





-
种群动态是一定时间和空间范围内种群的大小和数量变化规律,是植物个体生存能力与环境相互作用的结果,已成为植物生态学研究的热点之一[1]。种群年龄结构反映了种群在特定时间世代重叠的特征,是分析探索种群动态的有效方法,被广泛应用于种群动态研究中[2]。静态生命表、种群结构动态量化分析法和时间序列预测法是研究种群数量动态变化和进行种群统计的重要工具。青海云杉Picea crassifolia为松科Pinaceae云杉属Picea植物,是贺兰山海拔最高、分布面积最广的顶级森林群落,呈斑块状分布于贺兰山山脊,对当地保持水土、涵养水源、维护生态稳定以及促进经济发展具有重要意义。目前,关于贺兰山青海云杉的研究主要集中于植物区系组成、植物群落类型多样性、针叶林结构和特征、水文特征以及动物区系组成和分布等[3-8],而关于其种群动态的研究鲜有报道。本研究通过对贺兰山青海云杉种群年龄结构的调查研究,探讨贺兰山青海云杉天然林的种群的生长状况和动态规律,以期为贺兰山森林植被的保护管理提供理论依据。
-
在进行充分实地考察和资料论证分析的基础上,确定研究区域为贺兰山苏峪口国家森林公园。研究区域自然概况见文献[4]。外业调查于2011年7-8月进行,后于2012年6月进行补充调查。考虑到青海云杉为贺兰山高海拔地区单一建群种,乔木组成简单(常见乔木仅4种),且林木分布均匀,林下植被稀疏,地形限制较大等因素,确定调查样方大小为10 m × 10 m;采用典型样地法布设样方。共设样方20个(其中青海云杉纯林样地16个,用于种群动态分析;林窗样地4个,用于初始种群调查)。采用相邻网格法,将每个10 m × 10 m样方划分为4个5 m × 5 m小样方进行木本植物调查,分别记录乔木(树高>1.5 m)胸径、树高、冠幅、枝下高,记录灌木(包括乔木幼苗,树高<1.5 m)种类、数量、高度、基径和冠幅等;草本记录种类和频度、盖度等。群落因子调查包括海拔、坡度、坡向、坡位、郁闭度、土壤厚度、枯枝落叶层厚度、人为干扰程度等。
-
种群年龄结构是种群的重要特征,是分析探索种群动态的有效方法[1]。青海云杉种群的年龄与胸径具有显著的相关性,因此,可以用胸径代替种群个体的大小[9-10]。树高<1.5 m的幼苗、幼树,大小径级为1,树高≥1.5 m的按2 cm径阶,划为2,4,6,8,…,28径阶的大小级,划分2~14个大小级,共14个大小级,分析种群结构。结合林业苗木生产过程中的惯例,又可将青海云杉划分为4个年龄阶段:幼苗、幼树阶段,树高≤1.5 m(对应于1级);小树阶段,胸径2~10 cm(2~5级);中树阶段,12~20 cm(6~10级);大树阶段,胸径>22 cm(11~14级)。其中,幼苗和幼树为青海云杉种群的主要更新苗木。
-
采用陈晓德[11]的量化方法定量描述贺兰山青海云杉的种群动态。量化方法如下:
$${{V}_{n}}=\frac{{{S}_{n}}-{{S}_{n+1}}}{\max ({{S}_{n}},{{S}_{n+1}})}\times 100%;{{V}_{pi}}=\frac{1}{\sum\limits_{n=1}^{k-1}{{{S}_{n}}}}\sum\limits_{n=1}^{k-1}{({{S}_{n}}{{V}_{n}})}。$$ 考虑未来干扰时:
$$V{{'}_{pi}}=\frac{\sum\limits_{n=1}^{k-1}{({{S}_{n}}{{V}_{n}})}}{K\min ({{S}_{1}},{{S}_{2}},{{S}_{3}},\cdots {{S}_{k}})\sum\limits_{n=1}^{k-1}{{{S}_{n}}}}。$$ (1) 式(1)中:k为种群年龄级数量,Sn,Sn+1分别表示第n与第n+1年龄种群个体数,Vn表示种群相邻级间(n与n+1级)个体数量变化动态指数,Vpi表示整个种群结构数量变化动态指数,V′pi 表示在考虑未来外部干扰条件下的种群结构数量变化动态指数,V′pi ,Vpi和Vn取正、负、零值的意义分别反映种群或相邻年龄级个体数量的增长、衰退、稳定的动态关系。
-
以青海云杉纯林数据为基础,以其最大种群密度值为初始种群数(a0),将各径级实际存活数换算成密度(株·hm-2),编制静态生命表。通过实测值ax计算各项,具体计算如下[9, 12-13]:
$$\begin{align} & {{l}_{x}}={{a}_{x}}l{{a}_{0}}\times 1000;{{d}_{x}}={{l}_{x}}-{{l}_{x+1}};{{q}_{x}}={{d}_{x}}/{{l}_{x}};{{L}_{x}}= \\ & \left( {{l}_{x}}+{{l}_{x+1}} \right)/2;{{T}_{x}}=\sum\limits_{x}^{\infty }{{{L}_{x}}};{{e}_{x}}={{T}_{x}}/{{l}_{x}};{{K}_{x}}=\ln {{l}_{x}}-{{l}_{x+1}}。 \\ \end{align}$$ (2) 式(2)中:x为单位时间年龄等级的中值;ax为x龄级开始时的实际存活个体数;ax′为匀滑后的单位面积(hm2)存活个体数;lx为x龄级开始时的标准化存活数,将各年龄间隔中存活量最大的株数标准化为1 000;dx为从x到x+1期间标准化死亡个体数,即标准化死亡量; qx为各龄级的个体死亡率;Lx为从x到x+1期的平均存活个体数;Tx为x年龄至超过x年龄的总个体数或种群中活到x年龄的所有个体的剩余总寿命;ex为进入x龄级个体的平均生命期望,可以理解为平均每个树木在现有年龄之后还能存活年龄间隔期的倍数;lnlx为死亡率的自然对数,常用于绘制存活曲线;Kx为消失率。以生命表的标准化存活数(lx)和标准化死亡量(dx)为纵坐标,大小级为横坐标(x),绘制青海云杉种群的存活曲线[12]。引入4个种群生存分析函数[1]进行种群生命动态分析,即S(t):生存率函数;F(t):积累死亡率函数;f(t):死亡密度函数;λ(t):危险率函数。计算公式如下:
$$\begin{align} & \hat{S}={{{\hat{p}}}_{1}}{{{\hat{p}}}_{2}}\cdots {{{\hat{p}}}_{i}}({{{\hat{p}}}_{i}}为存活频度);{{{\hat{F}}}_{i}}=1-{{{\hat{S}}}_{i}}; \\ & \hat{f}({{t}_{i}})=\frac{{{{\hat{S}}}_{i-1}}{{{\hat{S}}}_{i}}}{{{h}_{i}}}=\frac{{{{\hat{S}}}_{i-1}}{{{\hat{q}}}_{i}}}{{{h}_{i}}}\left( {{h}_{i}}为区间长度,{{{\hat{q}}}_{i}}为死亡频率 \right); \\ & \hat{\lambda }({{t}_{i}})=\frac{2{{{\hat{q}}}_{i}}}{{{h}_{i}}\left( 1+{{{\hat{p}}}_{i}} \right)}。 \\ \end{align}$$ (3) -
采用时间序列分析的一次移动平均法[1, 13]对青海云杉种群龄级结构进行模拟和预测,其模型如下:
$$M_{t}^{\left( 1 \right)}=\frac{1}{n}\sum\limits_{k=t-n+1}^{t}{{{x}_{k}}}。$$ (4) 式(4)中:n为需要预测的未来时间年限,t为龄级,是近期n个观测值在t时刻的平均值,成为第n周期的移动平均,即表示未来n年时t龄级的种群的大小,xt表示当前t龄级的种群的大小。根据所调查青海云杉种群大小级。本研究中,分别取种群经历2,4,6,8个大小级的时间后,对种群未来发展趋势进行预测。
-
青海云杉是贺兰山区高海拔地区分布面积最大的乔木优势种,种群最大密度为8 500株·hm-2,最小密度1 425株·hm-2,平均密度3 900株·hm-2,70%的样地低于平均密度,密度分布不均匀。不同林分林下幼苗更新存在差异(表 1)。青海云杉被破坏后形成林窗,在水热条件较好的东南坡和平缓地带,多衍生出次生山杨林和次生灌丛,林窗青海云杉天然更新良好,种群密度极大,通常可以自然向青海云杉纯林发展。青海云杉纯林是贺兰山青海云杉种群的主体部分,幼苗天然更新能力较差,不足林窗更新的三分之一。这说明青海云杉的天然更新强弱与林分类型有关,林窗能够为青海云杉的种子萌发和幼苗生长提供较好的条件,促进其生长发展。
表 1 贺兰山青海云杉种群天然更新特征
Table 1. Characteristic of seedling regeneration of Picea crassifolia population in Helan Mountain
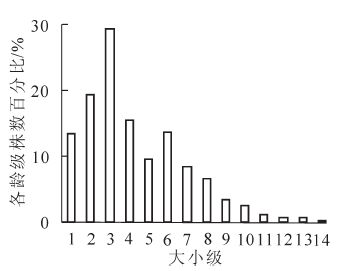
林分 海拔范围/m 坡向 坡度/(°) 土壤厚度/cm 郁闭度 幼亩幼树平均密度/(株·hm-2) 林窗 2 250~2 300 E,NW 2~15 45 0.1~0.6 6 700 非林窗 2 350~2 800 N,NE,NW 17~35 65 0.7 2 700 按照14个大小级划分,绘制贺兰山青海云杉种群年龄结构图(图 1)。贺兰山自然保护区青海云杉种群年龄结构整体呈左偏态分布,低径阶植株较多,高径阶植株偏少,其中幼苗幼树(1级)92株,占总体的13.45%;小树(2~5级)503株,占整体的59.24%;中树237株,占整体的27.91%;大树17株,占整体的2.02%。说明贺兰山青海云杉种群主体处于小树和中树阶段,林分基本郁闭,种群具有较大的发展潜力,为增长型种群,但1,2,5级植株偏少,幼苗天然更新较差,种群可能发生衰退。种群结构动态量化结果显示,各径级间个体数量变化动态指数Vn分别为-43.48%,-51.52%,47.00%,38.68%,-44.62%,38.30%,22.41%,48.89%,26.09%,58.82%,42.86%,-25.00%和80.00%,其中V1和V2皆小于0,说明林下幼苗缺少,种群天然更新较差;V5和V12小于0,说明青海云杉种群在发展中具有波动性;整个种群年龄的动态指数为Vpi=0.010 7;在考虑外部干扰的情况下为Vpi′=0.001 5>0,表明种群趋近于稳定型,具有一定的增长性,但增长较为缓慢。

图 1 贺兰山青海云杉种群年龄结构
Figure 1. Age structure of Picea crassifolia population in Helan Mountain
-
对于长寿命多年生植物种群,应用“空间差异代替时间变化”的方法来研究其动态具有实际意义,年龄结构和静态生命表以及种群动态预测都是基于现有调查数据,能够反映种群的基本属性,结果符合种群的自然动态规律[1]。由于很难追踪乔木树种的全部生活史,静态生命表常采用同一时期收集的种群所有个体径级进行编制,是由“空间推导时间”,所以所得数据并不满足编制生命表的3个假设,因而生命表分析中常产生死亡率为负的情况。对此,Wretten等[13]认为,生命表中产生的负值虽然与数学假设不符,但仍能提供有用的生态学记录,表明种群并非静止不动,而是在发展或者衰退中。对此许多学者[1, 14-15]采取匀滑技术对数据进行处理。本研究亦采用相同方法,对2~10级、11~14级2个阶段数据进行匀滑处理后,绘制静态生命表(表 2)。
表 2 贺兰山青海云杉种群静态生命表
Table 2. Static life table of Picea crassifolia population in Helan Mountain
x 径级中值 ax ax′ lx dx qx Lx Tx ex lnlx Kx 1 1 92 8 500 1 000 743 0.743 629 1 420 1.42 6.908 1.357 2 3 132 2 188 257 30 0.116 243 915 3.55 5.551 0.123 3 5 200 1 935 228 30 0.131 213 766 3.36 5.428 0.140 4 7 106 1 682 198 30 0.150 183 617 3.12 5.288 0.163 5 9 65 1 429 168 30 0.178 153 468 2.78 5.125 0.196 6 11 94 1 175 138 30 0.216 123 329 2.38 4.929 0.244 7 13 58 921 108 30 0.275 93 214 1.97 4.685 0.321 8 15 45 668 79 30 0.379 64 127 1.61 4.364 0.476 9 17 23 415 49 30 0.610 34 69 1.40 3.888 0.941 10 19 17 162 19 10 0.512 14 37 1.95 2.948 0.718 11 21 7 79 9 2 0.228 8 23 2.47 2.229 0.259 12 23 4 61 7 1 0.148 7 15 2.06 1.971 0.160 13 25 5 52 6 1 0.173 6 8 1.33 1.811 0.190 14 27 1 43 5 5 - 3 3 0.50 1.621 - 由种群静态生命表(表 2)可以看出,随着年龄的增加,青海云杉种群的死亡数(dx)大致呈逐渐减小的趋势,1~6级的死亡数(dx)较高,特别是1级死亡数最大。平均存活数(Lx)和总存活数(Tx)稳定减少,生命期望值(ex)1级较小,2级最大,随后缓慢下降,10级时突然升高,而后下降逐渐减小;消失率(Kx)1级最大,2级以后呈波动上升,至9,10级出现1个小峰值,至11级迅速降低。青海云杉种群的死亡数、生命期望和消失率分别在1级(树高<1.5 m)和9~10级出现异常,说明这2个阶段种群中个体通过环境筛选的压力增大。
-
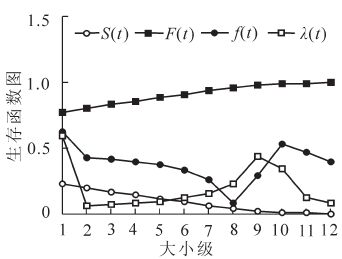
依据静态生命表数据计算种群各生存函数值(图 2)。生存函数稳定降低,积累死亡率稳定升高;危险率函数与种群密度死亡函数的表现具有一定的相似性,两者的最大值都出现在1级,1~2级曲线较陡,至2~8级趋于平缓,9~12级开始出现较大波动。其中,危险率函数在2~8级和10~12级呈减小趋势,低谷值出现于8级,高峰值出现于9级;种群密度函数从2~9级缓慢升高,9~12级快速下降,峰值出现于9级。青海云杉种群在2级、9~10级2个阶段数据的异常表现说明环境筛选强度增大。死亡曲线和消失率表现相似(图 3),皆为单峰双谷曲线,高峰值出现于1级和9级,分别为0.743,1.357和0.610,0.941;低谷值出现于2,12和14级,且2~7级、10~12级的曲线斜率较小,说明这2个阶段种群较为稳定。其结果与种群死亡密度和危险率表现相一致。这个结果与丁国民等[9]研究甘肃青海云杉种群的结论稍有差异,其原因可能与种群年龄和生境条件有关。此外,4个生存函数结果表明贺兰山青海云杉种群在发展过程中具有波动性,具有前后期波动,中期稳定的特点(图 2~3)。

图 2 青海云杉种群生存函数图
Figure 2. Survival functional rate of Picea crassifolia population

图 3 青海云杉死亡曲线(qx)和消失率曲线(Kx)函 数曲线
Figure 3. Function cure of mortality curve (qx) and disappearance rate (Kx) of P. crassifolia population
-
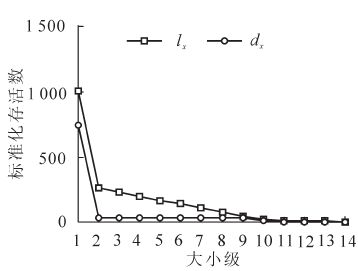
本研究参照丁国民等[9]和吴承祯等[15]绘制存活曲线的方法,绘制贺兰山青海云杉存活曲线和死亡曲线(图 4)。从种群存活曲线(图 4)可以看出:1级个体成活率较低,只有大约20%的幼苗能穿过此筛选活到2级,其原因是10 年生(树高<1.5 m)以下幼苗生长缓慢,种内竞争较弱;20 年生(树高≥1.5 m)后生长加速,种内竞争加剧,使得只有少量个体通过环境筛选进入下一个生长阶段。由于1级苗木的“自疏”作用,单位面积内植株个体明显减小,环境条件基本可以满足所有个体对于光温水气、空间、养分等的需求,种群死亡量降低,成活率逐渐稳定。种群存活曲线介于DeeveyⅡ型和DeeveyⅢ型之间。采用幂函数方程式Nx=N0e-bx和指数函数方程式Nx=N0e-bx对DeeveyⅡ型和Ⅲ型存活曲线进行检验,得到2组方程,Nx=32.910x-0.419,F=54.646,R2=0.820;Nx=9.295e-0.003x,F=40.950,R2=0.773。对2组方程的检验结果显示,幂函数方程的F值和R2值都大于指数函数说明青海云杉的存活曲线趋近于DeeveyⅡ型。

图 4 贺兰山青海云杉种群的存活曲线(lx)和死亡曲线(qx)
Figure 4. Survival curve and mortality curve (qx) of Picea crassifolia population in Helan Mountain
-
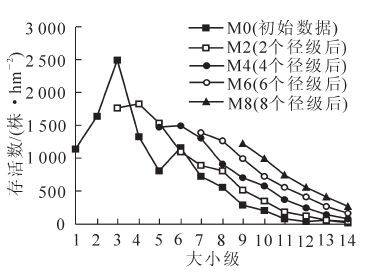
时间序列是以实地调查数据,按一次移动平均法[14]对种群未来个体数做一个粗略评估。将各径级数据换算成株·hm-2,绘制贺兰山青海云杉种群2,4,6,8个径级时间后的径级与个体数的关系图(图 5)。从同一时间看,各曲线数值随径级增加而大致表现为减少趋势。随着时间的推移(从M2到M8),除2个径级后(M2)曲线表现稍有波动外,青海云杉种群其余各径级数量皆表现为单调增加趋势。从M0(初始数据)到M2(2个径级后),3级对应存活数减少明显,7~14级对应存活数均低于初始数据(M0)。而M4到M8(4到8个径级后),6级以上个体数量皆大于M0相应径级数量。因此,时间序列预测结果表明贺兰山青海云杉种群具有一定的增长能力和恢复能力,与种群动态量化结果一致。可以推断,如果能维持种群现有的环境状况,没有人为干扰,青海云杉种群将在一定时期内保持相对稳定的种群数量,而加强青海云杉林封育保护,将更有利种群生长发育,能够促进种群的恢复发展。

图 5 贺兰山青海云杉2, 4,6,8个龄级后的时间序列预测
Figure 5. Time sequence prediction in the quantitative dynamics of P. crassifolia population
-
青海云杉多生长在阴坡和半阳坡上,耐寒耐旱耐瘠薄,是优良的水土保持树种和经济用材,对贺兰山经济发展和生态保护具有重要意义。青海云杉种群密度分布非常不均匀,天然更新主要发生在郁闭度较小的山杨林、林窗和林隙中,尤以郁闭度为0.4的山杨林下,种群密度最大,可达8 500株·hm-2,青海云杉纯林天然更新较差。青海云杉的这些特点是主要是由青海云杉纯林内生境条件和其本身生物学特性所决定的。其一,青海云杉林具有较高的草本类盖度,这导致青海云杉种子无法接触到土壤而难以萌发;其二,虽然青海云杉幼苗有一定的耐阴性,但小树生长却需要足够的光照条件。青海云杉林内较差的光照条件,影响了小树的生长使它们具有较高死亡率,从而导致了这种稳定型种群中幼苗数量稀少的现象。但是由于青海云杉种子储量巨大,一旦林窗或林隙为它们创造适宜条件,种子就可以大量萌发,迅速占据林间空地,使得成熟型和衰退型青海云杉种群可以自然更新并修复自己的种群结构,使种群结构又趋于稳定型,因此青海云杉可以高概率的维持自己在群落中的优势地位,长期保持稳定。
贺兰山青海云杉种群的主体为小树和中树,约占整体的82.4%,而幼苗幼树和高径阶个体数量偏少;Vpi′=0.001 5>0,说明贺兰山青海云杉种群处于增长型向稳定型的过渡阶段,种群生长过程中虽有波动,但并不易导致衰退。关于此点,可通过对贺兰山青海云杉长期定点观测作进一步的证明。存活曲线趋近于DeeveyⅡ型,4个生存函数在2级、8~10级出现异常波动,说明贺兰山青海云杉在发展过程中具有2次“自疏”现象。第1次发生在青海云杉林底层,为幼苗幼树时期,主要表现为种群个体对光温水气、养分、空间的需求与自然供给的矛盾,种内竞争激烈,80.0%以上个体在此阶段被淘汰;第2次发生在林冠层,竞争主体为中树,表现为个体对于空间的争夺,死亡量较低。总的来说,贺兰山青海云杉林仍处于增长发展阶段,趋近于稳定阶段,大部分林木未达到采伐标准,仍需要进一步的封育保护。因此,禁止滥砍滥伐,继续封山育林,以及加强幼苗抚育工作,对于青海云杉的保护和发展具有重要意义。贺兰山青海云杉林广泛采用的清理林下枯立灌木的抚育方法虽然能为青海云杉幼苗提供更多的生存空间和光照条件,但过度清理林下灌木也会不利于野生动物的栖息生存,大大降低当地生物多样性,不利于整个生态系统的稳定。因此,我们建议根据青海云杉群落特性和已有经验,采用清理枯病木、采伐成熟林、制造林窗,以及加强林隙和林窗的幼苗抚育等管理措施,将更有利于青海云杉的天然更新和种群的稳定发展。
Population dynamics of Picea crassifolia on the eastern slope of Helan Mountain
-
摘要: 采用典型样地法调查了贺兰山东坡苏峪口国家森林公园青海云杉Picea crassifolia种群,分析了种群年龄结构,量化了种群结构动态,绘制了静态生命表和时间序列模型,以探讨种群数量动态变化特征和规律。结果表明:研究区域内青海云杉种群主体为小树和中树(占总数的82.4%),幼苗幼树和大树数量偏少。在考虑未来外部干扰情况下,种群年龄结构动态量化指数为V'pi=0.001 5>0,存活曲线趋近于DeeveyⅡ型,死亡率高峰分别出现在1级和9级,说明贺兰山青海云杉种群正处于增长型向稳定型的过渡阶段,种群具有2次"自疏"过程,分别发生在幼苗时期和中树向大树过渡时期。虽然贺兰山青海云杉种群具有较强的自我更新能力和恢复能力,并不易导致衰退,但该种群整体径级较小,天然更新较差,仍需要长期的封育保护。Abstract: To determine characteristics and regularity in variation for population dynamics,a typical sample plot method was used for the Picea crassifolia population of Suyukou National Park located on the eastern slope of Helan Mountain. Then,based on all the data collected from 20 plots(10 m×10 m),population age structure was analyzed,population structure types were quantified, and static life table and time sequence models were drawn. Results of population age structure showed that the P. crassifolia population consisted mostly of small and medium-sized trees(82.4%),while seedlings,saplings,and mature trees were scarce; when future external interference from the time sequence model was taken into account, the population age structure dynamic quantitative index was V'pi=0.001 5>0,the survival curve leveled off at Deevey-Type Ⅱ,and the mortality rate peaked at 1 and 9 age-class. This indicated that the P. crassifolia population in Helan Mountain was in a transitory period between growth and steady-state forms;however,despite its self-renewal and resilience capabilities, with a small diameter class population and weak natural regeneration of saplings,a long protection period to facilitate afforestation is needed.
-
Key words:
- forest ecology /
- Picea crassifolia /
- Helan Mountain /
- population dynamics
-
图 1 贺兰山青海云杉种群年龄结构
Figure 1 Age structure of Picea crassifolia population in Helan Mountain
图 3 青海云杉死亡曲线(qx)和消失率曲线(Kx)函 数曲线
Figure 3 Function cure of mortality curve (qx) and disappearance rate (Kx) of P. crassifolia population
图 4 贺兰山青海云杉种群的存活曲线(lx)和死亡曲线(qx)
Figure 4 Survival curve and mortality curve (qx) of Picea crassifolia population in Helan Mountain
图 5 贺兰山青海云杉2, 4,6,8个龄级后的时间序列预测
Figure 5 Time sequence prediction in the quantitative dynamics of P. crassifolia population
表 1 贺兰山青海云杉种群天然更新特征
Table 1. Characteristic of seedling regeneration of Picea crassifolia population in Helan Mountain
林分 海拔范围/m 坡向 坡度/(°) 土壤厚度/cm 郁闭度 幼亩幼树平均密度/(株·hm-2) 林窗 2 250~2 300 E,NW 2~15 45 0.1~0.6 6 700 非林窗 2 350~2 800 N,NE,NW 17~35 65 0.7 2 700  下载: 导出CSV
下载: 导出CSV
表 2 贺兰山青海云杉种群静态生命表
Table 2. Static life table of Picea crassifolia population in Helan Mountain
x 径级中值 ax ax′ lx dx qx Lx Tx ex lnlx Kx 1 1 92 8 500 1 000 743 0.743 629 1 420 1.42 6.908 1.357 2 3 132 2 188 257 30 0.116 243 915 3.55 5.551 0.123 3 5 200 1 935 228 30 0.131 213 766 3.36 5.428 0.140 4 7 106 1 682 198 30 0.150 183 617 3.12 5.288 0.163 5 9 65 1 429 168 30 0.178 153 468 2.78 5.125 0.196 6 11 94 1 175 138 30 0.216 123 329 2.38 4.929 0.244 7 13 58 921 108 30 0.275 93 214 1.97 4.685 0.321 8 15 45 668 79 30 0.379 64 127 1.61 4.364 0.476 9 17 23 415 49 30 0.610 34 69 1.40 3.888 0.941 10 19 17 162 19 10 0.512 14 37 1.95 2.948 0.718 11 21 7 79 9 2 0.228 8 23 2.47 2.229 0.259 12 23 4 61 7 1 0.148 7 15 2.06 1.971 0.160 13 25 5 52 6 1 0.173 6 8 1.33 1.811 0.190 14 27 1 43 5 5 - 3 3 0.50 1.621 -
下载: 导出CSV
-
[1] 刘贵峰,臧润国,成克武,等. 天山云杉的种群动态[J]. 应用与环境生物学报,2011,17(5):632-637. LIU Guifeng,ZANG Runguo,CHENG Kewu,et al. Population dynamics of Picea schrenkiana var. tianschanica in the Tianshan Mountains[J]. Chin J Appl Enciron Biol,2011,17(5):632-637. [2] 胡尔查,王晓江,刘永洪,等. 乌拉山自然保护区油松种群结构与分布格局研究[J]. 林业科学研究,2011,24(3):363-369. HU Eecha,WANG Xiaojiang,LIU Yonghong,et al. Population structure and distribution pattern of Pinus tabulaeform is in Wulashan Natural Reserve of Inner Mongolian[J]. For Res,2011,24(3):363-369. [3] 梁存柱,朱宗元,王炜,等. 贺兰山植物群落类型多样性及其空间分异[J]. 植物生态学报, 2004,28(3):361-368. LIANG Cunzhu,ZHU Zongyuan,WANG Wei,et al. The diversity and spatial distribution of plant communities in the Helan Mountains[J]. Acta Phytoecol Sin,2004,28(3):361-368. [4] 朱源,康慕谊,刘全儒,等. 贺兰山针叶林结构特征与种类组成的比较[J]. 地理研究,2007, 26(2):305-312. ZHU Yuan,KANG Muyi,LIU Quanru,et al. Comparison of forest structurer and floristic compositon of coniferous forest in Helan Mountain[J]. Geogr Res,2007,26(2):305-312. [5] 朱源,康慕谊,刘全儒,等. 贺兰山油松和青海云杉交错区的物种丰富度研究[J]. 植物资源与环境学报, 2007, 16(3):1-6. ZHU Yuan,KANG Muyi,LIU Quanru,et al. Study on species richness in ecotone between Pinus tabulaeformis and Pices crassifolia on Helan Mountain[J]. J Plant Resour Environ,2007,16(3):1-6. [6] 朱源,康慕谊,江源,等. 贺兰山木本植物群落物种多样性的海拔格局[J]. 植物生态学报, 2008,32(3):574-581. ZHU Yuan,KANG Muyi,JIANG Yuan,et al. Altitudinal pattern of species diversity in woody plant communities of Mountain Helan[J]. J Plant Ecol,2008,32(3):574-581. [7] 赵春晓,刘建军,任军辉,等. 贺兰山4种典型森林类型凋落物持水性能研究[J]. 水土保持研究,2011,18(2):107-111. ZHAO Chunxiao,LIU Jianjun,REN Junhui,et al. Study on water-holding capacity of litter in four forest types of Helan Mountain[J]. Res Soil & Water Conserv,2011,18(2):107-111. [8] 杨贵军,于有志,王新谱. 宁夏贺兰山拟步甲科的区系组成与生态分布[J]. 宁夏大学学报:自然科学版,2011,32(1):67-72. YANG Guijun,YU Youzhi,WANG Xinpu. Tendbrionidae fauna and ecological distribution in Ningxia Helan Mountain Nature Reserve[J]. J Ningxia Univ Nat Sci Ed,2011,32(1):67-72. [9] 丁国民,刘兴明,倪自银,等. 甘肃祁连山青海云杉种群数量动态的初步研究[J]. 植物资源与环境学报, 2005,14(4):36-41. DING Guomin,LIU Xingming,NI Ziyin,et al. A preliminary study of population dynamics of P. crassifolia in Qi lian Mountains[J]. J Plant Resour Environ,2005,14(4):36-41. [10] 刘建泉,丁国民,郝虎,等. 青海云杉群落特征和动态的研究[J]. 西北林学院学报,2008, 23(1):14-17. LIU Jianquan,DING Guomin,HAO Hu,et al. Structure and dynamic of P. crassifolia community in Qilian Mountains Foresst Area[J]. J Northwest For Univ,2008,23(1):14-17. [11] 陈晓德. 植物种群与群落结构动态量化分析方法研究[J]. 生态学报,1998,18(2):214-217. CHEN Xiaode. A study on the method of quantitative analysis for plant population and community structural dynamics[J]. Acta Ecol Sin,1998,18(2):214-217. [12] 张雷,王晓江,胡尔查,等. 科尔沁沙地蒙古黄榆种群结构与空间分布格局[J].中国沙漠, 2011,31(1):115-120. ZHANG Lei,WANG Xiaojiang,HU Ercha,et al. Population structure and spatial pattern of Ulmus macrocarpa var. mongolica in Horqin Sandy Land[J]. J Desert Res,2011,31(1):115-120. [13] WRETTEN S. Field and Laboratory Exercises in Ecology[M]. London:Edward and Arnad Publishers Limited,1980:44-63. [14] 李晓笑,王清春,崔国发,等. 濒危植物梵净山冷杉野生种群结构及动态特征[J]. 西北植物学报,2011,31(7):1479-1486. LI Xiaoxiao,WANG Qingchun,CUI Guofa,et al. Structure and dynamics characteristics of a wild population of endangered Abies fanjingshanensis[J]. Acta Bot Boreal-Occident Sin,2011, 31(7):1479-1486. [15] 吴承祯,洪伟,谢金寿,等. 珍稀濒危植物长苞铁杉种群生命表分析[J]. 应用生态学报, 2000,11(3):333-336. WU Chengzhen,HONG Wei,XIE Jinshou,et al. Life table analysis of Tsuga longibracteata population[J]. Chin J Appl Ecol,2000,11(3):333-336. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2014.01.008
 点击查看大图
点击查看大图
计量
- 文章访问数: 4156
- HTML全文浏览量: 917
- PDF下载量: 608
- 被引次数: 0
