




-
生物多样性是人类赖以生存及经济发展的基础[1-2]。然而,随着经济的发展,高强度的森林采伐,导致生物多样性减少,水资源和非木质产品不断损失[3]。生物多样性的损失或许是影响人类生存最重要的因素。林分结构是林分特征中最重要的内容,林分结构的研究对充分发挥森林多项功能,科学经营森林具有重大的意义。因此,开展人为干扰下林分结构及生物多样性的研究具有重要的理论和现实意义。干扰是自然界中普遍存在的、不同时空尺度上偶然发生的不可预知的事件,是自然生态系统演替过程中一个重要的组成部分,直接影响着森林物种组成、群落结构和生物多样性。不同干扰类型和强度对森林群落结构特征有着显著的影响。随着人类活动的加剧,人为干扰对森林生态系统的影响也日益受到重视。国内外关于人为干扰对森林影响做过大量研究,主要集中于物种组成和多样性研究[4-12],并认为中度干扰强度下的物种多样性有增加的趋势,即“中度干扰假说”,然而干扰与生物多样性的关系非常紧密,也非常复杂[13],并不是所有的干扰对物种多样性的影响都符合该假说所描述的规律[14-18]。常绿阔叶次生林作为中国南方典型的地带性植被类型,具有显著的水源涵养、多样性保护等多项生态功能[19-20]。然而,自20世纪50年代以来,由于森林资源不合理开发利用,人为干扰加剧,南方大部分常绿阔叶林不断减少,退化严重,生态环境问题日益突出。目前,国内外关于常绿阔叶次生林的研究主要集中于林分结构[21-24]、更新演替[25]、森林功能[19, 26-27]、群落特征[28-30]、更新评价[31]、健康评价[32-33]和经营模式[34-36]等方面,有关人为干扰对林分结构影响研究较少。本研究通过对不同干扰等级下常绿阔叶次生林林分结构及多样性变化的研究,探讨林分结构及多样性变化规律,了解人类活动对其的影响,以期寻求合理的森林经营模式,为常绿阔叶次生林的植被恢复和生物多样性保护提供科学依据。
-
福建省将乐林场位于福建省三明市西北部将乐县城区,处武夷山脉东南麓,扼闽江支流金溪中下游,介于26°26′~27°04′N,117°05′~117°40′E,东临顺昌,西接泰宁,南连明溪,北毗邵武,东南与沙县接壤,境内山峰林立,溪流密布,最高峰海拔为1 640.2 m。将乐林场土地总面积为7 113.6 hm2,森林覆盖率为94.8%,有林地面积为6 568.9 hm2。将乐林场属亚热带季风气候,具有海洋性和大陆性气候特点,年平均气温为18.7 ℃,年平均降水量为1 703.7 mm,最高年度为2 460.4 mm,最低年度仅1 007.8 mm,年际差异较大。将乐林场土壤以红壤分布面积最大,土壤质地一般为沙壤土或轻壤土,块状结构,紧实度适中,比较潮湿。
将乐林场常绿阔叶次生林主要树种为栲树Castanopsis fargesii,赤杨叶Alniphyllum fortune,木荷Schima superba,甜槠Castanopsis eyrei,米槠Castanopsis carlesii,青冈Cyclobalanopsis glauca,枫香Liquidamba formosana,苦槠Castanopsis sclerophylla,南酸枣Choerospondias axillaria,黄樟Cinnamomum porrectum,冬青Ilex chinensis,光叶山矾Symplocos lancifolia,杨梅Myrica rubra,檵木Loropetalum chinense,杉木Cunninghamia lanceolata,檫木Sassafras tzumu,山矾Symplocos sumuntia等。
-
在福建将乐县林场全面踏查的前提下,采用典型样地调查的方法,选取具有代表性地段设置样地25块。这些林分均是1958年皆伐后自然生长起来的常绿阔叶次生林。本研究仅局限于人为干扰,不包括自然干扰。样地形状为矩形,面积为0.04~0.12 hm2,记录样地的基本情况,包括经纬度、海拔、坡度、坡向等,并画出样地的方位示意图。采用相邻格子法将每块样地分成10 m × 10 m的调查单元,在每个调查单元内进行每木检尺;在样地的中心和四角分别设置5个5 m × 5 m小样方,调查并记录样方内幼苗幼树的树种、直径(树高>1.3 m的测胸径,树高<1.3 m的只测地径)、树高、株数等。样地基本情况见表 1。
表 1 样地基本情况表
Table 1. Bazis zituation of ztand
样地号 坡度/(°) E N 面积/hm2 郁闭郁闭 平均胸径/cm 平均树高/m 树种组成 1 40° 129°02, 33.72" 46°41, 50.22" 0.06 0.7 19.0 9.7 5米槠3栲树2苦槠+檫木+木荷 2 44° 129°02'33.72" 46°41'50.22" 0.06 0.7 18.0 9.3 5苦槠1栲树1米槠1油桐1山矾1赤杨叶+茅栗+檫木+福建 3 40° 117°26'06.72" 26°42' 15.18" 0.06 0.7 13.9 8.1 5赤杨叶2枫香1檵木1栲树1苦槠+黄樟+天仙果 4 25° 117°26'06.78" 26°43'40.80" 0.06 0.8 12.5 11.3 4栲树2赤杨叶1枫香1苦槠1木荷1南酸枣+黄樟+天仙果 5 23° 117°24'43.44" 26°47'36.30" 0.06 0.8 13.5 9.8 3栲树2苦槠1赤杨叶1青冈1杉木1木荷1南酸枣 6 23° 117°24'44.10" 26°47'37.32" 0.06 0.8 10.8 8.8 4赤杨叶3栲树1檫木1木荷1杉木+苦槠+光叶山矾+青冈+黄樟+冬青 7 38° 117°24'44.76" 26°47'36.24" 0.06 0.6 15.3 7.0 6栲树1木荷1苦槠1杨梅1青冈+南酸枣+台湾冬青+檵木+刨花润楠 8 37° 117°21'57.30" 26°47'5.40" 0.06 0.5 19.4 9.5 7栲树2青冈1甜槠+细柄阿丁枫+枫香+苦槠 9 34° 117°21'57.30" 26°47'5.40" 0.06 0.6 16.4 10.2 5栲树2青冈1南酸枣1山矾1木荷+米槠+苦槠 10 36° 117°22' 13.26" 26°46'54.42" 0.06 0.6 13.7 9.9 4栲树2木荷1黄樟1甜槠1青冈1黄毛润楠+米槠+山矾+南酸枣 11 35° 117°24'51.30" 26°47'51.90" 0.06 0.8 12.5 9.8 3栲树2赤杨叶2臭椿1木荷1杨桐1苦槠+黄樟+冬青+南酸枣 12 31° 117°24'07.26" 26°47'51.54" 0.06 0.6 15.8 8.4 8赤杨叶1黄樟1檵木+椤木石楠+天仙果+猴欢喜+枫香 13 31° 117°26'08.46" 26°43'39.30" 0.06 0.5 14.9 7.9 5苦槠1栲树1米槠1油桐1山矾1赤杨叶+茅栗+檫木+福建青冈 14 25° 117°24'41.46" 26°47'35.16" 0.12 0.7 12.2 9.2 4栲树2赤杨叶1杉木1南酸枣1木荷1苦槠+马尾松+木油桐+光叶山矾+杨梅 15 40° 117°22'20.22" 26°46'53.70" 0.06 0.6 6.9 6.1 4赤杨叶2刨花润楠1山乌桕1樟树1绒毛山胡椒1栲树 16 39° 117°22' 14.88" 26°46'47.98" 0.06 0.6 9.3 8.3 3栲树2柿1杉木1滇青冈1赤杨叶1浙闽樱 17 38° 117°22'14.40" 26°46'49.32" 0.06 0.6 7.9 6.8 5苦槠2栲树1山乌桕1红楠 18 37° 117°22'16.35" 26°46'50.51" 0.06 0.6 13.5 8.8 3苦槠2栲树2马尾松1山乌桕1枫香1红楠 19 24° 117°25'23.29" 26°47'36.01" 0.06 0.9 11.9 11.6 2栲树2苦槠1木荷1刨花润楠1吊皮锥1乐昌含笑1红楠 20 24° 117°25'23.40" 26°47'36.78" 0.04 0.9 11.5 12.4 4栲树1木蜡树1细柄阿丁枫1吊皮锥1红楠1木荷 21 36° 117°24'52.74" 26°47'56.10" 0.06 0.8 11.6 9.7 5栲树2黄樟1刺毛杜鹃1苦槠+鼠刺 22 41° 117°24'52.02" 26°47'50.92" 0.06 0.8 10.3 9.7 5栲树2木荷1苦槠1罗浮锥 23 31° 117°19'19.00" 26°31'28.00" 0.06 0.6 17.4 10.4 5甜槠2红楠1细齿叶柃1福建冬青 24 32° 117°18'21.00" 26°31'22.00" 0.06 0.8 13.7 10.6 4栲树3甜槠2石木姜子 25 30° 117°15'33.00" 26°32'36.00" 0.04 0.7 14.5 11.0 4甜槠3栲树1木荷1苦槠+米槠+刨花润楠 说明:油桐Vernicia fordii; 茅栗Castanea seguinii; 福建青冈Cyclobalanopsis chungii; 天仙果Ficuss beecheyana; 台湾冬青Iles formosana; 刨花润楠Machilus pauhoi; 细柄阿丁枫Altingia angustifolia; 黄毛润楠Machilus chrysotricha; 臭椿Ailanthus altissima; 杨桐Adinandra millettii; 椤木石楠Photinia davidsoniae; 猴欢喜Sloanea sinensis; 马尾松Pinus massoniana; 木油桐Vernicia montana; 山乌桕Sapium discolor; 樟树Cinnamomum camphora; 绒毛山胡椒Lindera nacusua; 柿Diospyros kaki; 滇青冈Cyclobalanopsis glaucoides; 浙闽樱(桃)Cerasus schneideriana; 红楠Machilus thunbergii; 吊皮锥Castanopsis karakamii; 乐昌含笑Michelia chapensis; 木蜡树Tosicodendron sylvestre; 刺毛杜鹃Rhododendron championae; 鼠刺Itea chinensis; 罗浮锥Castanopsis fabri; 细齿叶柃Eurya nitida; 福建冬青Iles fukienensis; 石木姜子Litsea elongata -
在调查样地的同时,对每个样地的干扰情况进行调查分析。根据将乐林场常绿阔叶次生林的实际调查情况,选取相对郁闭度、幼树相对密度、受害木(压弯、折断或掘根)相对密度、受害木相对断面积、伐倒木相对密度和伐倒木相对断面积6个干扰影响因素对常绿阔叶次生林受干扰程度进行定量评价。采用相对影响方法[1],计算6个干扰因素对常绿阔叶次生林干扰程度的相对值。如伐倒木密度中2号样地密度最小,为17株·hm-2,1号,3号和4号样地伐倒木株数分别为50,50和33株·hm-2,则以伐倒木密度最小的2号样地产生干扰的相对值为1,1号、3号和4号样地相对值分别为50/17=2.9,50/17=2.9和33/17=1.9。其余5个干扰影响因素对常绿阔叶次生林产生干扰的相对值依此计算(表 2);然后,将6个干扰因素对常绿阔叶次生林干扰强度的相对值的总和作为人为干扰强度值;最后,参考生态干扰度划分系统[37],结合将乐林场常绿阔叶次生林的实际情况,采用等距法依据常绿阔叶次生林干扰影响因素值即干扰度值(FDI),将25块常绿阔叶次生林样地划分为Ⅰ~Ⅴ等5个干扰等级(表 3)。
表 2 常绿阔叶次生林干扰等级
Table 2. Dizturbanse gradient of evergreen broad-leaved zesondary foreztz
干扰等级 样地号 相对郁度 幼树相对密度 受害木相对密度 受害木相对断面积 伐倒木相对密度 伐倒木相对断面积 总和 干扰等级 样地号 相对郁度 幼树相对密度 受害木相对密度 受害木相对断面积 伐倒木相对密度 伐倒木相对断面积 总和 Ⅰ 1 1.2 6.2 1.0 1.1 3.9 12.2 17.8 Ⅲ 8 1.2 8.5 18.0 10.7 2.9 1.9 52.7 Ⅰ 2 1.2 6.1 1.3 2.1 2.9 1.5 15.1 Ⅲ 9 1.7 3.0 21.7 23.4 2.9 3.9 55.5 Ⅰ 3 1.6 5.1 1.3 1.5 1.0 2.9 25.6 Ⅲ 11 1.6 4.2 18.3 15.6 5.9 13.0 64.0 Ⅰ 4 1.2 8.3 2.0 3.6 3.9 6.9 18.7 Ⅲ 12 1.7 1.7 17.3 21.8 2.9 13.3 61.6 Ⅰ 5 1.4 1.0 6.3 15.3 2.9 1.7 25.9 Ⅲ 15 1.6 9.3 24.3 20.6 4.4 9.9 56.6 Ⅰ 6 1.6 3.4 5.3 3.6 2.9 1.0 13.4 Ⅲ 19 1.2 3.1 1.7 1.0 2.9 8.3 58.7 Ⅰ 7 1.4 1.7 12.7 34.8 1.0 1.1 28.6 Ⅲ 20 1.6 4.9 10.0 24.4 1.9 4.7 58.6 Ⅰ 18 1.2 3.7 5.7 3.0 3.9 1.2 18.2 Ⅳ 16 1.2 4.9 27.3 25.9 2.9 6.2 70.1 Ⅱ 10 1.0 4.2 4.3 4.4 3.9 19.5 37.3 Ⅳ 22 1.6 4.4 4.7 18.9 1.9 8.0 68.4 Ⅱ 13 1.4 3.0 22.0 22.7 2.9 3.5 32.4 Ⅳ 23 1.2 6.7 13.3 20.1 15.7 16.0 73.0 Ⅱ 14 1.2 6.2 16.7 25.9 1.9 9.7 43.2 Ⅴ 24 1.2 5.9 23.0 29.4 8.7 25.8 94.0 Ⅱ 17 1.0 8.7 11.0 8.0 1.9 1.8 47.5 Ⅴ 25 1.4 10.1 43.3 37.4 1.9 7.0 101.1 Ⅱ 21 1.6 4.1 21.7 31.4 1.9 3.3 39.5 表 3 干扰度指数分级
Table 3. Hierarchical list of index of disturbance
干扰等级 干扰度值 Ⅰ 13.4≤FDI < 30.9 Ⅱ 30.9≤FDI < 48.5 Ⅲ 48.5≤FDI < 66.0 Ⅳ 66.0≤FDI < 83.6 Ⅴ 83.6≤FDI < 101.1 -
采用体现树种空间隔离程度的树种混交度(Mi),反映林木个体大小的大小比数(Ui),描述林木个体在水平地面上分布格局的角尺度(Wi)3个参数对林分空间结构进行分析[38]。本研究选定相邻木n=4, 标准角为α0=72°。计算公式如下:
$$ \begin{array}{l} {M_i} = \frac{1}{n}\sum\limits_{j = 1}^n {{V_{ij}}} , {V_{ij}}\left\{ \begin{array}{l} 1, \;参照树i与第j株相邻木为非同种\\ 0, \;否则 \end{array} \right.\;\;\;\;\;\;\;\;\;\;\;\;\;\;\;\;\;\;\;\;\;\;\;\;;\\ {U_i} = \frac{1}{n}\sum\limits_{j = 1}^n {{K_{ij}}} , {K_{ij}}\left\{ \begin{array}{l} 1, \;若相邻木j比参照树i大\\ 0, \;否则 \end{array} \right.\;\;\;\;\;\;\;\;\;\;\;\;\;\;\;\;\;\;;\\ {W_i} = \frac{1}{n}\sum\limits_{j = 1}^n {{Z_{ij}}} , {Z_{ij}}\left\{ \begin{array}{l} 1, \; 当第j个\alpha 角小于标准角{{\alpha }_{0}}\\ 0, \;否则 \end{array} \right.\;\;\;\;\;\;\;\;\;\;\;\;\;\;\;\;\;\;。 \end{array} $$ -
采用物种丰富度,Margalef丰富度指数,Shannon-Wiener指数,Pielou均匀度指数作为物种α多样性测度指标,用Jaccard相似指数来评价β多样性,Simpson优势度指数作为树种优势度测量指标。①物种丰富度:S=出现在样地内的物种数量。②Margalef物种丰富度指数(DM):DM=(S-1)/lnN。其中:S为样方中物种的数目,N为样方中所有物种个体的数目。③Shannon-Wiener指数(H):$H=-\sum\limits_{i=1}^{S}{{{P}_{i}}\ln {{P}_{i}}} $。其中:Pi为物种i的相对重要值,S为样地的物种总数。④Pielou均匀度指数(E):E=H/lnS。其中:H为Shannon-Wiener指数,S为样地的物种总数。⑤Jaccard相似指数(IS):IS=c/(a+b+c)×100。其中:c为2个样方共有的物种数,a为样地1特有的物种数,b为样地2特有的物种数。⑥Simpson优势度指数(C):$C=\sum\limits_{i-1}^{S}{P_{i}^{2}} $。其中:Pi为物种i的相对重要值,S为样地的物种总数。
-
不同干扰等级下常绿阔叶次生林林分结构不同(表 4)。随着干扰等级的增加,林分平均胸径和平均树高逐渐减小。林分平均密度和幼树密度随着干扰等级的增加先增大后减小,均在干扰等级Ⅳ达到最高值,干扰等级Ⅴ与其他干扰等级相比,林分密度和幼树密度均为最小。
表 4 不同干扰等级林分基本情况
Table 4. Basic situation of forest along a disturbance gradient
干扰等级 林分密度/(株·hm-2) 平均胸径/cm 平均树高/m 幼树密度/(株·hm-2) Ⅰ 1 215 14.3 11.7 381 Ⅱ 1 208 14.4 10.5 367 Ⅲ 1 448 13.2 9.8 335 Ⅳ 1 627 12.1 9.1 605 Ⅴ 600 11.8 8.8 100 -
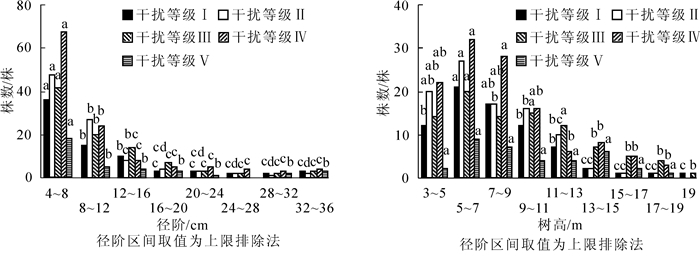
不同干扰等级下常绿阔叶次生林的直径结构差异显著(P<0.05)(图 1A),乔木个体在各径阶的数量分布不同。不同干扰等级的径阶分布均为倒“J”型曲线,即不同干扰等级的直径分布均是随着径级的增大,株数随之减少。在同一干扰等级下,小径阶(DBH<12 cm)个体数显著大于中径阶(12 cm≤DBH<24 cm)和大径阶(DBH≥24 cm)个体数(P<0.05),约占63.89%~78.72%,说明该林分具有潜在的天然更新能力。在同一径阶结构下,干扰等级Ⅴ的小径阶个体数、中径阶个体数和大径阶个体数显著小于其他干扰等级(P<0.05),表明重度干扰等级下,人为的选择性采伐不仅造成大径级树木个体的减少,还因小径阶和中径阶个体数量的减少而降低了林分天然更新能力和生产力。

图 1 不同干扰等级下胸径和树高分布
Figure 1. Diztribution of DBH and tree keigkt along a dizturbanse gradient
干扰等级Ⅰ~Ⅳ常绿阔叶次生林的树高结构差异显著(P<0.05)(图 1B),乔木个体在各高度级的数量分布不同。干扰等级Ⅰ~Ⅳ均为单峰山状曲线,乔木层的高度级峰值均出现在高度级5~7级,并向两侧递减,低高度级(树高<9 m)个体数显著多于中高度级(9 m≤树高<15 m)和高高度级(树高≥15 m)个体数(P<0.05),表明各干扰等级林分垂直结构趋向于稳定。干扰等级Ⅴ各高度级个体数量差异不显著(P>0.05),其分布有2个峰值(5~7级和15~17级),并且缺失最大树高级(高度级19),说明干扰等级Ⅴ树高结构受人为干扰影响较大,林分垂直分布结构不稳定。
-
不同干扰等级的常绿阔叶次生林随着干扰等级的增加,角尺度逐渐增大(表 5)。依据平均角尺度(W)的计算标准,当W>0.517时为团状分布,因此5种干扰类型均属于团状分布,且随着干扰程度的加重,林分越趋向于团状分布。
表 5 不同干扰等级空间结构特征
Table 5. Structural characteristics of spatial structure along a disturbance gradient
干扰等级 角尺度 大小比数 混交度 Ⅰ 0.559 7 0.506 5 0.761 6 Ⅱ 0.564 3 0.500 0 0.737 2 Ⅲ 0.566 4 0.499 0 0.710 7 Ⅳ 0.582 7 0.492 2 0.708 3 Ⅴ 0.590 8 0.485 0 0.676 1 P 0.045 < 0.05 0.039 < 0.05 0.017 < 0.05 不同干扰等级的常绿阔叶次生林随着干扰等级的增加,大小比数逐渐减小。不同干扰等级的常绿阔叶次生林均处于中庸状态,即每株林木的4株相邻木中有一半比参照树的胸径大。
不同干扰等级的常绿阔叶次生林随着干扰等级的增加,混交度逐渐减小。不同干扰等级的常绿阔叶次生林均趋向于强度混交的状态,即每株林木周围均有2~3株其他树种。一般认为,混交度越大,林分相对越稳定[39]。随着干扰等级的增加,林分稳定性不断降低,不利于林分的健康稳定生长。
-
从表 6可以看出:不同干扰等级下的多样性指数存在显著差异。中度干扰林分(干扰等级Ⅱ)具有最高的物种丰富度,重度干扰林分物种丰富度最低。轻度干扰林分的Margalef指数,Shannon-Wiener指数及个体物种数均高于其他干扰等级,重度干扰林分Margalef指数,Shannon-Wiener指数及个体物种数最低,其他干扰强度各指数相差不大。Pielou指数随着干扰强度的增大,指数不断减小。总体来看,随着干扰等级的增大,α多样性指数逐渐减小,轻度干扰和中度干扰林分的α多样性指数均高于重度干扰林分。Simpson优势度指数在不同干扰等级下显著变化,重度干扰森林优势度指数最高,轻度干扰森林优势度指数最低。
表 6 不同干扰等级下α多样性
Table 6. α-diversity along a different disturbance gradient
干扰等级 多样性测量 物种丰富度 特有物种数目 Margalef指数 Shannon-Wiener指数 个体物种数 Pielou指数 Simpson优势度指数 Ⅰ 44 6 4.50 ± 0.49 2.60 ± 0.12 0.24 ± 0.04 1.20 ± 0.08 0.13 ± 0.02 Ⅱ 59 17 3.51 ± 0.19 2.25 ± 0.07 0.20 ± 0.01 0.81 ± 0.02 0.15 ± 0.01 Ⅲ 45 8 3.51 ± 2.22 2.22 ± 0.13 0.22 ± 0.04 0.85 ± 0.02 0.10 ± 0.02 Ⅳ 41 9 3.62 ± 0.75 2.31 ± 0.23 0.19 ± 0.03 0.82 ± 0.03 0.11 ± 0.02 Ⅴ 31 6 3.20 ± 0.39 1.49 ± 1.20 0.12 ± 0.05 0.45 ± 0.33 0.52 ± 0.40 F 3.359 13.316 1.261 24.402 82.596 P 0.029<0.05 0.000<0.05 0.032<0.05 0.000<0.05 0.000<0.05 随着干扰等级提升,Jaccard相似指数呈降低的趋势(表 7),干扰等级Ⅰ,Ⅱ,Ⅲ,Ⅳ与干扰度等级Ⅴ的Jaccard相似指数都偏低,其中轻度干扰和重度干扰之间物种相似性较低,中度干扰林分之间物种相似性最高。说明干扰强度越大,常绿阔叶次生林物种异质性越强,破碎化也越严重。
表 7 不同干扰等级下物种相似值
Table 7. Jaccard's coefficient along a disturbance gradient
干扰等级 Ⅰ Ⅱ Ⅲ Ⅳ Ⅴ Ⅰ 100 45.8 38.5 40.0 27.1 Ⅱ 100 46.6 32.1 28.8 Ⅲ 100 35.4 33.9 Ⅳ 100 34.1 Ⅴ 100 -
干扰是普遍存在于森林生态系统中的正常现象[40-41]。常绿阔叶次生林随着干扰强度的增加,不同干扰等级直径结构和树高结构之间差异显著。不同干扰等级直径分布均趋向于倒“J”型,且随着干扰等级的增大,森林潜在更新能力和生产力皆随之降低。干扰等级Ⅰ~Ⅳ树高分布均为单峰山状曲线,垂直结构较为稳定,干扰等级Ⅴ有2个峰值且各高度级之间个体数差异不显著,垂直结构不稳定。从林分空间结构分析结果来看,随着干扰强度的增加,林分角尺度逐渐增大,林分均属于团状分布;大小比数逐渐减下,林分均处于平庸状态;混交度逐渐减小,林分均趋向于轻度混交的状态。林分受外界干扰的影响,导致幼树幼苗被压、天窗形成、地形破坏、大径级立木缺失等情况发生,改变了原有群落的组织结构,丧失了这些优势木的庇护作用,扰动林内微环境,使林分处于暂时不稳定状态,影响群落的稳定性和演替。这些同时表明,不同干扰强度对林分结构产生最直接的影响。随着干扰强度的加剧,林分结构退化,森林生态功能衰退,甚至导致物种濒危甚至灭绝,从而间接影响到物种组成。
人为干扰对常绿阔叶次生林的物种多样性产生负面影响。随着干扰等级的增大,林分物种数减少,各多样性指数和均匀度指数均呈现递减的趋势。这与认为适度干扰能够增加物种多样性的“中度干扰假说”的结论不大一致[42],但与很多其他森林群落乔木物种多样性研究结果一致。这可能与当地民众滥砍盗伐具有较高经济价值的壳斗科Fagaceae优势树种有关,尤其以栲树为主。
重度干扰林分的Simpson优势度指数高于轻度干扰林分。在所有干扰等级林分中,栲树均为优势树种,在重度干扰林分的重要值最高。这与栲树自身耐荫性有关,有利于早期幼苗生长。随着干扰等级的增强,留下的伴生树种多为耐荫树种,能够忍受优势树种与它们进行光照、水分和养料的竞争,同时也与现阶段林场采用封山育林的阔叶林经营措施相关。因此,不同人为干扰强度下呈现不同的物种多样性特征与人为干扰作用范围和物种耐受性、群落繁殖策略等生态习性有关。
由于人为干扰的复杂性导致干扰强度的量化问题研究一直十分困难。本研究加入林分干扰因素,综合建立适当的干扰体系,定量干扰强度,与当地的实际情况较为符合,能够客观反映出常绿阔叶次生林的林分结构及多样性在不同干扰程度下呈现出的规律性。
Forest structure and tree species diversity across a disturbance gradient in evergreen broadleaved secondary forests
-
摘要: 研究人为干扰对福建将乐林场常绿阔叶次生林林分结构及树种多样性的影响,为常绿阔叶次生林的植被恢复和生物多样性保护提供科学依据。以将乐林场常绿阔叶次生林为研究对象,通过踏查选择试验区立地条件相似且具有代表性的地段,设置25块(0.04~0.12 hm2)固定样地,采用相对影响法将常绿阔叶次生林分为5个干扰等级,采用方差分析和LSD多重比较法分析不同干扰等级林分结构及树种多样性变化。结果表明:不同干扰等级下常绿阔叶次生林直径结构均呈倒"J"型分布,干扰等级Ⅴ各径阶个体数显著小于其他干扰等级(P=0.022),林分潜在更新能力和生产力较低;干扰等级Ⅰ~Ⅳ树高呈单峰山状曲线,干扰等级Ⅴ为多峰山状曲线,林分垂直结构不稳定。随着干扰强度的增加,角尺度(P=0.045)逐渐增大,大小比数(P=0.039)和混交度(P=0.017)逐渐减小。α多样性指数随着干扰等级的升高呈递减的趋势(P < 0.05),Simpson优势度指数随着干扰等级的升高而呈上升趋势(P=0.000)。不同干扰等级林分之间物种相似度较低(Jaccard's相似指数 < 47%)。不同干扰等级常绿阔叶次生林林分结构差异显著,随着干扰等级的增加,乔木多样性及林分结构稳定性降低,不利于植被恢复及多样性保护。Abstract: To provide scientific evidence for vegetative recovery and biodiversity protection of evergreen broadleaved secondary forests, changes in stand structure and tree species diversity across a disturbance gradient of evergreen broadleaved secondary forests in Jiangle Forest Farm, Fujian Province were studied. The representative locations with similar site condition were selected through the field surveys, then a total of 25(0.04-0.12 hm2) permanent sample plots were set using typical sampling methods for evergreen broadleaf secondary forests. Five disturbance gradients (Ⅰ-Ⅴ with Ⅴ having the most disturbance) were assigned based on the relative value of major disturbance indicators. Then using ANOVA and LSD multiple comparision to analyze changes in stand structure and species diversity along a disturbance gradient. Results showed that for the five disturbance gradients, individual tree diameter conformed to an inverse J-shape curve. The individual number of Ⅴ disturbance gradient was significantly lower than other disturbance gradients (P=0.022) indicating the lowest capacity of natural regeneration and productivity. The individual tree heights of Ⅰ-Ⅳ disturbance gradients conformed to a normal curve; whereas, the Ⅴ disturbance gradient conformed to a bimodal curve indicating the lower stability in the forests vertical structure. With an increasing disturbance gradient, uniform angle indexes(P=0.045) increased, but neighborhood comparison indexes(P=0.039) and mingling indexes(P=0.017) decreased. The alpha diversity measures declined(P < 0.05) along the disturbance gradient as dominance increased(P=0.000). In addition, the similarity in species composition between each pair of disturbed forests was generally low (Jaccard's similarity index < 47%). With an increase in disturbance gradient, average DBH, average tree height, and alpha diversity measures decreased(P < 0.05), but the dominance index increased(P=0.000). Overall, as the disturbance gradient increased in the evergreen broadleaf secondary forests, the diversity index and stability of the forest structure decreased, which was not conducive to vegetative recovery and species diversity.
-
Key words:
- forest ecology /
- evergreen broadleaved secondary forest /
- important value /
- species richness /
- diversity
-
图 1 不同干扰等级下胸径和树高分布
Figure 1 Diztribution of DBH and tree keigkt along a dizturbanse gradient
表 1 样地基本情况表
Table 1. Bazis zituation of ztand
样地号 坡度/(°) E N 面积/hm2 郁闭郁闭 平均胸径/cm 平均树高/m 树种组成 1 40° 129°02, 33.72" 46°41, 50.22" 0.06 0.7 19.0 9.7 5米槠3栲树2苦槠+檫木+木荷 2 44° 129°02'33.72" 46°41'50.22" 0.06 0.7 18.0 9.3 5苦槠1栲树1米槠1油桐1山矾1赤杨叶+茅栗+檫木+福建 3 40° 117°26'06.72" 26°42' 15.18" 0.06 0.7 13.9 8.1 5赤杨叶2枫香1檵木1栲树1苦槠+黄樟+天仙果 4 25° 117°26'06.78" 26°43'40.80" 0.06 0.8 12.5 11.3 4栲树2赤杨叶1枫香1苦槠1木荷1南酸枣+黄樟+天仙果 5 23° 117°24'43.44" 26°47'36.30" 0.06 0.8 13.5 9.8 3栲树2苦槠1赤杨叶1青冈1杉木1木荷1南酸枣 6 23° 117°24'44.10" 26°47'37.32" 0.06 0.8 10.8 8.8 4赤杨叶3栲树1檫木1木荷1杉木+苦槠+光叶山矾+青冈+黄樟+冬青 7 38° 117°24'44.76" 26°47'36.24" 0.06 0.6 15.3 7.0 6栲树1木荷1苦槠1杨梅1青冈+南酸枣+台湾冬青+檵木+刨花润楠 8 37° 117°21'57.30" 26°47'5.40" 0.06 0.5 19.4 9.5 7栲树2青冈1甜槠+细柄阿丁枫+枫香+苦槠 9 34° 117°21'57.30" 26°47'5.40" 0.06 0.6 16.4 10.2 5栲树2青冈1南酸枣1山矾1木荷+米槠+苦槠 10 36° 117°22' 13.26" 26°46'54.42" 0.06 0.6 13.7 9.9 4栲树2木荷1黄樟1甜槠1青冈1黄毛润楠+米槠+山矾+南酸枣 11 35° 117°24'51.30" 26°47'51.90" 0.06 0.8 12.5 9.8 3栲树2赤杨叶2臭椿1木荷1杨桐1苦槠+黄樟+冬青+南酸枣 12 31° 117°24'07.26" 26°47'51.54" 0.06 0.6 15.8 8.4 8赤杨叶1黄樟1檵木+椤木石楠+天仙果+猴欢喜+枫香 13 31° 117°26'08.46" 26°43'39.30" 0.06 0.5 14.9 7.9 5苦槠1栲树1米槠1油桐1山矾1赤杨叶+茅栗+檫木+福建青冈 14 25° 117°24'41.46" 26°47'35.16" 0.12 0.7 12.2 9.2 4栲树2赤杨叶1杉木1南酸枣1木荷1苦槠+马尾松+木油桐+光叶山矾+杨梅 15 40° 117°22'20.22" 26°46'53.70" 0.06 0.6 6.9 6.1 4赤杨叶2刨花润楠1山乌桕1樟树1绒毛山胡椒1栲树 16 39° 117°22' 14.88" 26°46'47.98" 0.06 0.6 9.3 8.3 3栲树2柿1杉木1滇青冈1赤杨叶1浙闽樱 17 38° 117°22'14.40" 26°46'49.32" 0.06 0.6 7.9 6.8 5苦槠2栲树1山乌桕1红楠 18 37° 117°22'16.35" 26°46'50.51" 0.06 0.6 13.5 8.8 3苦槠2栲树2马尾松1山乌桕1枫香1红楠 19 24° 117°25'23.29" 26°47'36.01" 0.06 0.9 11.9 11.6 2栲树2苦槠1木荷1刨花润楠1吊皮锥1乐昌含笑1红楠 20 24° 117°25'23.40" 26°47'36.78" 0.04 0.9 11.5 12.4 4栲树1木蜡树1细柄阿丁枫1吊皮锥1红楠1木荷 21 36° 117°24'52.74" 26°47'56.10" 0.06 0.8 11.6 9.7 5栲树2黄樟1刺毛杜鹃1苦槠+鼠刺 22 41° 117°24'52.02" 26°47'50.92" 0.06 0.8 10.3 9.7 5栲树2木荷1苦槠1罗浮锥 23 31° 117°19'19.00" 26°31'28.00" 0.06 0.6 17.4 10.4 5甜槠2红楠1细齿叶柃1福建冬青 24 32° 117°18'21.00" 26°31'22.00" 0.06 0.8 13.7 10.6 4栲树3甜槠2石木姜子 25 30° 117°15'33.00" 26°32'36.00" 0.04 0.7 14.5 11.0 4甜槠3栲树1木荷1苦槠+米槠+刨花润楠 说明:油桐Vernicia fordii; 茅栗Castanea seguinii; 福建青冈Cyclobalanopsis chungii; 天仙果Ficuss beecheyana; 台湾冬青Iles formosana; 刨花润楠Machilus pauhoi; 细柄阿丁枫Altingia angustifolia; 黄毛润楠Machilus chrysotricha; 臭椿Ailanthus altissima; 杨桐Adinandra millettii; 椤木石楠Photinia davidsoniae; 猴欢喜Sloanea sinensis; 马尾松Pinus massoniana; 木油桐Vernicia montana; 山乌桕Sapium discolor; 樟树Cinnamomum camphora; 绒毛山胡椒Lindera nacusua; 柿Diospyros kaki; 滇青冈Cyclobalanopsis glaucoides; 浙闽樱(桃)Cerasus schneideriana; 红楠Machilus thunbergii; 吊皮锥Castanopsis karakamii; 乐昌含笑Michelia chapensis; 木蜡树Tosicodendron sylvestre; 刺毛杜鹃Rhododendron championae; 鼠刺Itea chinensis; 罗浮锥Castanopsis fabri; 细齿叶柃Eurya nitida; 福建冬青Iles fukienensis; 石木姜子Litsea elongata  下载: 导出CSV
下载: 导出CSV
表 2 常绿阔叶次生林干扰等级
Table 2. Dizturbanse gradient of evergreen broad-leaved zesondary foreztz
干扰等级 样地号 相对郁度 幼树相对密度 受害木相对密度 受害木相对断面积 伐倒木相对密度 伐倒木相对断面积 总和 干扰等级 样地号 相对郁度 幼树相对密度 受害木相对密度 受害木相对断面积 伐倒木相对密度 伐倒木相对断面积 总和 Ⅰ 1 1.2 6.2 1.0 1.1 3.9 12.2 17.8 Ⅲ 8 1.2 8.5 18.0 10.7 2.9 1.9 52.7 Ⅰ 2 1.2 6.1 1.3 2.1 2.9 1.5 15.1 Ⅲ 9 1.7 3.0 21.7 23.4 2.9 3.9 55.5 Ⅰ 3 1.6 5.1 1.3 1.5 1.0 2.9 25.6 Ⅲ 11 1.6 4.2 18.3 15.6 5.9 13.0 64.0 Ⅰ 4 1.2 8.3 2.0 3.6 3.9 6.9 18.7 Ⅲ 12 1.7 1.7 17.3 21.8 2.9 13.3 61.6 Ⅰ 5 1.4 1.0 6.3 15.3 2.9 1.7 25.9 Ⅲ 15 1.6 9.3 24.3 20.6 4.4 9.9 56.6 Ⅰ 6 1.6 3.4 5.3 3.6 2.9 1.0 13.4 Ⅲ 19 1.2 3.1 1.7 1.0 2.9 8.3 58.7 Ⅰ 7 1.4 1.7 12.7 34.8 1.0 1.1 28.6 Ⅲ 20 1.6 4.9 10.0 24.4 1.9 4.7 58.6 Ⅰ 18 1.2 3.7 5.7 3.0 3.9 1.2 18.2 Ⅳ 16 1.2 4.9 27.3 25.9 2.9 6.2 70.1 Ⅱ 10 1.0 4.2 4.3 4.4 3.9 19.5 37.3 Ⅳ 22 1.6 4.4 4.7 18.9 1.9 8.0 68.4 Ⅱ 13 1.4 3.0 22.0 22.7 2.9 3.5 32.4 Ⅳ 23 1.2 6.7 13.3 20.1 15.7 16.0 73.0 Ⅱ 14 1.2 6.2 16.7 25.9 1.9 9.7 43.2 Ⅴ 24 1.2 5.9 23.0 29.4 8.7 25.8 94.0 Ⅱ 17 1.0 8.7 11.0 8.0 1.9 1.8 47.5 Ⅴ 25 1.4 10.1 43.3 37.4 1.9 7.0 101.1 Ⅱ 21 1.6 4.1 21.7 31.4 1.9 3.3 39.5
下载: 导出CSV
表 3 干扰度指数分级
Table 3. Hierarchical list of index of disturbance
干扰等级 干扰度值 Ⅰ 13.4≤FDI < 30.9 Ⅱ 30.9≤FDI < 48.5 Ⅲ 48.5≤FDI < 66.0 Ⅳ 66.0≤FDI < 83.6 Ⅴ 83.6≤FDI < 101.1
下载: 导出CSV
表 4 不同干扰等级林分基本情况
Table 4. Basic situation of forest along a disturbance gradient
干扰等级 林分密度/(株·hm-2) 平均胸径/cm 平均树高/m 幼树密度/(株·hm-2) Ⅰ 1 215 14.3 11.7 381 Ⅱ 1 208 14.4 10.5 367 Ⅲ 1 448 13.2 9.8 335 Ⅳ 1 627 12.1 9.1 605 Ⅴ 600 11.8 8.8 100
下载: 导出CSV
表 5 不同干扰等级空间结构特征
Table 5. Structural characteristics of spatial structure along a disturbance gradient
干扰等级 角尺度 大小比数 混交度 Ⅰ 0.559 7 0.506 5 0.761 6 Ⅱ 0.564 3 0.500 0 0.737 2 Ⅲ 0.566 4 0.499 0 0.710 7 Ⅳ 0.582 7 0.492 2 0.708 3 Ⅴ 0.590 8 0.485 0 0.676 1 P 0.045 < 0.05 0.039 < 0.05 0.017 < 0.05
下载: 导出CSV
表 6 不同干扰等级下α多样性
Table 6. α-diversity along a different disturbance gradient
干扰等级 多样性测量 物种丰富度 特有物种数目 Margalef指数 Shannon-Wiener指数 个体物种数 Pielou指数 Simpson优势度指数 Ⅰ 44 6 4.50 ± 0.49 2.60 ± 0.12 0.24 ± 0.04 1.20 ± 0.08 0.13 ± 0.02 Ⅱ 59 17 3.51 ± 0.19 2.25 ± 0.07 0.20 ± 0.01 0.81 ± 0.02 0.15 ± 0.01 Ⅲ 45 8 3.51 ± 2.22 2.22 ± 0.13 0.22 ± 0.04 0.85 ± 0.02 0.10 ± 0.02 Ⅳ 41 9 3.62 ± 0.75 2.31 ± 0.23 0.19 ± 0.03 0.82 ± 0.03 0.11 ± 0.02 Ⅴ 31 6 3.20 ± 0.39 1.49 ± 1.20 0.12 ± 0.05 0.45 ± 0.33 0.52 ± 0.40 F 3.359 13.316 1.261 24.402 82.596 P 0.029<0.05 0.000<0.05 0.032<0.05 0.000<0.05 0.000<0.05
下载: 导出CSV
表 7 不同干扰等级下物种相似值
Table 7. Jaccard's coefficient along a disturbance gradient
干扰等级 Ⅰ Ⅱ Ⅲ Ⅳ Ⅴ Ⅰ 100 45.8 38.5 40.0 27.1 Ⅱ 100 46.6 32.1 28.8 Ⅲ 100 35.4 33.9 Ⅳ 100 34.1 Ⅴ 100
下载: 导出CSV
-
[1] SAGAR R, RAGHUBANSHI A S, SINGH J S. Tree species composition, dispersion and diversity along a disturbance gradient in a dry tropical forest region of India[J]. For Ecol Manage, 2003, 186(1/3):61-71. [2] ZHU Jiaojun, MAO Zhihong, HU Lile, et al. Plant diversity of secondary forests in response to anthropogenic disturbance levels in montane regions of northeastern China[J]. J For Res, 2007, 12(6):403-416. [3] LAMB D, ERSKINE P D, PARROTTA J A. Restoration of degraded tropical forest landscapes[J]. Science, 2005, 310(5754):1628-1632. [4] 苏日古嘎, 张金屯, 王永霞.北京松山自然保护区森林群落物种多样性及其神经网络预测[J].生态学报, 2013, 33(11):3394-3403. Suriguga, ZHANG Jintun, WANG Yongxia. Species diversity of forest communities and its forecasting by neural network in the Songshan National Nature Reserve, Beijing[J]. Acta Ecol Sin, 2013, 33(11):3394-3403. [5] 鲁庆彬, 游卫云, 赵昌杰, 等.旅游干扰对青山湖风景区植物多样性的影响[J].应用生态学报, 2011, 22(2):295-302. LU Qingbin, YOU Weiyun, ZHAO Changjie, et al. Effects of tourism disturbance on plant diversity in Qingshan Lake scenic area of Zhejiang Province[J]. J Appl Ecol, 2011, 22(2):295-302. [6] 陈芙蓉, 程积民, 刘伟, 等.不同干扰对黄土区典型草原物种多样性和生物量的影响[J].生态学报, 2013, 33(9):2856-2866. CHEN Furong, CHENG Jimin, LIU Wei, et al. Effects of different disturbances on diversity and biomass of communities in the typical steppe of loess region[J]. Acta Ecol Sin, 2013, 33(9):2856-2866. [7] 王海梅, 李政海, 王珍.气候和放牧对锡林郭勒地区植被覆盖变化的影响[J].应用生态学报, 2013, 24(1):156-160. WANG Haimei, LI Zhenghai, WANG Zhen. Effects of climate and grazing on the vegetation cover change in Xilinguole League of Inner Mongolia, North China[J]. Chin J Appl Ecol, 2013, 24(1):156-160. [8] 周伶, 上官铁梁, 郭东罡, 等.晋、陕、宁、蒙柠条锦鸡儿群落物种多样性对放牧干扰和气象因子的响应[J].生态学报, 2012, 32(1):111-122. ZHOU Ling, SHANGGUAN Tieliang, GUO Donggang, et al. Response of species diversity in Caragana korshinskii communities to climate factors and grazing disturbance in Shanxi, Shaanxi, Ningxia and Inner Mongolia[J]. Acta Ecol Sin, 2012, 32(1):111-122. [9] SAEKI I. Effects of tree cutting and mowing on plant species composition and diversity of the wetland ecosystems dominated by the endangered maple, Acer pycnanthum[J]. For Ecol Manage, 2007, 242(2/3):733-746. [10] ANITHA K, JOSEPH S, CHANDRAN R J, et al. Tree species diversity and community composition in a human-dominated tropical forest of Western Ghats biodiversity hotspot, India[J]. Ecol Complex, 2010, 7(2):217-224. [11] MUTHULINGAM U, THANGAVEL S.Density, diversity and richness of woody plants in urban green spaces:a case study in Chennai Metropolitan City[J]. Urban For Urban Green, 2012, 11(4):450-459. [12] 吴甘霖, 黄敏毅, 段仁燕, 等.不同强度旅游干扰对黄山松群落物种多样性的影响[J].生态学报, 2006, 26(12):3924-3930. WU Ganlin, HUANG Minyi, DUAN Renyan, et al. Disturbing effects of tourism on species diversity in Pinus taiwanensis communities[J]. Acta Ecol Sin, 2006, 26(12):3924-3930. [13] 邱扬, 李湛东, 张玉钧, 等.火干扰对大兴安岭北部原始林下层植物多样性的影响[J].生态学报, 2006, 26(9):2863-2869. QIU Yang, LI Zhandong, ZHANG Yujun, et al. The effects of fire disturbance on the biodiversity of understory plant in virginal forest, northern region of Daxinganling Mountains, China[J]. Acta Ecol Sin, 2006, 26(9):2863-2869. [14] FULBRIGHT T E. Disturbance effects on species richness of herbaceous plants in a semi-arid habitat[J]. J Arid Environ, 2004, 58(1):119-133. [15] PHILLIPS O L, HALL P, GENTRY A H, et al. Dynamics and species richness of tropical rain forests[J]. Proc Natl Acad Sci USA, 1994, 91(7):2805-2809. [16] SCHWILK D W, KEELEY J E, BOND W J. The intermediate disturbance hypothesis does not explain fire and diversity pattern in fynbos[J]. Plant Ecol, 1997, 132(1):77-84. [17] WILSON S D, TILMAN D. Interactive effects of fertilization and disturbance on community structure and resource availability in an old-field plant community[J]. Oecologia, 1991, 88(1):61-71. [18] PILLIOS O, HALL P, SAWYER S, et al. Species richness, tropical forest dynamics and sampling:response to Sheil[J]. Oikos, 1997, 79(1):183-187. [19] 王建国.建德山区的常绿阔叶林生态效益分析[J].植物生态学与地植物学丛刊, 1985, 9(2):112-118. WANG Jianguo. Analysis of the ecological benefits of the evergreen broad-leaved forests in the mountain area of Jiande County[J]. Acta Phytoecol Geobot Sin, 1985, 9(2):112-118. [20] 刘伦辉, 刘文耀, 郑征.滇中山地主要植物群落水土保持效益比较[J].水土保持学报, 1990, 4(1):36-43. LIU Lunhui, LIU Wenyao, ZHENG Zheng. Comparison of the effects on the soil and water conservation of main communities in Tonghai County of Yunnan[J]. Acta Conserv Soil Aquae Sin, 1990, 4(1):36-43. [21] 陈平腾.福建南平市延平区常绿阔叶次生林林分结构和物种多样性研究[D].福州: 福建农林大学, 2009. CHEN Pingteng. Studies on Stand Structure and Species Diversity of Evergreen Broad-leaved Forest in Yanping District, Nanping, Fujian[D]. Fuzhou: Fujian Agriculture and Forestry University, 2009. [22] 杨青, 丁晖, 方炎明, 等.武夷山大安源甜槠常绿阔叶林群落物种组成及多样性分析[J].植物资源与环境学报, 2014, 23(1):44-50. YANG Qing, DING Hui, FANG Yanming, et al. Analyses on species composition and diversity of evergreen broad-leaved forest community of Castanopsis eyrei at Da'anyuan in Wuyi Mountain[J]. J Plant Resour Environ, 2014, 23(1):44-50. [23] 贺金生, 陈伟烈, 谢宗强, 等.福建龙栖山自然保护区的常绿阔叶林类型及其群落物种多样性分析[J].生态学杂志, 1998, 17(3):1-6. HE Jinsheng, CHEN Weilie, XIE Zongqiang, et al. Types of the evergreen broadleaf forest and their community species diversity in Longqishan Nature Reserve, Fujian Province[J]. Chin J Ecol, 1998, 17(3):1-6. [24] 丁晖, 杨云方, 徐海根, 等.武夷山典型常绿阔叶林物种组成与群落结构[J].生态学报, 2015, 35(4):1142-1154. DING Hui, YANG Yunfang, XU Haigen, et al. Species composition and community structure of the typical evergreen broad-leaved forest in the Wuyi Mountains of Southeastern China[J]. Acta Ecol Sin, 2015, 35(4):1142-1154. [25] 包维楷, 刘照光, 刘朝禄, 等.亚热带次生常绿阔叶林主要乔木种群自然恢复15年来的变化[J].林业科学, 2001, 37(1):7-14. BAO Weikai, LIU Zhaoguang, LIU Zhaolu, et al. Fifteen-year change of tree populations in the secondary subtropical evergreen broad-leaved forests in southwestern China[J]. Sci Silv Sin, 2001, 37(1):7-14. [26] 杜燕, 郑小贤, 罗梅.将乐县常绿阔叶次生林主导功能类型的研究[J].中南林业科技大学学报, 2013, 33(2):39-43. DU Yan, ZHENG Xiaoxian, LUO Mei. Study on leading function types of evergreen broad-leaved forest in Jiangle County, Fujian Province[J]. J Cent South Univ For Technol, 2013, 33(2):39-43. [27] 任引.武夷山主要类型常绿阔叶次生林结构与功能特征的研究[D].南京: 南京林业大学, 2008. REN Yin. Study on Structure and Function Characteristics of Evergreen Broad-leaved Forest in Wuyi Mountain[D]. Nanjing: Nanjing Forestry University, 2008. [28] 宋育红, 张新文, 周斌.格氏栲自然保护区常绿阔叶林群落特征[J].生态科学, 2005, 24(3):228-232. SONG Yuhong, ZHANG Xinwen, ZHOU Bin. Features of evergreen broad leaved forest community in Castanopsis kawakamii Nature Reserve[J]. Ecol Sci, 2005, 24(3):228-232. [29] 黄建雄, 郑凤英, 米湘成.不同尺度上环境因子对常绿阔叶林群落的谱系结构的影响[J].植物生态学报, 2010, 34(3):309-315. HUANG Jianxiong, ZHENG Fengying, MI Xiangcheng. Influence of environmental factors on phylogenetic structure at multiple spatial scales in an evergreen borad-leaved forest of China[J]. Chin J Plant Ecol, 2010, 34(3):309-315. [30] 罗梅, 郑小贤.福建将乐林场常绿阔叶次生林主要种群种间联结性研究[J].西北农林科技大学学报(自然科学报), 2016, 44(8):135-142. LUO Mei, ZHENG Xiaoxian. Iner-specific correlation of main populations in evergreen broad-leaved secondary forest in Jiangle forest farm[J]. J Northwest A&F Univ Nat Sci Ed, 2016, 44(8):135-142. [31] 李杰, 高祥, 徐光, 等.福建将乐林场常绿阔叶林幼苗结构分析与更新评价[J].西北农林科技大学学报(自然科学版), 2014, 42(5):62-68. LI Jie, GAO Xiang, XU Guang, et al. Structure analysis and evaluation of seedlings in evergreen broad-leaved forest in Jiangle Forest Farm, Fujian[J]. J Northwest A&F Univ Nat Sci Ed, 2014, 42(5):62-68. [32] 杜燕.将乐林场常绿阔叶次生林健康评价研究[D].北京: 北京林业大学, 2013. DU Yan. Health Assessment of Evergreen Broad-leaved Forest in Jiangle Forest Farm[D]. Beijing: Beijing Forestry University, 2013. [33] 王琦, 杜燕, 郑小贤.将乐林场常绿阔叶林健康评价指标体系构建[J].西南林业大学学报, 2014, 34(5):74-78. WANG Qi, DU Yan, ZHENG Xiaoxian. Construct the health evaluation of index system for evergreen broadleaf forest in Jiangle Forest Farm[J]. J Southwest For Univ, 2014, 34(5):74-78. [34] 周洋.福建三明将乐林场低质低效常绿阔叶次生林经营技术[D].北京: 北京林业大学, 2015. ZHOU Yang. Study on Low-quality and Low-benefit of Broad-leaved Evergreen Forests Management Techniques in Jiangle, Fujian[D]. Beijing: Beijing Forestry University, 2015. [35] 陈昌雄.天然常绿阔叶次生林生长潜力及择伐经营研究[D].南京: 南京林业大学, 2005. CHEN Changxiong. Study on Growth Potential and Selection Cutting of Natural Evergreen Broad-leaved Forests[D]. Nanjing: Nanjing Forestry University, 2015. [36] WU Lichao, LIU Jie, ATSUSHI T, et al. Effect of selective logging on stand structure and tree species diversity in a subtropical evergreen broad-leaved forest[J]. Ann For Sci, 2013, 70:535-543. [37] CHRONOPOULOS G, CHRISTODOULAKIS D. Analysis of the adventive flora of a Greek city-the example of Patras[J]. Bot Helv, 2000, 110(2):171-189. [38] 汤孟平, 唐守正, 雷相东.两种混交度的比较分析[J].林业资源管理, 2004, 4(8):25-27. TANG Mengping, TANG Shouzheng, LEI Xiangdong. Comparison analysis on two minglings[J]. For Resour Manage, 2004, 4(8):25-27. [39] 罗梅, 郑小贤.八达岭辽东栎-油松混交林空间结构及其多样性[J].中南林业科技大学学报, 2012, 32(9):55-58. LUO Mei, ZHENG Xiaoxian. Study of spatial structure and species diversity of Quercus liaotungensis-Piuns tabulaeformis mixed stand in Badaling forest farm[J]. J Cent South Univ For Technol, 2012, 32(9):55-58. [40] BAKER W L. The landscape ecology of large disturbances in the design and management of nature reserves[J]. Landscape Ecol, 1992, 7(3):181-194. [41] MAROZAS V, RACINSKAS J, BARTKECICIUS E. Dynamics of ground vegetation after surface fires in hemiboreal Pinus sylvestris forests[J]. For Ecol Manage, 2007, 250(1):47-55. [42] COLLINS S L, GLENN S M, GIBSON D J. Experimental analysis of intermediate disturbance and initial floristic composition:decoupling cause and effect[J]. Ecology, 1995, 76(2):486-492. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2019.01.004
 点击查看大图
点击查看大图
计量
- 文章访问数: 4768
- HTML全文浏览量: 965
- PDF下载量: 179
- 被引次数: 0
