





-
氮素的吸收、同化与运转直接影响着作物的生长发育状况,对作物生长至关重要。传统计算植株氮质量分数的方法需要破坏性取样,且要进行室内化学分析,具有一定程度上的滞后性和缓慢性。叶绿素仪具有快速、简便和无损的特点,常被用来监测诊断作物氮素营养[1]。前人基于作物高产条件下建立的土壤、作物分析仪器开发(SPAD)值与产量的关系确定了适宜SPAD值[2-4]。近年来,在小麦Triticum aestivum,玉米Zea mays,茅草Imperata cylindrica,水稻Oryza sativa等作物上利用SPAD值估算作物氮质量分数快速无损诊断氮素营养状况已经被广泛应用[3],但传统的叶片氮营养诊断方法有不少的缺点,例如准确度低,诊断费时费力,且破坏样本和诊断相对滞后[5-7]。若采用SPAD测量仪诊断氮素营养方法则较为省时省力且能够实时监测,但SPAD测量仪所测量出的数据受很多环境因素影响,在某些情况下会出现准确度低和稳定性较差等缺点。前人主要以单叶SPAD作为诊断方法进行研究[8],凌启鸿等[9]研究表明顶3叶与顶4叶的叶色差可以准确诊断水稻植株氮素的丰亏,李刚华等[8]研究表明顶3叶可作为较为理想指示叶诊断水稻氮素营养状况,姜继萍等[10]研究表明水稻顶4叶和顶3叶间的差值与施氮水平之间存在明显的相关性,李杰等[11]研究表明顶3叶SPAD值乘顶4叶SPAD值/顶部4张叶片平均SPAD值与田块表观供氮量之间具有显著的线性关系。前人大都基于顶3叶与顶4叶对水稻氮素营养诊断进行研究,而不同叶位构建归一化SPAD指数(normalized differential SPADij,INDSPADij)与氮素营养之间关系的研究较少。本研究通过不同叶位建立归一化SPAD指数与水稻氮素营养之间的定量关系,通过归一化SPAD值估算氮质量分数,从而快速精确地诊断水稻氮素营养,并通过不同叶位与氮素营养状况的相关性高低判定更能指示水稻植株氮质量分数的叶位,以期为水稻氮素营养诊断提供参考。
-
于2015年在浙江省德清县浙江农林大学现代农业科技园,选用2个水稻品种:‘甬优538’‘Yongyou 358’,5月28日播种,大田用种量为18.0 kg·hm-2,秧龄为25 d,2本·丛-1,移栽稻密度为18.0万穴·hm-2;行距28.0 cm ×株距20.0 cm。‘秀水134’‘Xiushui 134’,5月28日播种,大田用种量为45.0 kg·hm-2,秧龄25 d,4本·丛-1,移栽稻密度为25.5万穴·hm-2。行距28.0 cm ×株距14.0 cm。2品种独立试验,设置5个氮肥水平,重复3次,采用随机区组设计,小区15个·品种-1。小区之间以田埂相隔,埂上覆膜,独立排灌。南北行向,小区东西长8.0 m,南北宽4.0 m,面积为32.0 m2。耕作层土壤有机质为20.10 g·kg-1,全氮为1.19 g·kg-1,碱解氮为104.64 mg·kg-1,速效磷为46.45 mg·kg-1,速效钾为81.49 mg·kg-1。
-
设置5个氮肥水平,施肥量分别为0(N0对照),70.0(N1),140.0(N2),210.0(N3),280.0(N4)kg·hm-2。m(基肥):m(糵肥):m(穗肥)=4:2:4(表 1)。在施用基肥时,一次性施入过磷酸钙990.0 kg·hm-2;氯化钾基肥用175.0 kg·hm-2,穗肥用175.0 kg·hm-2,所有小区水平一致。
表 1 不同时期氮肥用量
Table 1. Nitrogen consumption at different stages
编号 氮肥/(kg·hm-2) 组成/(kg·hm-2) 基肥 蘖肥 穗肥 N0 0 0 0 0 N1 70.0 28.0 14.0 28.0 N2 140.0 56.0 28.0 56.0 N3 210.0 84.0 42.0 84.0 N4 280.0 112.0 56.0 112.0 -
通过分析顶部4张叶片SPAD值的变化特征,明确叶位SPAD归一化指数与植株氮质量分数的定量关系,构建最优的叶位SPAD组合指数,实时可靠地反映水稻植株氮素营养状况。
叶位SPAD组合指数的计算方法如下:叶位SPAD归一化指数INDSPADij=(ISPADi-ISPADj)/(ISPADi+ISPADj)。其中:ISPADi和ISPADj分别代表水稻冠层主茎第i和j叶位的SPAD值,i,j≤4。
-
用MINOLTA产SPAD-502型叶绿素计,在水稻拔节前期、拔节期、孕穗期、抽穗期、开花期和灌浆前期,选取4株健康主茎测定其顶部4张完全展开叶上部1/3处、中部和下部1/3处的SPAD值,取平均值作为各叶片的SPAD值[8]。
-
水稻拔节前期、拔节期、孕穗期、抽穗期、开花期、灌浆前期、灌浆中期和成熟期,分别选取代表性水稻植株4株·小区-1,根据植株器官发育情况,将样品植株分离为叶、茎和穗,放入恒温干燥箱内烘干,在105 ℃杀青30 min,80 ℃烘干48 h至恒量后称量。样品粉碎后使用半微量凯氏定氮法测定水稻植株不同器官的全氮质量分数。
-
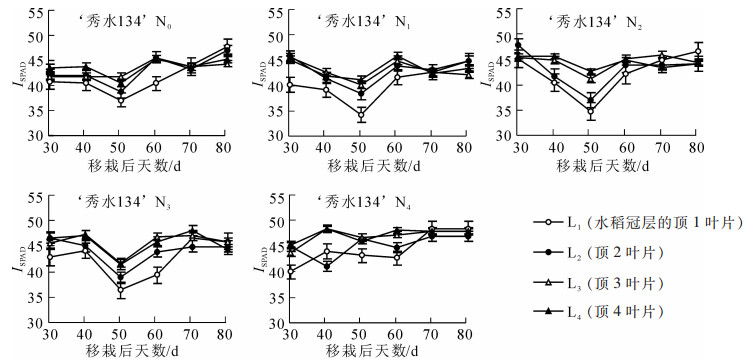
图 1和图 2显示:L1~L4分别代表水稻冠层的顶1,顶2,顶3及顶4叶片,N0~N4分别代表 5个施氮水平0,70.0,140.0,210.0,280.0 kg·hm-2。从图 1可以看出:‘甬优538’在不同时间SPAD值出现明显差异,进入衰老期后期SPAD值进入持续下降阶段,大部分植株在衰老期前后L4的SPAD值下降数值明显大于L1,L2和L3。‘秀水134’SPAD值呈现先降低后增加再降低趋势,进入衰老期后,SPAD值下降明显。

图 1 ‘甬优538’水稻冠层叶片SPAD值在不同的5个施氮水平下的动态变化趋势
Figure 1. Trend of dynamic change of SPAD value of canopy leaf of 'Yongyou 538' rice under different 5 nitrogen fertilizer levels

图 2 ‘秀水134’水稻品种冠层叶片SPAD值在不同的5个施氮水平下的动态变化趋势
Figure 2. Trend of dynamic change of SPAD value of canopy leaf of 'Xiushui 134' rice under different 5 nitrogen fertilizer levels
从图 1~2可以看出:‘甬优538’和‘秀水134’绝大部分L4的SPAD数值大于L3和L2,其中L1的数值在生育期中大部分低于L2,L3和L4。因为氮素都是从下往上运输的,所以最接近根部的L4的SPAD值大于其他叶片。又因为L1测定SPAD值时展开程度不同,L1对光能的利用效果不佳从而导致叶片颜色偏浅,SPAD数值低于其他叶片数值。
2015年的大田试验(对照除外),在移栽后30 d至移栽后90 d的生育期内,植株冠层叶片的SPAD值可以从叶片叶色的变化中看出,经过各个生育期叶色比对可以发现叶色经历了“二黑二黄”2次交替波动的变化,施氮量的增加叶色的变化程度明显随之加大。大田水稻在移栽后的30 d,即水稻处于分集高峰期,水稻叶片经历了的第一次叶色“黑黄”交替变化,这一时期水稻SPAD值达到一个峰值,从大田材料可观察到冠层叶色较深,随后变淡。水稻冠层叶片的深浅交替变化有助于分蘖数的增加,促进秧苗在育苗阶段的生长,保证在生育后期拥有充足的穗数。在植株进入这一阶段时植株氮代谢活跃,冠层叶片生长旺盛,分蘖数有所增加。其中‘甬优538’N4和‘秀水134’N3,N4的分蘖高峰期往后偏移。分蘖数量和施氮量等影响分蘖的终止日期,在分蘖的中后期,水稻植株对土壤供氮水平要求较高以求达到分孽所需的含氮水平。因而随着施氮量的增加,分孽的终止日期将延迟,分蘖期因此有所延长。所以高氮水平下水稻的分蘖高峰期也随之向后偏移[12-13]。
由于品种条件的影响,‘秀水134’秧苗生长较‘甬优538’偏慢,水稻处于抽穗期时(即‘甬优538’在移栽后的75 d后,‘秀水134’在移栽后的85 d后),叶片开始大量积累营养物质,叶片在这一阶段叶绿素和氮质量分数有所升高,叶片颜色较深,为之后的抽穗期提供物质储备。在植株进入抽穗期,穗部将大量接收叶片的营养物质,叶色又变淡(SPAD值下降),即水稻植株冠层叶片发生了第2次黑黄交替。“二黑二黄”是水稻植株茎秆生长增粗、防倒伏、稻穗增重增大的时期。通常“二黑”(2次颜色较深的时期)促进光合作用的进行,从而有效促进了碳水化合物的累积。若在生育期中“二黑”未出现或不明显,表明植株生长发育减缓,会造成以下后果:叶片偏小,抽穗期提早,穗部发育不良穗粒不足,导致产量降低。图 1和图 2可以看出:不同的施氮水平的水稻均会表现2次叶片颜色交替的现象(即SPAD值2次升降),不同的施肥水平和不同的施肥时间只会影响冠层叶片颜色深浅交替变化的幅度和出现的时间,并不能使这一变化的规律消失。有研究[9]认为,无论在何种施氮水平下,水稻冠层叶在生育期均会产生“黑黄”交替变化趋势,因为这是其自身内在因素控制的,不同的施氮水平无法使这一现象消失。
图 1和图 2表明:2个品种在不同施氮条件下,水稻L1的SPAD值普遍小于L4和L3。分析原因,在移栽后35 d左右,叶位靠下的L4及L3已经比较成熟且已完全展开[10],因而SPAD值较大。而L1为新生叶片,叶位靠上,因其可能为完全成熟且未完全展开,并且从大田实地观察也可以得出:L1叶面积较小,颜色较其他叶片较浅,导致SPAD值在大部分情况下小于L3和L4。
‘甬优358’在N0条件下及‘秀水134’在N0和N1条件下,水稻的L2的SPAD值大于L3及L4。分析原因,水稻植株缺氮条件下,L2会优先接收氮素营养,顶部叶片光合作用效率高于底部叶片,所以L2的SPAD值明显偏大。随着施氮水平的提高,L2~L4的SPAD差值变小,在而在氮素供给充足的情况下,L3和L4的SPAD值明显变大,出现了L3和L4的SPAD值大于L2的现象,推测是因为施氮量的增加,充足的氮素不仅可以满足新叶的生长也可以保证靠近根部的下位叶的氮素营养。可见,充足的氮素供应可以使不同叶位的功能叶生长发育更加平衡。另外也可以推测:L3和L4相比L2对氮素营养状况的相关性更高,因而更适合作为植株氮营养诊断的理想指示叶。通过不同叶片SPAD值表现可以看出氮素优先供应上部叶片,前期营养生长氮素主要供应L2,后期生殖生长氮素主要供应L1,当氮素充足的时候下部叶片能得到足够的养分供应,而当植株出现氮素亏缺,老叶最先衰老,下部叶片表现最为明显,因此,L4对氮素反应最为敏感,其次是L3,L4可以作为水稻植株氮素营养变化的指示叶片。此外,L1与L4的SPAD值差值比较发现,两者之间的差值随氮水平的升高逐渐减小。根据这一现象,可以利用上位叶和下位叶差值大小来判断叶片氮质量分数的高低,为下一步推断水稻施氮是否充足提供依据。
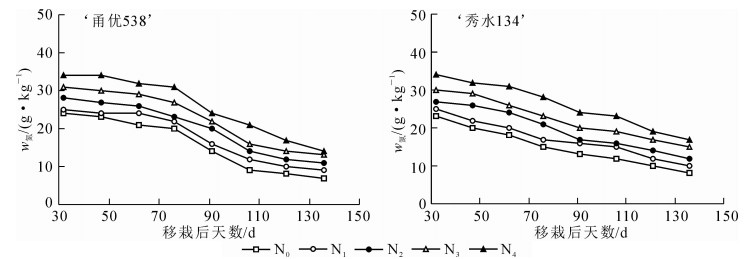
田间试验表明:水稻播种期、移栽期、拔节期、孕穗期、开花期和成熟期分别为5月28日、6月21日、8月10日、8月27日、9月7日和10月20日。图 3可以看出:随着施氮水平的增加水稻冠层叶片SPAD值逐渐增加,这说明施氮量越高,水稻叶片叶绿素质量分数也越高。

图 3 不同施氮量水平下‘甬优538’和‘秀水134’水稻冠层叶片SPAD值在的动态变化趋势
Figure 3. Trend of dynamic change of leaf SPAD values of 'Yongyou 538 'and 'Xiushui 134' under different 5 nitrogen fertilizer levels
-
由图 4可知:在各生育时期地上部冠层叶片氮质量分数随着施氮量的增加不断增加,5个施氮水平之间存在显著差异(P<0.05)。过量施氮会引起水稻干物质增加变缓,而冠层叶片氮浓度仍呈不断增加趋势。

图 4 不同施氮水平下水稻冠层叶片氮质量分数随移栽后天数的变化趋势
Figure 4. Variation trend of nitrogen concentration of rice leaf under different nitrogen levels with the days after transplantation
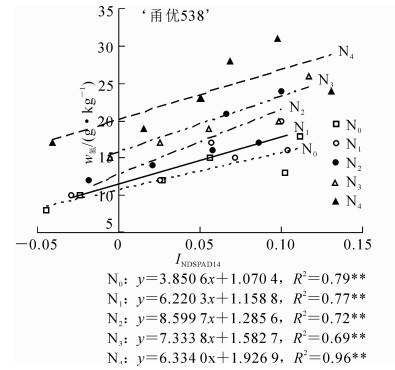
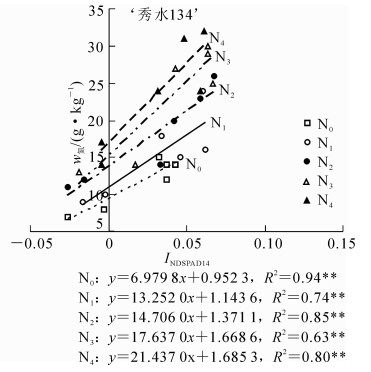
研究证实:水稻冠层不同叶位叶片SPAD差值与叶片氮质量分数为指数式回归关系,且此关系受年份、品种和生育时期影响较小[14]。本研究结果表明:以INDSPAD14与冠层叶片氮质量分数之间相关系数最为显著(图 5和图 6)。

图 5 ‘甬优358’水稻INDSPAD14与冠层叶片氮质量分数的定量关系
Figure 5. Quantitative relationship between the N content of leaf and INDSPAD14 in'Yongyou 358'

图 6 ‘秀水134’水稻INDSPAD14与冠层叶片氮质量分数的定量关系
Figure 6. Quantitative relationship between the leaf N content of rice and INDSPAD14 in'Xiushui 134'
为了减少品种和环境等因素对SPAD与植株氮质量分数关系的影响,对大田试验的‘甬优358’和‘秀水134’分别构建了归一化SPAD指数(INDSPADij)与植株氮质量分数之间关系,结果发现INDSPAD14与N0~N4所有不同施氮量组别之间呈显著正相关(P<0.05),‘甬优358’水稻品种决定系数R的范围为0.69~0.96,‘秀水134’决定系数R的范围为0.64~0.94,INDSPAD14与植株氮质量分数的定量关系受施氮量和环境的影响较小而保持稳定,可以用INDSPAD14来估算水稻植株氮质量分数。相比前人利用单叶SPAD和遥感等方法估算氮质量分数,本研究利用INDSPAD14估算水稻植株氮质量分数,简单快速,可以减少取样对植株带来的破坏以及植株品种、土壤、天气等对诊断的影响,仅测量水稻植株L1和L4 SPAD,即可估算出实际氮质量分数。另外,INDSPAD14指数对水稻植株氮素营养的估算效果优于INDSPAD13指数,进一步证明了L4较L3更能指示植株氮质量分数。李刚华等[8]研究表明:L3可作为较为理想指示叶诊断水稻氮素营养状况,与本研究结果不同。
水稻L1与L4的归一化SPAD指数与水稻冠层叶片氮质量分数存在显著的相关关系,能较好估计水稻冠层叶片氮质量分数,模型不易受环境因素影响,具有较高稳定性。后续研究将建立INDSPAD14与水稻氮营养指数之间的定量关系,用于水稻氮素营养诊断。
-
不同的施氮量的水稻均会表现出2次叶片颜色交替的现象(即SPAD值2次升降),这是其自身内在因素控制的,不同的施氮量无法使这一现象消失。水稻植株缺氮条件下,植株靠近根部的叶位L3和L4的SPAD值小于植株顶部叶片(L2)叶片的SPAD值。随着施氮量的提高,出现了L3和L4的SPAD值大于L2的现象。推测出L3和L4相比L2对氮素营养状况的相关性更高,更适合作为植株氮营养诊断的理想指示叶。
发现INDSPAD14与N0~N4所有不同施氮量组别之间呈显著正相关,‘甬优358’水稻品种决定系数R为0.69~0.96,‘秀水134’品种决定系数R为0.64~0.94,INDSPAD14与植株氮质量分数的定量关系受施氮量和环境的影响较小而保持稳定,用INDSPAD14估算氮质量分数较为准确,顶4叶相较其他叶片更能指示水稻植株氮质量分数。
Characteristics and diagnosis of nitrogen nutrition for rice canopy leaf SPAD value changes
-
摘要: 建立水稻Oryza sativa氮营养诊断模型,实时反映水稻植株氮素营养状况,对水稻田间管理至关重要。于2015年在浙江省德清市开展田间试验,选择‘甬优538’‘Yongyou 538’和‘秀水134’‘Xiushui 134’作为代表品种,设置5个施氮水平0(N0),70.0(N1),140.0(N2),210.0(N3),280.0(N4)kg·hm-2,通过研究不同施氮水平下2个水稻品种冠层叶片作物分析仪器开发值(SPAD)变化规律,探究5个不同施氮水平下植株氮质量分数的变化趋势,并利用归一化SPAD指数(INDSPAD14)估算植株氮质量分数。结果表明:顶4叶相较其他叶片更能指示水稻植株氮质量分数,归一化SPAD指数与N0~N4所有不同施氮量组别之间冠层叶片氮质量分数呈显著正相关(P < 0.05),‘甬优358’水稻品种决定系数为0.69~0.96,‘秀水134’品种决定系数为0.64~0.94。该指数可以对水稻冠层叶片氮质量分数快速估测。Abstract: Nitrogen(N) is important for rice growth. To establish a rice (Oryza sativa) nitrogen nutrition diagnostic model that could reflect the N status of a rice organism and that could be convenient for rice field management, a rice community test was conducted in Deqing City, Zhejiang Province in 2015. Oryza sativa 'Yongyou 538' and 'Xiushui 134' were taken as the representative cultivars. Five levels of N were established:0(N0)(ck), 70.0(N1), 140.0(N2), 210.0(N3), and 280.0(N4) kg·hm-2. Measurement of rice SPAD (soil and plant analyzer development) values were made using a portable SPAD-502 meter and followed by a correlation analysis. Results showed that the fourth leaf from the top was a good indication of plant nutrition status. There was a significant positive correlation between the normalized SPAD index (INDSPAD14) and N content in canopy leaves with the N0-N4 treatments. The coefficients of determination for 'Yongyou 358' r2=0.69-0.96 and for 'Xiushui 134' r2=0.64-0.94. By analyzing the relationship between INDSPAD14 and plant N content, a diagnostic model of rice N concentration was established.
-
Key words:
- botany /
- Oryza sativa /
- canopy leaf /
- SPAD value /
- nitrogen
-
图 1 ‘甬优538’水稻冠层叶片SPAD值在不同的5个施氮水平下的动态变化趋势
Figure 1 Trend of dynamic change of SPAD value of canopy leaf of 'Yongyou 538' rice under different 5 nitrogen fertilizer levels
图 2 ‘秀水134’水稻品种冠层叶片SPAD值在不同的5个施氮水平下的动态变化趋势
Figure 2 Trend of dynamic change of SPAD value of canopy leaf of 'Xiushui 134' rice under different 5 nitrogen fertilizer levels
图 3 不同施氮量水平下‘甬优538’和‘秀水134’水稻冠层叶片SPAD值在的动态变化趋势
Figure 3 Trend of dynamic change of leaf SPAD values of 'Yongyou 538 'and 'Xiushui 134' under different 5 nitrogen fertilizer levels
图 4 不同施氮水平下水稻冠层叶片氮质量分数随移栽后天数的变化趋势
Figure 4 Variation trend of nitrogen concentration of rice leaf under different nitrogen levels with the days after transplantation
图 5 ‘甬优358’水稻INDSPAD14与冠层叶片氮质量分数的定量关系
Figure 5 Quantitative relationship between the N content of leaf and INDSPAD14 in'Yongyou 358'
图 6 ‘秀水134’水稻INDSPAD14与冠层叶片氮质量分数的定量关系
Figure 6 Quantitative relationship between the leaf N content of rice and INDSPAD14 in'Xiushui 134'
表 1 不同时期氮肥用量
Table 1. Nitrogen consumption at different stages
编号 氮肥/(kg·hm-2) 组成/(kg·hm-2) 基肥 蘖肥 穗肥 N0 0 0 0 0 N1 70.0 28.0 14.0 28.0 N2 140.0 56.0 28.0 56.0 N3 210.0 84.0 42.0 84.0 N4 280.0 112.0 56.0 112.0  下载: 导出CSV
下载: 导出CSV
-
[1] ARREGUI L M, LASA B, LAFARGA A, et al. Evaluation of chlorophyll meters as tools for N fertilization in winter wheat under humid Mediterranean conditions[J]. Europ J Agron, 2006, 24(2):140-148. [2] HAWKINS J A, SAWYER J E, BARKER D W, et al. Using relative chlorophyll meter values to determine nitrogen application rates for corn[J]. Agron J, 2007, 99(4):1034-1040. [3] NOURA Z, MARIANNE B, GILLES B, et al. Chlorophyll measurements and nitrogen nutrition index for the evaluation of corn nitrogen status[J]. Agron J, 2010, 100(5):1264-1273. [4] PROST L, JEUFFROY M H. Replacing the nitrogen nutrition index by the chlorophyll meter to assess wheat N status[J]. Agron Sustain Dev, 2007, 27(4):321-330. [5] 李金文.基于水稻叶片生理生态学特征的氮营养诊断[D].杭州: 浙江大学, 2010. LI Jinwen. The Diagnosis of Rice N Status based on Leaf Physioecological Characteristics[D]. Hangzhou: Zhejiang University, 2010. [6] 彭显龙, 刘元英, 罗盛国, 等.实地氮肥管理对寒地水稻干物质积累和产量的影响[J].中国农业科学, 2006, 39(11):2286-2293. PENG Xianlong, LIU Yuanying, LUO Shengguo, et al. Effects of site-specific nitrogen management on yield and dry matter accumulation of rice from cold areas of northeastern China[J]. Agric Sci Sin, 2006, 39(11):2286-2293. [7] 杨虎.水稻冠层叶片氮素分布变化及氮营养状况快速诊断[D].杭州: 浙江大学, 2014. YANG Hu. Changes of Nitrogen Distribution and Rapid Diagnosis for Nitrogen Nutrition Status in the Canopy Leaf of Rice[D]. Hangzhou: Zhejiang University, 2014. [8] 李刚华, 薛利红, 尤娟, 等.水稻氮素和叶绿素SPAD叶位分布特点及氮素诊断的叶位选择[J].中国农业科学, 2007, 40(6):1127-1134. LI Ganghua, XUE Lihong, YOU Juan, et al. Spatial distribution of leaf N content and SPAD value and dselectioon of suitable leaf for N diagnosis in rice[J]. Sci Agric Sin, 2007, 40(6):1127-1134. [9] 凌启鸿, 王绍华, 丁艳锋, 等.关于用水稻"顶3顶4叶叶色差"作为高产群体叶色诊断统一指标的再论证[J].中国农业科学, 2017, 50(24):4705-4713. LING Qihong, WANG Shaohua, DING Yanfeng, et al. Re-evaluation of using the color difference between the top 3rd leaf and the 4th leaf as a unified indicator for high-yielding rice[J]. Sci Agric Sin, 2017, 50(24):4705-4713. [10] 姜继萍, 杨京平, 杨正超, 等.不同氮素水平下水稻叶片及相邻叶位SPAD值变化特征[J].浙江大学学报(农业与生命科学版), 2012, 38(2):166-174. JIANG Jiping, YANG Jingping, YANG Zhengchao, et al. Dynamic characteristics of SPAD value of rice leaf and adjacent leaf under different N application rates[J]. J Zhejiang Univ Agric Life Sci, 2012, 38(2):166-174. [11] 李杰, 冯跃华, 牟桂婷, 等.基于SPAD值的水稻施氮叶值模型构建及应用效果[J].中国农业科学, 2017, 50(24):4714-4724. LI Jie, FENG Yuehua, MOU Guiting, et al. Construction and application effect of the leaf value model based on SPAD value in rice[J]. Sci Agric Sin, 2017, 50(24):4714-4724. [12] 徐富贤, 熊洪, 张林, 等.杂交中稻齐穗后叶片SPAD值衰减对再生力的影响[J].中国农业科学, 2009, 42(10):3442-3450. XU Fuxian, XIONG Hong, ZHANG Lin, et al. Effects of the decreased index of SPAD value of leaf after full heading on ratooning ability[J]. Sci Agric Sin, 2009, 42(10):3442-3450. [13] 肖立中, 李之林, 张建国, 等.前期施氮对二系杂交水稻分蘖及其成穗的影响[J].华南农业大学学报, 1999, 20(3):10-14. XIAO Lizhong, LI Zhilin, ZHANG Jianguo, et al. Effects of nitrogen application during early growth stage on the tillers and the tiller-panicle turnover rate of two-line hybrid rice[J]. J South China Agric Univ, 1999, 20(3):10-14. [14] 姜继萍.水稻冠层叶片SPAD数值变化特征及其在氮素营养诊断中的应用[D].杭州: 浙江大学, 2012. JIANG Jiping. Dynamic Characteristics of SPAD Values of Different Positions Leaves in the Canopy of Rice and Its Application for Nitrogen Nutrition Diagnosis in Rice Production[D]. Hangzhou: Zhejiang University, 2012. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2019.05.014
 点击查看大图
点击查看大图

计量
- 文章访问数: 3889
- HTML全文浏览量: 764
- PDF下载量: 83
- 被引次数: 0
