。
-
由表3可知:2000−2010年,保护区的景观类型变化特征为生态林向竹林、建筑用地和道路转化,转化面积分别为1.84、3.55和4.89 hm2;竹林向道路转化,转化面积分别为0.23 hm2。由表4可知:2010−2017年,保护区的景观类型变化特征为竹林、山核桃林、建筑用地向生态林转化,转化面积分别为31.87、50.35和0.53 hm2。
表 3 2000−2010年保护区景观类型面积转移矩阵
Table 3. Area transfer matrix in reserve from 2000 to 2010
景观类型 转移面积/hm2 生态林 竹林 山核桃林 建筑用地 道路 耕地 水域 2000年 生态林 830.16 1.84 0.21 3.55 4.89 840.65 竹林 86.98 0.23 87.21 山核桃林 50.13 50.13 建筑用地 4.94 4.94 道路 0.26 0.26 耕地 1.04 1.04 水域 0.03 0.24 0.27 2010年 830.16 88.82 50.34 8.49 5.38 1.07 0.24 984.71 表 4 2010−2017年保护区景观类型面积转移矩阵
Table 4. Area transfer matrix in reserve from 2010 to 2017
景观类型 转移面积/hm2 生态林 竹林 山核桃林 建筑用地 道路 耕地 水域 2010年 生态林 830.16 830.16 竹林 31.87 53.95 3.01 88.83 山核桃林 50.35 50.35 建筑用地 0.53 7.69 8.22 道路 0.23 5.15 5.38 耕地 1.10 0.03 1.13 水域 0.24 0.24 2017年 913.14 53.95 0.00 7.69 5.15 4.11 0.27 984.71 2000−2010年社区的景观类型变化特征为生态林向竹林、山核桃林、建筑用地和道路转化;竹林景观向山核桃林、香榧林、茶园、建筑用地和道路转化;耕地向竹林、山核桃、建筑用地和道路转化(表5)。2010−2017年的景观类型变化特征为生态林向竹林、山核桃林、香榧林和道路转化;竹林景观向山核桃林、香榧林、建筑用地、道路和耕地转化;茶园向竹林、山核桃林和道路转化;一部分耕地向建筑用地转化(表6)。
表 5 2000−2010年社区景观类型面积转移矩阵
Table 5. Area transfer matrix in communities from 2000 to 2010
景观类型 转移面积/hm2 生态林 竹林 山核桃林 香榧林 茶园 建筑用地 道路 耕地 水域 裸地 2000年 生态林 5 452.09 112.36 121.32 3.53 5.77 5 695.07 竹林 1 452.15 129.20 1.21 2.29 4.41 2.19 1 591.44 山核桃林 122.20 2.80 125.00 香榧林 2.77 2.77 茶园 41.97 41.97 建筑用地 67.73 67.73 道路 34.22 34.22 耕地 73.67 27.95 7.69 4.72 54.73 168.76 水域 63.97 63.97 裸地 0.71 1.21 9.06 10.98 2010年 5 452.09 1 638.18 400.67 7.49 44.26 84.57 46.9 54.73 63.97 9.06 7 801.92 表 6 2010−2017年社区景观类型面积转移矩阵
Table 6. Area transfer matrix in communities from 2010 to 2017
景观类型 转移面积/hm2 生态林 竹林 山核桃林 香榧林 茶园 建筑用地 道路 耕地 水域 裸地 2010年 生态林 5 324.19 79.45 38.47 6.36 3.61 5 452.08 竹林 1 416.02 131.98 10.27 34.20 3.44 42.29 1 638.20 山核桃林 400.67 400.67 香榧林 7.49 7.49 茶园 16.97 19.10 6.11 1.98 44.16 建筑用地 84.57 84.57 道路 46.71 46.71 耕地 15.23 39.49 54.72 水域 63.68 63.68 裸地 1.12 7.94 9.06 2017年 5 324.19 1 512.44 590.22 24.12 6.11 135.12 55.74 81.78 63.68 7.94 7 801.92 -
保护区近20 a的景观格局整体变化不大,社区的景观破碎化程度则一直加剧(表7)。社区生态林的优势地位一直在下降,与保护区的演化趋势相反。社区各景观类型斑块密度均显著高于保护区,而且生态林、竹林、山核桃、香榧的斑块密度逐步增长。生态林、竹林的最大斑块指数逐步下降,山核桃林、香榧的最大斑块指数逐步上升。除了茶园以外,几乎所有景观类型的平均边缘面积比都呈上升趋势。生态林和竹林的斑块结合度呈下降趋势,山核桃和建筑用地呈上升趋势。
表 7 2000−2017年研究区类型水平景观格局特征
Table 7. Evolution of landscape patterns in class level from 2000 to 2017
年份 景观类型 斑块密度/(个·hm−2) 最大斑块指数/% 平均边缘面积比 斑块结合度/% 保护区 社区 保护区 社区 保护区 社区 保护区 社区 2000 生态林 0.20 0.55 4.35 90.82 465 1 269 99.99 99.69 竹林 1.02 3.86 0.10 51.35 1 005 1 392 99.48 99.44 山核桃林 0.30 1.54 1.98 5.57 591 968 96.22 97.22 香榧林 − 0.02 − 0.22 − 568 − 97.30 茶园 − 0.59 − 3.01 − 1 107 − 96.69 建筑用地 0.61 1.38 0.33 11.54 1 178 1 727 97.34 96.15 耕地 0.10 1.45 0.11 12.13 734 1 935 97.15 97.79 水域 0.10 0.59 0.03 1.88 866 2 771 94.32 98.12 裸地 − 0.42 − 0.42 − 1 476 − 94.20 2010 生态林 0.61 0.99 4.86 80.00 923 1 995 99.99 99.55 竹林 0.81 4.87 0.10 46.73 1 149 2 186 95.55 99.40 山核桃林 1.02 3.94 2.05 14.31 1 968 1 442 99.51 97.72 香榧林 − 0.09 − 0.34 − 660 − 97.34 茶园 − 1.72 − 3.44 − 1 231 − 96.71 建筑用地 1.02 3.26 0.32 8.70 1 245 2 484 96.66 96.47 耕地 0.10 3.24 0.14 2.57 786 2 176 97.52 95.87 水域 0.10 0.81 0.02 1.82 995 3 013 93.98 97.78 裸地 − 0.12 − 2.74 − 1 409 − 96.32 2017 生态林 0.41 4.22 5.06 59.13 996 2 119 99.99 99.51 竹林 0.51 7.86 0.10 41.83 1 492 4 872 92.84 99.18 山核桃林 − 6.32 − 15.57 − 2 906 − 97.96 香榧林 − 0.27 − 3.03 − 2 744 − 96.37 茶园 − 0.06 − 1.68 − 830 − 96.42 建筑用地 0.91 3.15 0.32 14.11 995 3 030 96.89 97.42 耕地 0.30 2.56 0.14 8.95 707 3 928 97.68 97.24 水域 0.10 1.19 0.02 1.90 961 3 635 94.36 97.42 裸地 − 0.35 − 0.44 − 1 600 − 95.33 说明:“−”表示研究区域没有该景观类型 -
近20 a保护区整体呈现生态林优势地位上升的趋势,而社区则整体呈现形状趋于复杂,景观异质性程度升高,生态林聚集度和优势度下降,各景观类型的分布趋于匀质化的趋势,具体表现为社区景观边缘密度、景观形状指数、香农多样性指数和均匀度指数均逐渐上升,而蔓延度逐渐下降(表8)。
表 8 2000−2017年研究区景观水平景观格局特征
Table 8. Evolution of landscape patterns in landscape level from 2000 to 2017
年份 景观边缘密度/(m·hm−2) 景观形状指数 蔓延度/% 香农多样性指数 香农均匀度指数 保护区 社区 保护区 社区 保护区 社区 保护区 社区 保护区 社区 2000 24.72 120.86 4.05 6.93 87.45 73.96 0.50 0.96 0.24 0.47 2010 40.66 162.71 5.30 8.85 86.02 71.00 0.54 1.03 0.26 0.52 2017 35.34 176.12 4.88 9.67 87.32 68.26 0.49 1.15 0.24 0.56 -
保护区内的生态林受到严格保护,基础设施建设也有严格的限制,这是其景观格局变化远低于社区的政策因素。而且2010年以来,保护区对毛竹林进行了有规划的采伐和抚育,竹林面积显著下降,采伐迹地逐渐演替为生态林。而社区因为没有严格的保护政策,景观格局受到较强的人为影响。
-
社区的景观格局变化主要由经济林的扩张和经济林树种的变化引起。竹林、山核桃林、香榧林的扩张导致了生态林面积的逐年下降,竹林向山核桃林和香榧林转化导致了景观破碎度和异质性的增加。根据统计年鉴和实地走访可知,竹产品笋干价格走低,销售困难,山核桃和香榧干果的需求上升且收益可观,是引起了经济林树种和面积变化的重要原因。
-
生态林与经济林之间的转化主要发生在道路两侧。这是因为研究区地形多为丘陵地带,没有便利的交通就难以运输生产物资和林产品,所以道路也是社区经济林面积扩张的重要因素之一。
-
随着社会经济水平的提高,人们对森林生态和社会文化功能的需求不断增加。但是在实际的生产和实践过程中,森林的生态、经济和社会价值之间往往存在着此消彼长的权衡关系。人为活动引起的土地利用变化是生态系统服务功能变化的最主要因素。城镇和道路的建设有助于经济效益的提升,但是这两者的扩张会导致生态效益的降低。已有研究表明:土地利用强度的提升会通过增加景观破碎度导致水土和养分的过量流失,提高生态系统的敏感性和脆弱性[6-8]。例如柳冬青等[9]对陇中黄土丘陵区的研究表明:随着土地利用强度的增加,该区域生态系统供给服务比例升高,但是调节和支持服务比例逐步降低。生态完整性好、景观破碎度的区域生态效益更高[10-11]。
本研究发现:近20 a来天目山集体林区的生态林不断向经济林转移,同时经济林种类也因为市场需求的变化而变化,从毛竹转向山核桃再转向香榧,因此土地利用的强度也较高。同时社区的建筑用地和道路面积占比虽然较低,但也在不断升高。土地利用的变化引起了社区生态林景观优势地位的不断下降,整个林区的景观破碎度、多样性和均匀性逐步升高。当地居民的经济需求是导致这一现象产生的主要原因,道路的扩张是促进这一现象的重要因素。社区土地利用的转变以及景观格局的变化很有可能会导致水土流失、生物多样性下降等一系列的生态问题[12-14],这不利于社区长期的经济发展和生态安全,为此有必要探索生态效益与社会经济效益协同发展的集体林经营机制和措施。
依据森林生态系统经营理念,结合本研究的结果,初步提出以下对策:首先需要明确森林经营的目的是实现人与自然的和谐共处,既满足当代人的需求又不损害后代人的利益[15];其次将经营水平从林分尺度转变为景观水平,将社区的居民区、农田、道路和森林作为一个整体进行经营,优化利用景观资源、权衡和协同多种生态系统的服务功能,这可以减少森林生态系统及其与之紧密相连的社区环境和经济的风险,提升自然资源供给、食物安全和社会发展的长期稳定性,同时还可以提高景观的文化价值[16-17]。在明确了这2个前提的基础上,通过政策限制经济林面积、提高经济林单位面积效益、退竹换阔等措施实现森林的可持续经营目标。对比保护区可以发现:严格的保护政策是保护区景观格局破碎度远低于社区的重要原因。因此,在社区也可以出台相应制度以控制经济林面积的扩张,避免生态林继续转化为经济林。需要注意的是:①由于集体林土地权属的问题,政策制定时还应考虑其特殊性和可行性。②政府通过技术支持和引导等手段提高经济林单位面积效益,例如开展林下经济、或结合当地的旅游产业开展森林采摘等体验式旅游形式,以缓解居民为追求经济收入一味扩大经济林面积的现象。③本研究还发现:由于自身较强的扩张能力,竹林在人为干扰较少区域仍在继续自行扩张侵占生态林,因此还需要对这些区域的竹林进行监测,必要时可进行人为砍伐。④道路是促进经济林扩张的重要条件,但也是人们享受森林生态系统服务的廊道,因此在研究区道路建设过程中,对生态系统影响较大的大型公路修建需要谨慎考虑,建议修建一些特色绿道。
Forests landscape pattern changes and driving forces in Mount Tianmu
-
摘要:
目的 研究南方集体林区的景观格局变化特征及驱动因素,可为森林可持续经营提供科学依据。 方法 采用景观类型面积转移矩阵和景观格局指数分析方法,对比研究了浙江天目山自然保护区国有林和周边社区集体林2000、2010和2017年的景观格局变化特征。 结果 生态林是保护区和社区的优势景观。保护区整体呈生态林面积增加,经济林面积减少趋势,而社区生态林面积逐年减少,经济林面积不断增加;保护区的竹林和山核桃Carya cathayensis林向生态林转化,社区呈现生态林向经济林、建筑、道路转化,经济林之间相互转化,经济林和耕地相互转化的特征。保护区的景观格局变化不大,社区景观的破碎度、异质性和均匀性逐年上升,生态林的优势地位逐年下降。政策因素、市场需求变化和道路是导致保护区和社区森林景观格局变化差异的主要驱动因素。 结论 社区景观格局的变化会影响森林的生态和文化功能,可以从政策、旅游业发展和道路建设等方面制定相关措施。图1表8参17 Abstract:Objective This research studied the characteristics and driving forces of landscape pattern change in southern collective forest area to provide scientific basis for sustainable forest management. Method Landscape pattern change characteristics of state-owned forest in Zhejiang Tianmu Mount nature reserve and collective forest in surrounding community of the reserve in 2000, 2010 and 2017 were compared and studied by using landscape type area transfer matrix and landscape pattern index analysis method. Result Ecological forest was the dominant landscape in the reserve and communities. The area of ecological forest increased in the reserve and decreased in the communities, while the area of economic forest changed in an opposite way. The bamboo forest and hickory forest transformed to ecological forest in the reserve, while ecological forest transformed to economic forest, buildings and roads, then mutual transformation between different kinds of economic forest or mutual transformation between economic forest and cultivated land in the communities. The landscape pattern in the reserve changed little, but landscape fragmentation, heterogeneity and uniformity in the communities increased year by year, and the ascendancy of ecological forest had been on the decline. Policy factors, market demand changes and roads were the main driving forces behind the changes of forest landscape patterns. Conclusion Changes of landscape patterns in community may affect ecological and cultural functions of forest. Measures, such as policy making, tourism development and road construction can be taken. [Ch, 1 fig. 8 tab. 17 ref.] -
Key words:
- forest management /
- Mount Tianmu /
- collective forest /
- landscape pattern /
- driving force /
- community
-
图 1 2000−2017年研究区景观格局时空变化示意图
Figure 1 Spatial and temporal changes of landscape pattern from 2000 to 2017
表 1 景观类型划分
Table 1. Standard for classification of landscape types
景观类型 描述与说明 生态林 经济林以外的,以发挥生态效益为主的各类天然或人工起源的乔木林 竹林 毛竹Phyllostachys edulis、早竹Phyllostachys praecox等 山核桃林 以山核桃Carya cathayensis等为主的经济林 香榧林 以香榧Torreya grandis ‘Merrillii’ 等为主的经济林 茶园 以茶树Camellia sinensis等 为主的经济林 建筑用地 居民房屋、生活基础设施、保护区和景区相关用地等 道路 宽度在3 m以上可以行车的道路 耕地 农业用地 水域 河流、湖泊、水库、坑塘、沟渠等 裸地 没有植被或者建筑覆盖的土地  下载: 导出CSV
下载: 导出CSV
表 2 保护区和社区景观类型面积比例变化
Table 2. Area propration change of landscape types in reserve and communities
景观类型 保护区面积比例/% 社区面积比例/% 2000年 2010年 2017年 2000年 2010年 2017年 生态林 85.42 84.31 92.77 73.00 69.88 68.24 竹林 8.83 9.02 5.48 20.39 21.00 19.39 山核桃林 5.09 5.11 0.00 1.60 5.14 7.57 香榧林 − − − 0.04 0.10 0.31 茶园 − − − 0.54 0.57 0.08 建筑用地 0.50 0.85 0.78 0.87 1.08 1.73 道路 0.03 0.55 0.52 0.44 0.60 0.71 耕地 0.11 0.14 0.42 2.16 0.70 1.05 水域 0.03 0.02 0.03 0.82 0.83 0.82 裸地 − − − 0.14 0.12 0.10 说明:“−”表示研究区没有该景观类型
下载: 导出CSV
表 3 2000−2010年保护区景观类型面积转移矩阵
Table 3. Area transfer matrix in reserve from 2000 to 2010
景观类型 转移面积/hm2 生态林 竹林 山核桃林 建筑用地 道路 耕地 水域 2000年 生态林 830.16 1.84 0.21 3.55 4.89 840.65 竹林 86.98 0.23 87.21 山核桃林 50.13 50.13 建筑用地 4.94 4.94 道路 0.26 0.26 耕地 1.04 1.04 水域 0.03 0.24 0.27 2010年 830.16 88.82 50.34 8.49 5.38 1.07 0.24 984.71
下载: 导出CSV
表 4 2010−2017年保护区景观类型面积转移矩阵
Table 4. Area transfer matrix in reserve from 2010 to 2017
景观类型 转移面积/hm2 生态林 竹林 山核桃林 建筑用地 道路 耕地 水域 2010年 生态林 830.16 830.16 竹林 31.87 53.95 3.01 88.83 山核桃林 50.35 50.35 建筑用地 0.53 7.69 8.22 道路 0.23 5.15 5.38 耕地 1.10 0.03 1.13 水域 0.24 0.24 2017年 913.14 53.95 0.00 7.69 5.15 4.11 0.27 984.71
下载: 导出CSV
表 5 2000−2010年社区景观类型面积转移矩阵
Table 5. Area transfer matrix in communities from 2000 to 2010
景观类型 转移面积/hm2 生态林 竹林 山核桃林 香榧林 茶园 建筑用地 道路 耕地 水域 裸地 2000年 生态林 5 452.09 112.36 121.32 3.53 5.77 5 695.07 竹林 1 452.15 129.20 1.21 2.29 4.41 2.19 1 591.44 山核桃林 122.20 2.80 125.00 香榧林 2.77 2.77 茶园 41.97 41.97 建筑用地 67.73 67.73 道路 34.22 34.22 耕地 73.67 27.95 7.69 4.72 54.73 168.76 水域 63.97 63.97 裸地 0.71 1.21 9.06 10.98 2010年 5 452.09 1 638.18 400.67 7.49 44.26 84.57 46.9 54.73 63.97 9.06 7 801.92
下载: 导出CSV
表 6 2010−2017年社区景观类型面积转移矩阵
Table 6. Area transfer matrix in communities from 2010 to 2017
景观类型 转移面积/hm2 生态林 竹林 山核桃林 香榧林 茶园 建筑用地 道路 耕地 水域 裸地 2010年 生态林 5 324.19 79.45 38.47 6.36 3.61 5 452.08 竹林 1 416.02 131.98 10.27 34.20 3.44 42.29 1 638.20 山核桃林 400.67 400.67 香榧林 7.49 7.49 茶园 16.97 19.10 6.11 1.98 44.16 建筑用地 84.57 84.57 道路 46.71 46.71 耕地 15.23 39.49 54.72 水域 63.68 63.68 裸地 1.12 7.94 9.06 2017年 5 324.19 1 512.44 590.22 24.12 6.11 135.12 55.74 81.78 63.68 7.94 7 801.92
下载: 导出CSV
表 7 2000−2017年研究区类型水平景观格局特征
Table 7. Evolution of landscape patterns in class level from 2000 to 2017
年份 景观类型 斑块密度/(个·hm−2) 最大斑块指数/% 平均边缘面积比 斑块结合度/% 保护区 社区 保护区 社区 保护区 社区 保护区 社区 2000 生态林 0.20 0.55 4.35 90.82 465 1 269 99.99 99.69 竹林 1.02 3.86 0.10 51.35 1 005 1 392 99.48 99.44 山核桃林 0.30 1.54 1.98 5.57 591 968 96.22 97.22 香榧林 − 0.02 − 0.22 − 568 − 97.30 茶园 − 0.59 − 3.01 − 1 107 − 96.69 建筑用地 0.61 1.38 0.33 11.54 1 178 1 727 97.34 96.15 耕地 0.10 1.45 0.11 12.13 734 1 935 97.15 97.79 水域 0.10 0.59 0.03 1.88 866 2 771 94.32 98.12 裸地 − 0.42 − 0.42 − 1 476 − 94.20 2010 生态林 0.61 0.99 4.86 80.00 923 1 995 99.99 99.55 竹林 0.81 4.87 0.10 46.73 1 149 2 186 95.55 99.40 山核桃林 1.02 3.94 2.05 14.31 1 968 1 442 99.51 97.72 香榧林 − 0.09 − 0.34 − 660 − 97.34 茶园 − 1.72 − 3.44 − 1 231 − 96.71 建筑用地 1.02 3.26 0.32 8.70 1 245 2 484 96.66 96.47 耕地 0.10 3.24 0.14 2.57 786 2 176 97.52 95.87 水域 0.10 0.81 0.02 1.82 995 3 013 93.98 97.78 裸地 − 0.12 − 2.74 − 1 409 − 96.32 2017 生态林 0.41 4.22 5.06 59.13 996 2 119 99.99 99.51 竹林 0.51 7.86 0.10 41.83 1 492 4 872 92.84 99.18 山核桃林 − 6.32 − 15.57 − 2 906 − 97.96 香榧林 − 0.27 − 3.03 − 2 744 − 96.37 茶园 − 0.06 − 1.68 − 830 − 96.42 建筑用地 0.91 3.15 0.32 14.11 995 3 030 96.89 97.42 耕地 0.30 2.56 0.14 8.95 707 3 928 97.68 97.24 水域 0.10 1.19 0.02 1.90 961 3 635 94.36 97.42 裸地 − 0.35 − 0.44 − 1 600 − 95.33 说明:“−”表示研究区域没有该景观类型
下载: 导出CSV
表 8 2000−2017年研究区景观水平景观格局特征
Table 8. Evolution of landscape patterns in landscape level from 2000 to 2017
年份 景观边缘密度/(m·hm−2) 景观形状指数 蔓延度/% 香农多样性指数 香农均匀度指数 保护区 社区 保护区 社区 保护区 社区 保护区 社区 保护区 社区 2000 24.72 120.86 4.05 6.93 87.45 73.96 0.50 0.96 0.24 0.47 2010 40.66 162.71 5.30 8.85 86.02 71.00 0.54 1.03 0.26 0.52 2017 35.34 176.12 4.88 9.67 87.32 68.26 0.49 1.15 0.24 0.56
下载: 导出CSV
-
[1] 鲁亚楠,姚顺波. 基于经济和政策双重作用的南方集体林区土地利用时空变化[J]. 南京林业大学学报(自然科学版), 2018, 42(5): 163 − 171. LU Ya’nan, YAO Shunbo. Spatial temporal dynamic characteristics for land use of the southern collective forest region based on the function of economy and policy [J]. J Nanjing For Univ Nat Sci Ed, 2018, 42(5): 163 − 171. [2] 李明诗,徐祎凡,李云梅,等. 南方集体林区森林破碎化模式及趋势遥感刻画: 以浙江余杭为例[J]. 南京林业大学学报(自然科学版), 2010, 34(4): 135 − 139. LI Mingshi, XU Yifan, LI Yunmei, et al. Remote sensing based characterization of fragmentation patterns and trends of the collective forests in southern China: a case study from Yuhang City of Zhejiang Province [J]. J Nanjing For Univ Nat Sci Ed, 2010, 34(4): 135 − 139. [3] 刘建锋,肖文发,江泽平,等. 景观破碎化对生物多样性的影响[J]. 林业科学研究, 2005, 18(2): 222 − 226. LIU Jianfeng, XIAO Wenfa, JIANG Zeping, et al. A study on the influence of landscape fragmentation on biodiversity [J]. For Res, 2005, 18(2): 222 − 226. [4] 重修西天目山山志编撰委员会. 西天目山志[M]. 北京: 方志出版社, 2009. [5] 林柳璇,尤添革,刘金福,等. 1985−2015年厦门市土地利用变化及驱动力[J]. 福建农林大学学报(自然科学版), 2019, 48(1): 103 − 110. LIN Liuxuan, YOU Tiange, LIU Jinfu, et al. Land use changes and driving forces in Xiamen City from 1985 to 2015 [J]. J Fujian Agric For Univ Nat Sci Ed, 2019, 48(1): 103 − 110. [6] XU Ying, TANG Haiping, WANG Bojie, et al. Effects of land-use intensity on ecosystem services and human well-being: a case study in Huailai County, China [J]. Environ Earth Sci, 2016, 75(5): 416 − 425. [7] 韩蕾,孔祥斌,郭洪峰,等. 西南山区生态敏感区土地生态安全评价: 以秀山县为例[J]. 水土保持研究, 2015, 22(1): 229 − 234. HAN Lei, KONG Xiangbin, GUO Hongfeng, et al. Study on land eco-security evaluation of ecological sensitive area in southwest mountains: a case study in Xiushan County [J]. Res Soil Water Conserv, 2015, 22(1): 229 − 234. [8] 刘亚香,李阳兵,易兴松,等. 贵州典型坝子土地利用强度空间演变及景观格局响应[J]. 应用生态学报, 2017, 28(11): 3691 − 3702. LIU Yaxiang, LI Yangbing, YI Xingsong, et al. Spatial evolution of land use intensity and landscape pattern response of the typical basins in Guizhou Province, China [J]. Chin J Appl Ecol, 2017, 28(11): 3691 − 3702. [9] 柳冬青,张金茜,巩杰,等. 陇中黄土丘陵区土地利用强度-生态系统服务-人类福祉时空关系研究: 以安定区为例[J]. 生态学报, 2019, 39(2): 637 − 648. LIU Dongqing, ZHANG Jinxi, GONG Jie, et al. Spatial and temporal relations among land-use intensity, ecosystem services, and human well-being in the Longzhong Loess Hilly Region: a case study of the Anding District, Gansu Province [J]. Acta Ecol Sin, 2019, 39(2): 637 − 648. [10] 欧阳志云,王效科,苗鸿. 中国陆地生态系统服务功能及其生态经济价值的初步研究[J]. 生态学报, 1999, 19(5): 607 − 613. OUYANG Zhiyun, WANG Xiaoke, MIAO Hong. A primary study on Chinese terrestrial ecosystem services and their ecological economic values [J]. Acta Ecol Sin, 1999, 19(5): 607 − 613. [11] 胡和兵,刘红玉,郝敬锋,等. 城市化流域生态系统服务价值时空分异特征及其对土地利用程度的响应[J]. 生态学报, 2013, 33(8): 2565 − 2576. HU Hebing, LIU Hongyu, HAO Jingfeng, et al. Spatio-temporal variation in the value of ecosystem services and its response to land use intensity in an urbanized watershed [J]. Acta Ecol Sin, 2013, 33(8): 2565 − 2576. [12] ZIMMERMANN P, TASSE E, LEITINGER G, et al. Effects of land-use and land-cover pattern on landscape-scale biodiversity in the European Alps [J]. Agric Ecosyst Environ, 2010, 139(1/2): 13 − 22. [13] TRISURAT Y Y, SHIRAKAWA H, JOHNSTON J M. Land-use/land-cover change from socio-economic drivers and their impact on biodiversity in Nan Province, Thailand [J]. Sustainability, 2019, 11(3): 1 − 22. [14] 王航,秦奋,朱筠,等. 土地利用及景观格局演变对生态系统服务价值的影响[J]. 生态学报, 2017, 37(4): 1286 − 1296. WANG Hang, QIN Fen, ZHU Jun, et al. The effects of land use structure and landscape pattern change on ecosystem service values [J]. Acta Ecol Sin, 2017, 37(4): 1286 − 1296. [15] 蒋有绪. 森林可持续经营与林业的可持续发展[J]. 世界林业研究, 2001, 14(2): 1 − 8. JIANG Youxu. Sustainable management and sustainable development of forestry [J]. World For Res, 2001, 14(2): 1 − 8. [16] 刘世荣,代力民,温远光,等. 面向生态系统服务的森林生态系统经营: 现状、挑战与展望[J]. 生态学报, 2015, 35(1): 1 − 9. LIU Shirong, DAI Limin, WEN Yuanguang, et al. A review on forest ecosystem management towards ecosystem services: status, challenges, and future perspectives [J]. Acta Ecol Sin, 2015, 35(1): 1 − 9. [17] 张冉,王义民,畅建霞,等. 基于水资源分区的黄河流域土地利用变化对人类活动的响应[J]. 自然资源学报, 2019, 34(2): 274 − 287. ZHANG Ran, WANG Yimin, CHANG Jianxia, et al. Response of land use change to human activities in the Yellow River Basin based on water resources division [J]. J Nat Resour, 2019, 34(2): 274 − 287. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20190306
 点击查看大图
点击查看大图
计量
- 文章访问数: 3145
- HTML全文浏览量: 914
- PDF下载量: 123
- 被引次数: 0
