





-
DNA拓扑异构酶(DNA topoisomerase,TOP)是一类能够改变DNA拓扑结构的酶,在细胞的一系列基础生命活动如DNA复制、转录和染色体分离等过程中发挥着重要作用[1−2]。DNA拓扑异构酶在1971年被首次发现,命名为ω蛋白,即DNA拓扑异构酶Ⅰ,是大肠埃希菌Escherichia coli中能够特异性地将负超螺旋DNA解旋的一种拓扑异构酶[3−4]。根据DNA拓扑异构酶的功能和作用机制,可以将其分成Ⅰ型和Ⅱ型DNA拓扑异构酶,Ⅰ型TOP只切割DNA的1条链,不具有腺苷三磷酸(ATP)依赖性,Ⅱ型TOP能切割DNA的2条链并形成双链断裂,具有ATP依赖性[5]。真核生物中,Ⅱ型DNA拓扑异构酶包括ⅡA型DNA拓扑异构酶和ⅡB型DNA拓扑异构酶:ⅡA型DNA拓扑异构酶在高等真核生物(如哺乳动物)中有2种异构体,分别为TOP2α和TOP2β,在低等真核生物中通常只有1种;ⅡB型DNA拓扑异构酶在真核生物中主要是与酵母Spo11同源的ⅡB型DNA拓扑异构酶,以及植物中与古细菌ⅡB型DNA拓扑异构酶同源的TOPⅥ[6]。
在动物细胞中的研究表明:Ⅱ型DNA拓扑异构酶主要在复制和转录过程中对DNA双链进行解旋,并且确保有丝分裂过程中姐妹染色单体的正确分离,对染色质的结构和凝缩也有重要作用[7]。TOP2α对细胞周期起重要调节作用,保证分裂旺盛细胞的活性,不分裂细胞中则没有TOP2α;TOP2β对小鼠Mus musculus胚胎发育过程中神经元的形成具有重要影响,缺失TOP2β的小鼠出生后无法存活[6, 8]。DNA拓扑异构酶还常被作为抗癌药物的靶标蛋白,通过抑制DNA拓扑异构酶活性导致复制叉停滞,形成双链断裂并干扰细胞周期,进而抑制肿瘤细胞的快速增殖[9−11]。植物中已对拟南芥Arabidopsis thaliana、小麦Triticum aestivum、花椰菜Brassica oleracea var. botrytis等的DNA拓扑异构酶进行了分离和生化性质等方面的研究[12−17],但DNA拓扑异构酶的作用机制研究大多集中在动物和酵母系统里,关于植物DNA拓扑异构酶作用机制的研究较少。研究表明:在烟草Nicotiana tabaccum中过表达Ⅱ型DNA拓扑异构酶能够改变烟草植株形态,提高烟草的盐胁迫耐受性[18]。关于拟南芥中DNA拓扑异构酶Ⅱ的研究表明:TOPⅡ和染色体运动有助于消除减数分裂过程中纠缠染色体之间的连锁[19]。研究发现:topⅡ突变体拟南芥幼苗的根生长受到明显抑制,表明TOPⅡ在根生长和细胞周期调控中发挥作用,还可通过同源重组的方式进行有丝分裂中的DNA修复和减数分裂中的DNA双链断裂(DSBs)修复[20],并与TOPⅡ结合蛋白(TopBP1)在DNA修复、有丝分裂和减数分裂过程中共同发挥作用[21]。
杨树Populus作为研究木本植株生长发育的模式树种,在DNA拓扑异构酶的功能和作用机制等方面仍有许多空白。本研究以银腺杨‘84K’ Populus alba × P. glandulosa ‘84K’(84K杨)为研究对象,利用生物信息学及分子生物学方法,对杨树DNA拓扑异构酶基因功能进行初步探究,以期为DNA拓扑异构酶基因在木本植物中的作用机制和木本植物分子育种提供理论依据。
-
以84K杨组培苗为研究材料,组培苗生长温度为(24±2) ℃,光照为16 h/8 h (光照/黑暗)。土培苗为将生长约4周的组培苗从1/2 MS培养基移栽至营养土中,在温度(24±2) ℃,光照16 h/8 h (光照/黑暗),相对湿度60%~80%的土培室中培养。
-
提前高温处理研钵、药匙等直接与样品接触的试验器具。用预冷的研钵将植物材料在液氮中快速研磨成细腻粉末,取约100 mg粉末至RNase-free离心管中,使用商业购买的RNAprep Pure多糖多酚植物总RNA提取试剂盒(TIANGEN #DP441)提取84K杨不同组织的总RNA。提取得到的RNA用商业购买的FastKing cDNA第一链合成试剂盒(TIANGEN #KR116)进行反转录合成cDNA。
-
84K杨基因组数据来源于国家基因库序列归档系统(CNGB Nucleotide Sequence Archive,CNSA),项目号为CNP0000339[22]。根据PagTOP2b基因蛋白质编码序列(CDS),用Oligo 7软件设计基因全长序列克隆扩增引物,以从84K杨中提取的RNA反转录得到的cDNA为模板进行PagTOP2b基因的克隆。
在84K杨基因组数据库的GFF文件中提取PagTOP2b基因非翻译区、外显子和内含子的长度及染色体位置信息,使用TBtools软件绘制基因结构图[23]。在美国国家生物技术信息中心(NCBI)网站(
https://www.ncbi.nlm.nih.gov/ )的保守结构域数据库(conserved domain database,CDD)中搜索蛋白序列的保守结构域[24]。在SOPMA (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa%20_sopma.html )网站上进行PagTOP2b蛋白二级结构在线分析。并通过SWISS-MODEL(https://swissmodel.expasy.org/ )在线预测PagTOP2b蛋白三级结构,结合相关注释信息绘图分析[25]。使用ExPASy (
https://web.expasy.org/protparam/ )在线分析网站对PagTOP2b蛋白的序列长度、蛋白分子质量、等电点、亲水性平均系数、脂肪族氨基酸指数、不稳定指数等蛋白质理化性质进行分析[26]。并在Cell-Ploc 2.0网站中(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/ )选择Plant-mPLoc进行PagTOP2b蛋白亚细胞定位预测[27]。 -
在Primer3 网站上以PagTOP2b基因CDS序列为模板设计实时荧光定量PCR特异性引物(表1)。以待分析样品的cDNA为模板,根据商业购买的荧光定量PCR试剂ChamQ SYBR qPCR Master Mix (Vazyme #Q311-02/03)使用说明配置反应体系,选择ACTIN基因为内参基因。在Applied Biosystems QuantStudio 3 (Thermo Fisher Scientific)实时荧光定量PCR系统中进行反应。设置3个生物学重复和4次技术重复,取平均值作为后续计算的阈值循环数(Ct值),采用
$2^{-\Delta\Delta C_{t}}$ 法计算目的基因的相对表达水平。表 1 基因克隆及实时荧光定量PCR (RT-qPCR)引物
Table 1. Primers for gene clone and RT-qPCR
引物名称 引物序列(5′→3′) PagTOP2b F: ATGGCGGCAGAGACCAAGAAAAT R: GTCAGAGGAAGAGTCAGAATCATCAT PagTOP2b qrtF: AGACTACTGGTGGGAGAAAT qrtR: GTCACCTTGGTCCAATTCTC ACTIN qrtF: AAACTGTAATGGTCCTCCCTCCG qrtR: GCATCATCACAATCACTCTCCGA 说明:F表示基因克隆的正向引物;R表示基因克隆的反向引物;qrtF表示RT-qPCR的正向引物;qrtR表示RT-qPCR的反向引物 -
遗传转化所用PagTOP2b基因过表达载体pCAMBIA-S1300-flag-PagTOP2b为实验室保存。农杆菌Agrobacterium tumefaciens GV3101化学感受态细胞购自上海唯地生物(#AC1001)。将载体转化至农杆菌GV3101中,采用农杆菌介导法转化84K杨。对鉴定为阳性的转基因植株及84K杨对照植株的相同组织取材进行RNA提取、反转录及实时荧光定量PCR分析表达量变化。以84K杨植株表达量为对照计算转基因植株相对表达量。
-
选取相对表达量较高的株系,组培苗扩繁至每个株系至少15棵重复。生长约4周后选取生长状态良好且长势一致的植株,同具有相同遗传背景的84K杨对照植株一起移栽至营养土中。对移栽后的植株进行表型观察和记录,测定株高和地径。切取新鲜杨树茎段,用速干强力胶固定在样品架上,使用Leica VT1200S振动式切片机,将茎段横切成厚度为40 μm的薄片。切片平铺在载玻片上,滴加质量分数为0.1%的甲苯胺蓝(TBO)染液,染色30~60 s后洗去染液,需根据染色程度调整染色时间。将染色后的切片放在载玻片上,用Leica DM6 B正置荧光显微镜观察并拍照记录,目镜放大倍数为10倍,物镜放大倍数为10倍。使用ImageJ软件测量木质部宽度。
-
用Excel软件进行数据统计,并对株高、地径和木质部宽度进行显著性分析。
-
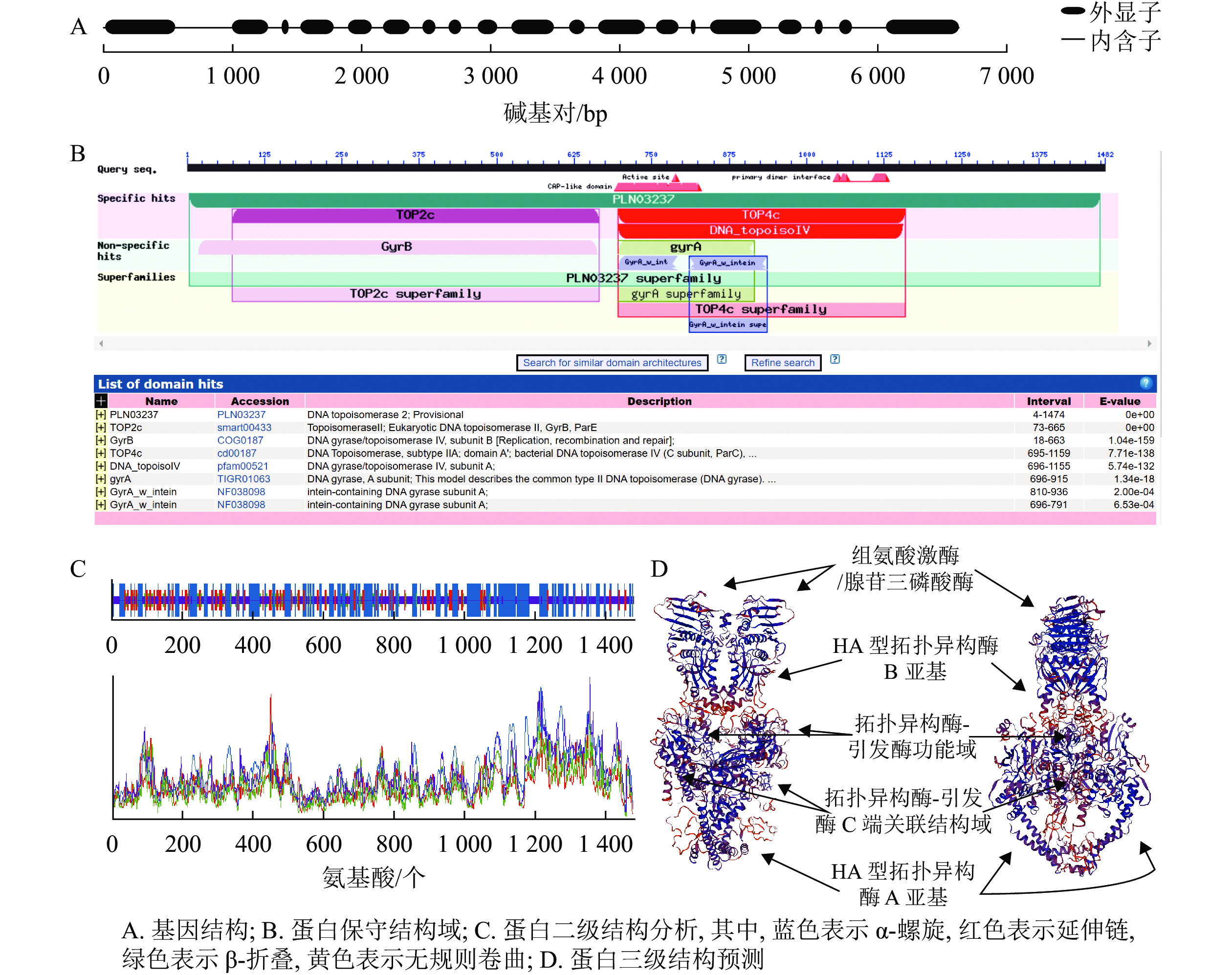
以84K杨cDNA为模板,克隆得到PagTOP2b基因全长序列,对其进行基因结构、蛋白理化性质、保守结构域及蛋白高级结构的分析。如图1A所示:PagTOP2b基因结构包含19个外显子,蛋白质编码区(CDS)长度为4 449 bp。如图1B所示:蛋白保守结构域主要包括PLN03237,目前被认为是跨越多个结构域的DNA拓扑异构酶Ⅱ模块,此外特定匹配的保守结构域还包括靠近N端的TOP2c,是真核生物中Ⅱ型拓扑异构酶具有的结构域;位于中部靠近C端的TOP4c,是ⅡA亚型拓扑异构酶具有的结构域。如图1C所示:蛋白二级结构中,α-螺旋占40.08%,延伸链占14.51%,β-折叠占5.20%,无规则卷曲占40.22%,即蛋白主要折叠方式为α-螺旋和无规则卷曲。如图1D所示:蛋白三级结构为同型二聚体,包含组氨酸激酶/腺苷三磷酸酶结构域,2个ⅡA型拓扑异构酶功能域,以及拓扑异构酶-引发酶功能域(topoisomerase-primase domain,TOPRIM domain),该结构域是在Ⅱ型拓扑异构酶等酶类中发现的核苷酸转移酶/水解酶功能域。

图 1 PagTOP2b基因及其编码蛋白的特征分析
Figure 1. Characteristics of PagTOP2b gene and its coding protein
蛋白理化性质分析结果表明:PagTOP2b蛋白共有1 482个氨基酸,蛋白分子质量为166.1 kDa。理论等电点为7.54,属碱性蛋白。亲水性平均系数为负数,表明蛋白均具有较强的亲水性。脂肪族氨基酸指数(蛋白中脂肪族侧链所占据的相对体积)为77.76,不稳定指数为41.04,大于40,为不稳定蛋白。亚细胞定位预测结果显示:PagTOP2b蛋白具有叶绿体和细胞核定位信号。
-
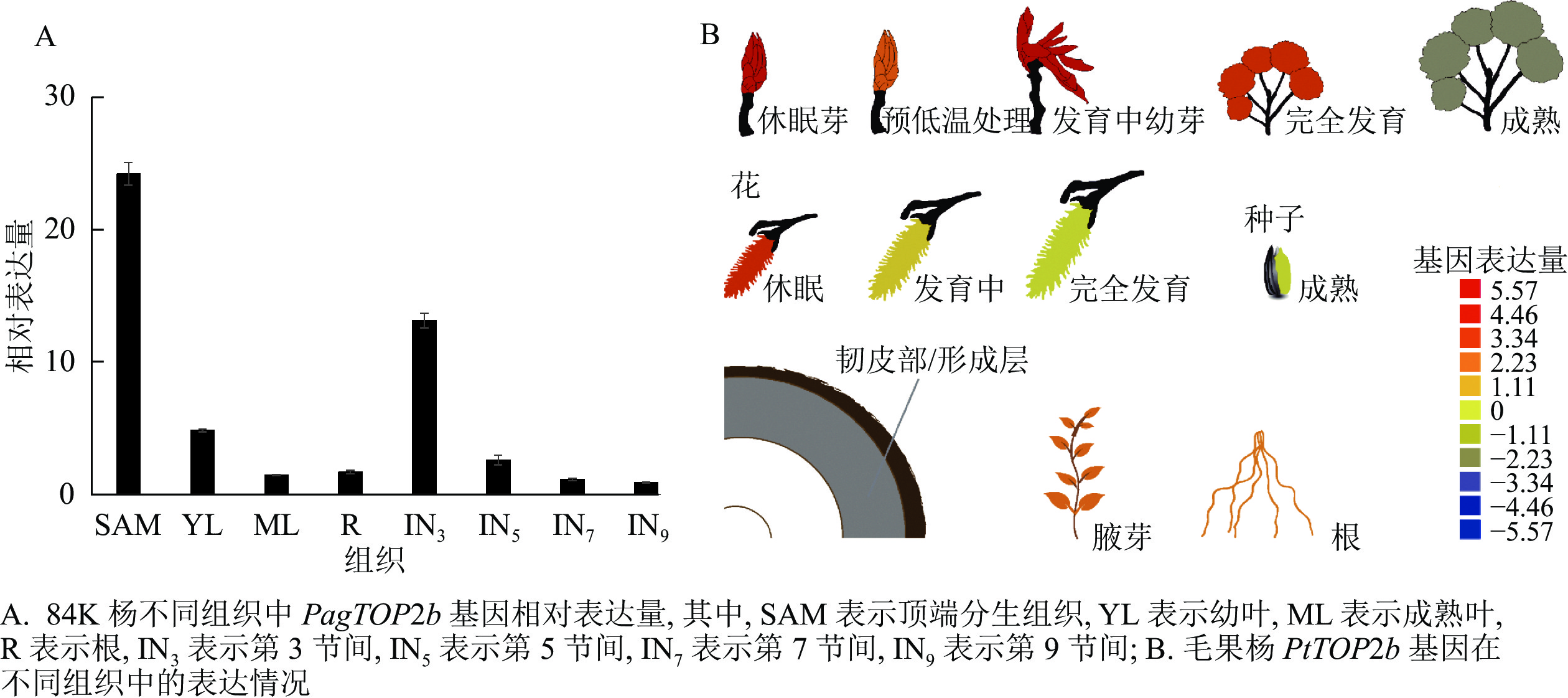
为研究PagTOP2b基因在84K杨不同组织的表达情况,分别取84K杨顶芽,第3、5、7、9节间,幼叶,成熟叶和根进行RNA提取,反转录合成cDNA后进行实时荧光定量分析。结果如图2A所示:PagTOP2b在茎顶端分生组织(SAM)表达量最高,其次在第3节间(IN3)表达量较高,在幼叶(YL)中的表达量也高于在成熟叶(ML),即该基因在84K杨的幼嫩组织中有较高的表达量。同时在杨树资源数据库网站PopGenIE 中,查找毛果杨P. trichocarpa中Ⅱ类TOP基因PtTOP2b在杨树不同组织中的表达情况,如图2B所示:PtTOP2b在休眠的叶芽、发育过程中的叶芽以及休眠的花芽中有较高的表达量,与84K杨中定量分析结果相一致。
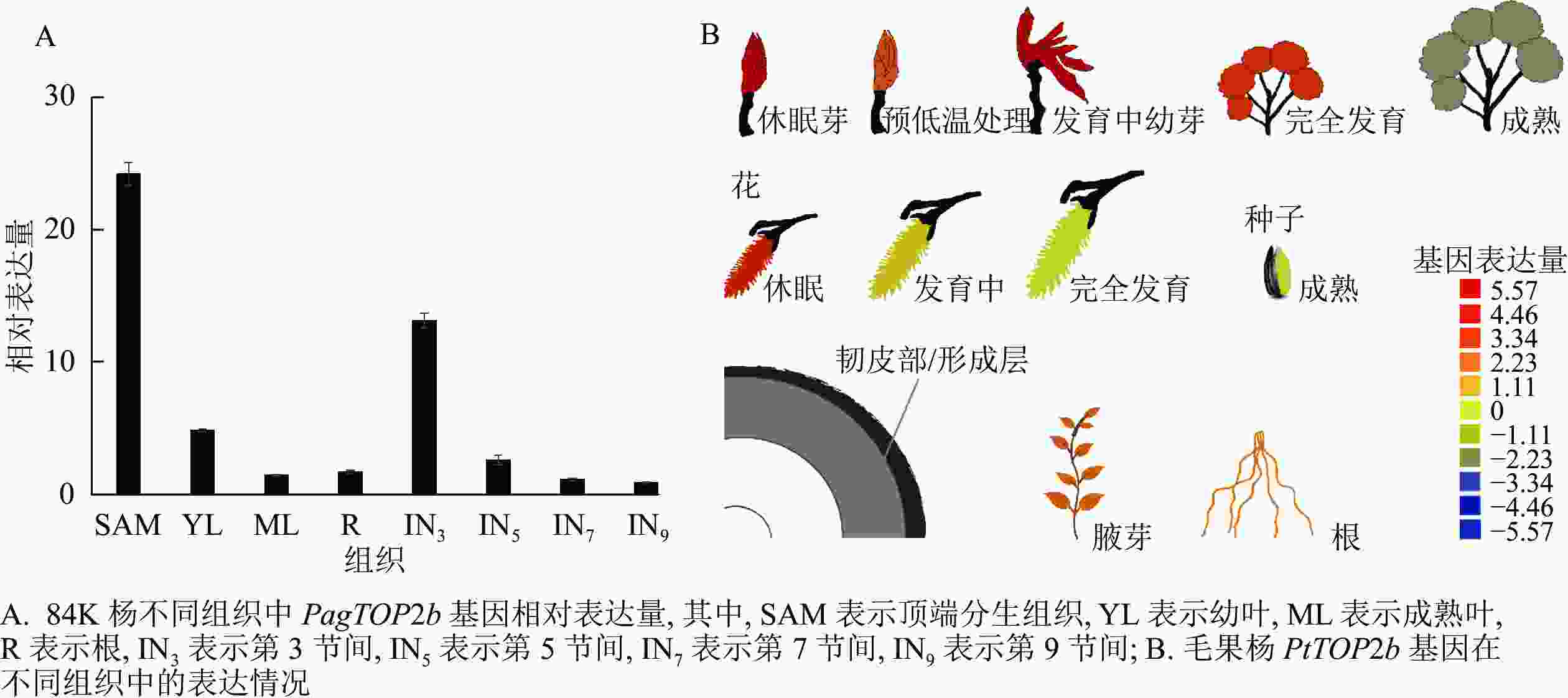
图 2 TOP2b基因在毛果杨和84K杨不同组织中的表达情况
Figure 2. Expression of TOP2b gene in different tissues of black cottonwood and poplar ‘84K’
-
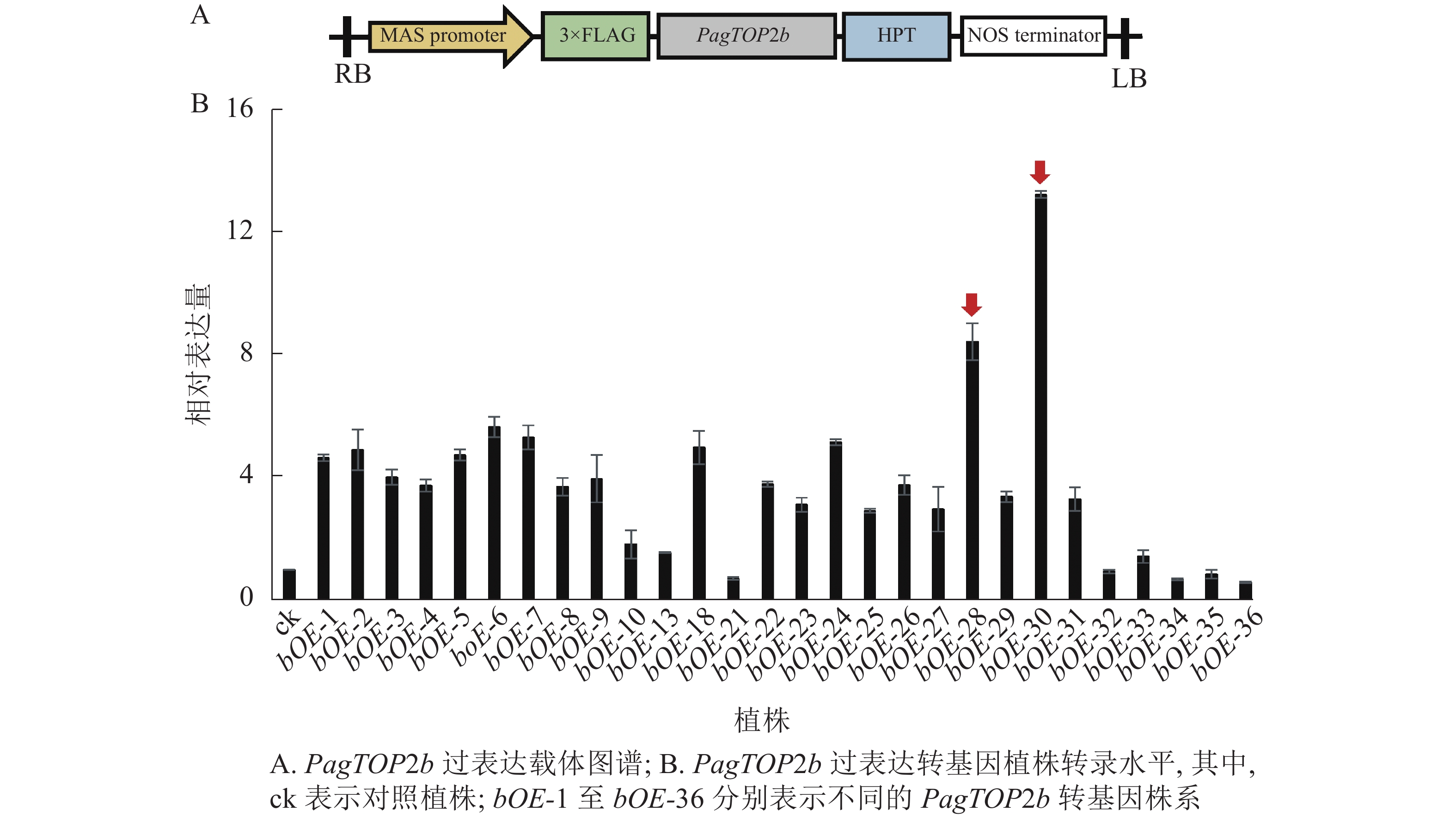
为进一步研究PagTOP2b基因对杨树生长发育的影响,通过农杆菌介导的叶盘转化法获得PagTOP2b过表达的转基因84K杨植株。过表达载体图谱见图3A,对转基因植株进行转录水平分析,选择表达量较高的株系用于后续试验。根据目的基因的相对表达量情况(图3B),选择PagTOP2b过表达株系bOE-28和bOE-30进行后续实验。过表达株系bOE-28中目的基因相对表达量为对照植株中目的基因表达量的8.46倍,bOE-30为13.29倍。
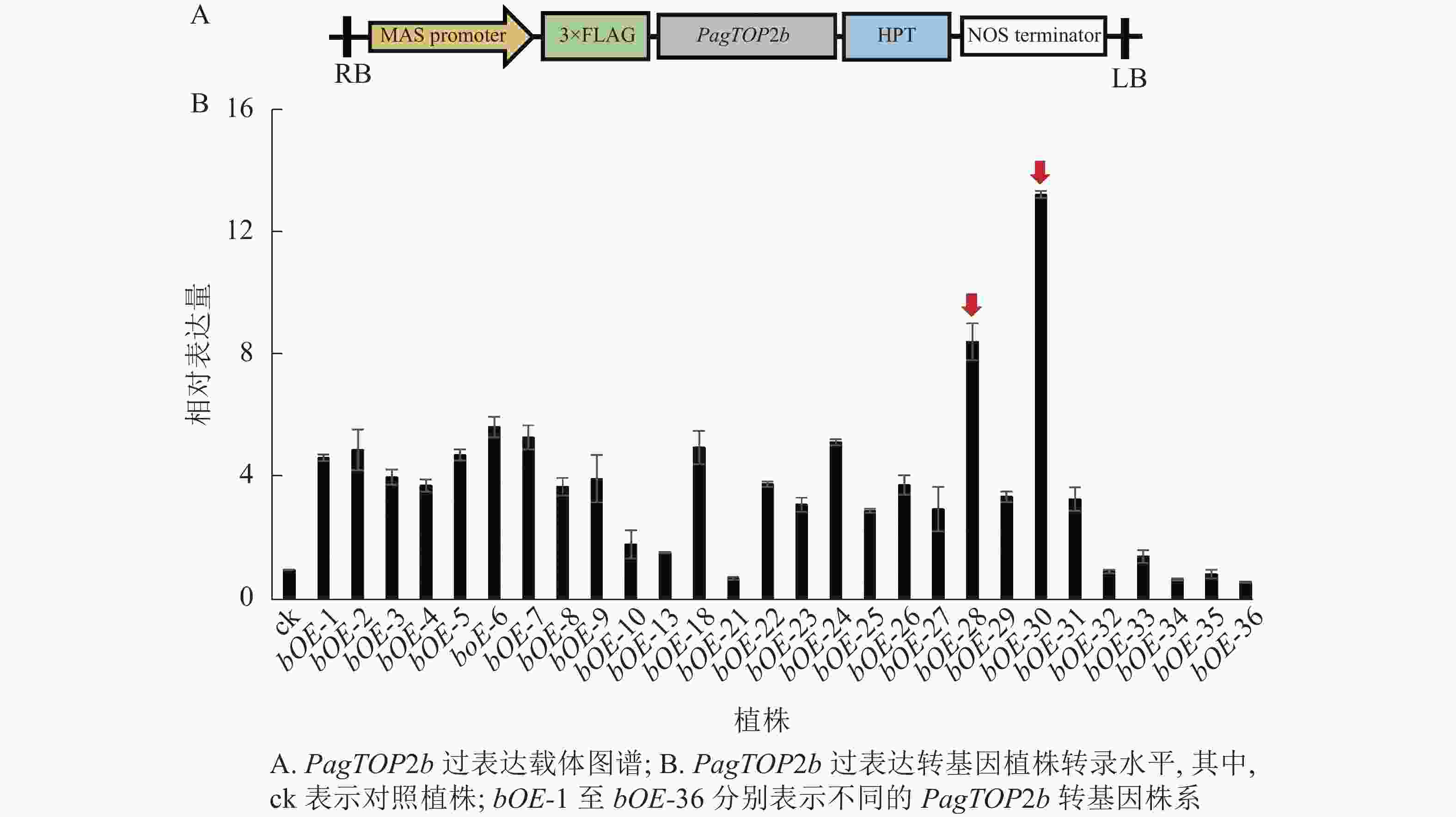
图 3 PagTOP2b过表达载体及转基因植株转录水平分析
Figure 3. PagTOP2b overexpression vector and transcriptional level analysis of transgenic plants
-
将过表达植株和对照植株移栽至土壤培养3个月后观察并记录表型(图4A)。取第7、9、11节间并对其横切面进行切片观察,发现对照与过表达植株相比细胞形态无明显差异(图4B)。相比于对照,PagTOP2b过表达株系bOE-28和bOE-30植株高度显著大于对照(P<0.05),相比对照分别增加了17.5%和11.5%(图4C);过表达株系bOE-28植株的地径显著大于对照(P<0.05),bOE-30植株的地径极显著大于对照(P<0.01),相比对照分别增加了10.7%和16.3%(图4D)。通过统计其木质部宽度发现:PagTOP2b过表达株系bOE-30自第7节间起木质部宽度极显著大于对照(P<0.01),过表达株系bOE-28第11节间木质部宽度极显著大于对照(P<0.01)(图4E)。

图 4 PagTOP2b过表达转基因杨树表型分析
Figure 4. Phenotypic analysis of PagTOP2b overexpression transgenic poplar
-
植物生长发育的过程,就是植物细胞、组织和器官在数量上不可逆增加以及细胞分化的过程。DNA拓扑异构酶在染色体复制、细胞分裂及基因转录中发挥重要作用。本研究对84K杨中的DNA拓扑异构酶基因PagTOP2b进行克隆及序列分析,对序列长度、保守结构域及蛋白三级结构注释信息的分析显示:该基因包含腺苷三磷酸酶结构域、TOPRIM结构域以及ⅡA亚型拓扑异构酶具有的结构域,与先前研究中总结的动植物ⅡA亚型拓扑异构酶结构相符[5, 28],因此判断该基因属于ⅡA亚型DNA拓扑异构酶。
对PagTOP2b基因在84K杨不同组织中的表达模式分析表明:该基因在顶端分生组织和第3节间中表达量较高。相比于84K杨,毛果杨具有更丰富的转录组数据,能够对基因在杨树不同组织的表达情况进行详细分析,同时对84K杨中DNA拓扑异构酶基因的表达情况而言具有一定借鉴意义。因此,对毛果杨中直系同源基因PtTOP2b的表达模式进行分析显示:PtTOP2b在叶芽及花芽中具有较高的表达量,即在幼嫩组织具有较高的表达量。有研究表明:豌豆Pisum sativum、玉米Zea mays和拟南芥中发现Ⅱ型DNA拓扑异构酶在生长活跃的植物组织中有较高的表达量[15]。本研究结果与前人研究结果一致,说明PagTOP2b基因主要在细胞活跃分裂、分化的组织中表达,推测其在植物的生长发育中起重要作用。同时,PagTOP2b在杨树的第3节间高表达,表明其可能在杨树的径向生长中发挥作用。因此,需要进一步研究确认PagTOP2b对杨树树高生长和径向生长的影响。
为了进一步确定该基因的作用,采用农杆菌介导的基因转化方法,获得了PagTOP2b基因过表达84K杨植株。与对照植株相比,转基因植株高度显著增加,即促进了杨树生物量的增加,说明该基因具有进行定向分子育种的潜在应用价值。除了促进杨树树高生长外,茎段的直径及木质部宽度的增加,说明该基因还影响了杨树木质部的发育,可能来源于对形成层活动的调控,需要进一步的研究予以证实。有研究表明:过表达Ⅱ型DNA拓扑异构酶NtTopoⅡα会对烟草生长发育及植株形态产生影响,相比于野生型,过表达植株开花结籽提前2~3周,茎长增加但直径减小,节间长度变长,根系更加发达[18]。本研究与其在株高增加上的表型一致,但茎直径变化不同,可能由于草本植物与木本植物的茎段次生生长存在差异。总而言之,PagTOP2b基因能够同时促进杨树径向和纵向的生长。
-
本研究通过克隆及序列分析确定PagTOP2b是84K杨中的一个ⅡA型DNA拓扑异构酶基因,在幼嫩组织具有较高的表达量。通过转基因技术过表达PagTOP2b基因,能够显著促进植株增高,茎直径增大,促进杨树径向及纵向的生长,增加植株生物量。
Effects of DNA topoisomerase PagTOP2b on growth and development of poplar ‘84K’
-
摘要:
目的 DNA拓扑异构酶基因是一类能够改变DNA拓扑结构的酶,在基础生命活动如DNA复制、转录、有丝分裂等过程中发挥重要作用。研究DNA拓扑异构酶基因对杨树Populus生长发育的影响,能够进一步了解DNA拓扑异构酶基因在木本植物中的作用机制,评估其是否可作为林木分子育种的靶标。 方法 从银腺杨‘84K’ Populus alba × P. glandulosa ‘84K’ (84K杨)中克隆得到DNA拓扑异构酶基因PagTOP2b,利用生物信息学方法对其进行序列分析、蛋白理化性质及结构分析。实时荧光定量PCR分析PagTOP2b在杨树不同组织中的表达水平。利用农杆菌Agrobacterium tumefaciens介导的叶盘转化法转化84K杨,得到PagTOP2b过表达转基因植株。通过表型及茎段切片分析过表达PagTOP2b基因对84K杨植株生长的影响。 结果 以84K杨cDNA为模板,克隆得到PagTOP2b基因全长序列,该基因蛋白质编码区(CDS)长度为4 449 bp,编码1 482个氨基酸,根据蛋白结构域分析属于ⅡA型DNA拓扑异构酶。PagTOP2b主要在杨树幼嫩组织中有较高的表达量。过量表达PagTOP2b基因会导致转基因植株高度、基部节间直径和木质部宽度显著增加。 结论 DNA拓扑异构酶作为一类参与基础生命活动的重要功能酶,在杨树中过量表达ⅡA型DNA拓扑异构酶基因PagTOP2b可促进植株的纵向及径向生长,增加植株生物量。图4表1参28 Abstract:Objective DNA Topoisomerase, as a kind of enzyme that can change the topological structure of DNA, plays an important role in fundamental cellular activities such as DNA replication, transcription and mitosis, therefore, this study, with an investigtion into the effects of DNA topoisomerase genes on the growth and development of poplar, is aimed to lay a solid foundation for further understanding of the mechanism of DNA topoisomerase in woody plants and to evaluate whether it can be used as a target for molecular breeding. Method First, DNA topoisomerase gene PagTOP2b was cloned from poplar ‘84K’ (Populus alba × P. glandulosa ‘84K’) before an analysis was conducted of its sequence and the protein physicochemical properties and structure with the employment of Bioinformatic method. Then, with an exploration of the expression levels in different tissues of poplar by real-time quantitative PCR, Agrobacterium tumefaciens-mediated leaf disc transformation in poplar ‘84K’ was adopted to obtain PagTOP2b-overexpression transgenic plants with observations made of the effects of overexpression of PagTOP2b gene on the growth and development of plants employing a phenotypic and anatomic analysis. Result The coding sequence (CDS) length of the PagTOP2b gene cloned by using poplar ‘84K’ cDNA as the template was 4 449 bp, encoding 1 482 amino acids. According to protein domain analysis, it is classified as type ⅡA DNA topoisomerase. PagTOP2b was mainly expressed in immature and younger tissues of poplar. Overexpression of PagTOP2b gene resulted in a significant increase in height, basal internode diameter and xylem width of transgenic plants. Conclusion As an important functional enzyme involved in basic life activities, the overexpression of type ⅡA DNA topoisomerase gene PagTOP2b in poplar can significantly promote the longitudinal and radial growth in plant, thus increasing plant biomass. [Ch, 4 fig. 1 tab. 28 ref.] -
Key words:
- DNA topoisomerase /
- Populus alba × P. glandulosa ‘84K’ /
- overexpression /
- biomass
-
图 1 PagTOP2b基因及其编码蛋白的特征分析
Figure 1 Characteristics of PagTOP2b gene and its coding protein
图 2 TOP2b基因在毛果杨和84K杨不同组织中的表达情况
Figure 2 Expression of TOP2b gene in different tissues of black cottonwood and poplar ‘84K’
图 3 PagTOP2b过表达载体及转基因植株转录水平分析
Figure 3 PagTOP2b overexpression vector and transcriptional level analysis of transgenic plants
图 4 PagTOP2b过表达转基因杨树表型分析
Figure 4 Phenotypic analysis of PagTOP2b overexpression transgenic poplar
表 1 基因克隆及实时荧光定量PCR (RT-qPCR)引物
Table 1. Primers for gene clone and RT-qPCR
引物名称 引物序列(5′→3′) PagTOP2b F: ATGGCGGCAGAGACCAAGAAAAT R: GTCAGAGGAAGAGTCAGAATCATCAT PagTOP2b qrtF: AGACTACTGGTGGGAGAAAT qrtR: GTCACCTTGGTCCAATTCTC ACTIN qrtF: AAACTGTAATGGTCCTCCCTCCG qrtR: GCATCATCACAATCACTCTCCGA 说明:F表示基因克隆的正向引物;R表示基因克隆的反向引物;qrtF表示RT-qPCR的正向引物;qrtR表示RT-qPCR的反向引物  下载: 导出CSV
下载: 导出CSV
-
[1] DURAND-DUBIEF M, SVENSSON J P, PERSSON J, et al. Topoisomerases, chromatin and transcription termination [J]. Transcription, 2011, 2(2): 66 − 70. [2] OSHEROFF N, ASHLEY R E. Regulation of DNA topology by topoisomerases: mathematics at the molecular level[M]. ADAMS C, GORDON C, JONES V, et al. Knots Low-Dimensional Topology and Applications. Cham: Springer International Publishing, 2019: 411 − 433. [3] WANG J C. Interaction between DNA and an Escherichia coli protein ω [J]. J Mol Biol, 1971, 55(3): 523 − 533. [4] KRESGE N, SIMONI R D, HILL R. Unwinding the DNA topoisomerase story: the work of James C. Wang[J/OL]. J Biol Chem, 2007, 282(22): e17-e19[2022-04-20]. doi: 10.1016/S0021-9258(20)64989-4. [5] CHAMPOUX J J. DNA topoisomerases: structure, function, and mechanism [J]. Annu Rev Biochem, 2001, 70: 369 − 413. [6] NITISS J L. DNA topoisomerase Ⅱ and its growing repertoire of biological functions [J]. Nat Rev Cancer, 2009, 9(5): 327 − 337. [7] CHEN S H, CHAN N L, HSIEH T S. New mechanistic and functional insights into DNA topoisomerases [J]. Annu Rev Biochem, 2013, 82: 139 − 170. [8] AUSTIN C A, LEE K C, SWAN R L, et al. TOP2B: the first thirty years[J/OL]. Int J Mol Sci, 2018, 19(9): 2765[2022-04-25]. doi: 10.3390/ijms19092765. [9] DELGADO J L, HSIEH C M, CHAN N L, et al. Topoisomerases as anticancer targets [J]. Biochem J, 2018, 475(2): 373 − 398. [10] HEVENER K, VERSTAK T A, LUTAT K E, et al. Recent developments in topoisomerase-targeted cancer chemotherapy [J]. Acta Pharm Sin B, 2018, 8(6): 844 − 861. [11] LIANG Xiaoxia, WU Qiang, LUAN Shangxian, et al. A comprehensive review of topoisomerase inhibitors as anticancer agents in the past decade [J]. Eur J Med Chem, 2019, 171: 129 − 168. [12] RUDENKO G N. Plant DNA-topoisomerase type Ⅱ: isolation, characterization, and properties [J]. Mol Biol, 1992, 25: 887 − 896. [13] FUKATA H, OHGAMI K, FUKASAWA H. Isolation and characterization of DNA topoisomerase Ⅱ from cauliflower inflorescences [J]. Plant Mol Biol, 1986, 6(3): 137 − 44. [14] XIE Shaoping, LAM E. Characterization of a DNA topoisomerase Ⅱ cDNA from Arabidopsis thaliana [J]. Plant Physiol, 1994, 106(4): 1701 − 1702. [15] MAKAREVITCH I, SOMERS D A. Purification and characterization of topoisomerase ⅡA from Arabidopsis thaliana [J]. Plant Sci, 2005, 168(4): 1023 − 1033. [16] XIE Shaoping, LAM E. Abundance of nuclear DNA topoisomerase Ⅱ is correlated with proliferation in Arabidopsis thaliana [J]. Nucleic Acids Res, 1994, 22(25): 5729 − 5736. [17] DYNAN W S, JENDRISAK J J, HAGER D A, et al. Purification and characterization of wheat germ DNA topoisomerase Ⅰ (nicking-closing enzyme) [J]. J Biol Chem, 1981, 256(11): 5860 − 5865. [18] JOHN R, GANESHAN U, SINGH B N, et al. Over-expression of topoisomerase Ⅱ enhances salt stress tolerance in tobacco[J/OL]. Front Plant Sci, 2016, 7: 1280[2022-05-01]. doi: 10.3389/fpls.2016.01280. [19] MARTINEZ-GARCIA M, SCHUBERT V, OSMAN K, et al. TOPⅡ and chromosome movement help remove interlocks between entangled chromosomes during meiosis [J]. J Cell Biol, 2018, 217(12): 4070 − 4079. [20] MARTINEZ-GARCIA M, WHITE C I, FRANKLIN F C H, et al. The role of topoisomerase Ⅱ in DNA repair and recombination in Arabidopsis thaliana[J/OL]. Int J Mol Sci, 2021, 22(23): 13115[2022-05-04]. doi: 10.3390/ijms222313115. [21] PARRA-NUNEZ P, COOPER C, SANCHEZ-MORAN E. The role of DNA topoisomerase binding protein 1 (TopBP1) in genome stability in Arabidopsis[J/OL]. Plants, 2021, 10(12): 2568[2022-05-08]. doi: 10.3390/plants10122568. [22] QIU Deyou, BAI Shenglong, MA Jianchao, et al. The genome of Populus alba×Populus tremula var. glandulosa clone 84K [J]. DNA Res, 2019, 26(5): 423 − 431. [23] CHEN Chengjie, CHEN Hao, ZHANG Yi, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data [J]. Mol Plant, 2020, 13(8): 1194 − 1202. [24] LU Shennan, WANG Jiyao, CHITSAZ F, et al. CDD/SPARCLE: the conserved domain database in 2020 [J/OL]. Nucleic Acids Res, 2020, 48(D1): D265-D268[2022-05-01]. doi: 10.1093/nar/gkz991. [25] WATERHOUSE A, BERTONI M, BIENERT S, et al. SWISS-MODEL: homology modelling of protein structures and complexes[J/OL]. Nucleic Acids Res, 2018, 46(W1): W296-W303[2022-05-02]. doi: 10.1093/nar/gky427. [26] WILKINS M R, GASTEIGER E, BAIROCH A, et al. Protein identification and analysis tools in the ExPASy server [J]. Methods Mol Biol, 1999, 112: 531 − 552. [27] CHOU K C, SHEN H B. Cell-PLoc 2.0: an improved package of web-servers for predicting subcellular localization of proteins in various organisms [J]. Nat Sci, 2010, 2(10): 1090 − 1103. [28] SINGH B N, SOPORY S K, REDDY M K. Plant DNA topoisomerases: structure, function, and cellular roles in plant development [J]. Crit Rev Plant Sci, 2004, 23(3): 251 − 269. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220372
 点击查看大图
点击查看大图
计量
- 文章访问数: 2140
- HTML全文浏览量: 533
- PDF下载量: 144
- 被引次数: 0

