





-
苗木品质是影响造林效果的首要因子,选择品质高的苗木不仅能够提高造林成活率,而且能达到良好的造林效果[1]。施肥是改善苗木品质,提高苗木自身竞争力的重要手段。氮素是植物生长所必需的矿质营养元素之一,它可以调控植物的生长发育,影响植物的光合作用,是组成各种重要有机化合物不可或缺的部分,氮素需求一直是研究热点。目前,常采用的施肥方法主要有常规施肥(CF)和指数施肥(EF)。常规施肥是指在生长期内重复施用同样剂量的肥料。指数施肥则是根据植物在各生长阶段对养分的需求规律,采用指数递增的养分添加方式的一种施肥方法[2-3]。该方法可诱导植株稳态奢侈养分消耗,有效提高苗木体内养分载荷,增强苗木的竞争能力,从而使苗木更好地适应造林地的立地条件[4-5],相对于常规施肥,指数施肥培育的苗木成活率更高,生长效果更好[6],也能避免因肥料过量施用造成的土壤污染。目前,国外用于指数施肥研究的树种有黑云杉Picea mariana[7],日本落叶松Larix kaempferi[8],西铁杉Tsuga heterophylla[9]和花旗松Pseudotsuga menziesii[10]等,且研究成果已经达到应用水平,指数施肥已逐渐成为国外苗木培育的主要施肥方式。近年来,国内利用稳态养分理论对栓皮栎Quercus variabilis[11],山桃稠李Padus maackii[12],楸树Catalpa bungei[13],檀香Santalum album[14]和红楠Machilus thunbergii[15]等树种开展了指数施肥的研究。杉木Cunninghamia lanceolata是中国南方16省区最重要的造林树种之一,是中国特有的速生商品树种,生长快,材质好。与实生苗相比,采用杉木优良无性系造林,林相整齐,林分生长一致,便于集约经营,其产量可增加58.7%[16],且无性系的木材材性均匀一致,收获期一致,是营造工业用材林的最佳材料[17]。因此,无性系造林已逐渐成为杉木造林的主要方法,而无性系优质苗木是成功营造杉木人工林的基础。指数施肥可有效促进杉木实生苗的生长,改善苗木的营养状态[18],推测指数施肥对杉木无性系苗木可能有相似的效果,但目前指数施肥对杉木无性系苗育苗的效果尚不清楚。本研究以1年生杉木无性系苗为研究对象,设置不同的氮素施入方式和用量,在盆栽条件下研究在一个生长季内杉木无性系苗的生长和氮养分的积累对不同施肥处理的响应,旨在揭示杉木无性系幼苗对氮素的需求规律,确定适宜的施氮量,为杉木无性系育苗的营养管理提供参考。
-
试验地设在浙江省临安市浙江农林大学实验基地的温室大棚(30°16′ N,119°44′E)。该地属于中亚热带季风气候,温暖湿润,雨水充沛。年均日照时长为1 847.3 h,年均降水量为1 628.6 mm,年均气温为16.4 ℃,1月均温3.8 ℃,7月均温28.6 ℃,极端最高气温40.4 ℃,极端最低气温-9.2 ℃,年无霜期约为250 d。棚内温度为23.0~33.0 ℃,湿度为55%~75%。
-
试验苗为浙江省开化县林场培育的“开4 ×那11”杂交后代1号优良单株,选取生长基本一致的1年生杉木无系性扦插苗300株,苗高(25.50±0.30)cm,地径(3.12±0.05)mm。2014年3月1日采用23 cm × 24 cm × 28 cm(底径×上口径×高)的花盆,栽植1株·盆-1。为了防止水肥流失,每个花盆配有塑料托盘,供试土壤为花岗岩发育的红壤土类,pH 6.6,有机碳为4.8 g·kg-1,碱解氮为22.7 mg·kg-1,有效磷为0.9 mg·kg-1,速效钾为55.5 mg·kg-1,装土壤10 kg·盆-1。
-
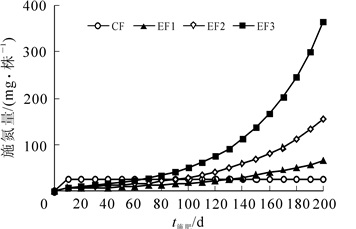
盆栽试验采用完全随机区组设计,共设对照,常规施肥(CF),指数施肥(EF1,EF2,EF3)共5个处理,氮素施用总量分别为0,0.5,0.5,1.0,2.0 g·株-1,重复3次·处理-1,苗木20株·重复-1。待缓苗至3月21日进行第1次施肥,以后隔10 d施氮肥1次,共施20次。常规施肥每次施氮量均为25 mg·株-1,指数施肥则以指数施肥模型[2]来确定每次的施氮量:Nt = Ns(ert-1)-Nt-1,NT = Ns(ert-1)。其中:Nt为在氮素增加率r下第t次的施氮量,Ns为苗木的初始氮含量,Nt-1为包括第t-1次施肥在内的施氮总量,NT为t次施氮后苗木最终的氮含量(假设肥效为100%)。杉木幼苗的初始氮含量为72.19 mg·株-1。氮肥为质量分数46%的尿素,具体施肥进度见图 1。五氧化二磷为质量分数14%的过磷酸钙(7.0 g·株-1)和氧化钾为50%的硫酸钾(2.0 g·株-1)均作底肥在第1次施肥时施入。施肥方法:过磷酸钙和硫酸钾是用小铲在离植株5 cm根圈挖取2 cm深的环沟,然后等量施入后埋平。尿素按不同处理溶于水中,然后用量筒对同一处理的苗木进行等量浇灌。

图 1 不同处理的施肥进度
Figure 1. Different treatments of fertilization
-
生物量测定:施肥处理210 d后,分别取不同处理的苗木各5株收获全株,重复3次,用游标卡尺和卷尺测量苗木地径及苗高。然后用清水洗净,再用去离子水润洗。按根、茎、叶分别剪下,装入信封中,置于烘箱内,105 ℃杀青30 min,70 ℃烘48 h至恒量,用电子天平测其干质量,即为生物量。总生物量(g·株-1)=根生物量+茎生物量+叶生物量。养分测定:称量后的样品粉碎过0.5 mm筛,用元素分析仪(德国Elementer,VARIO Macro)测定氮质量分数。氮积累量(mg·株-1)=氮质量分数(g·kg-1)×相应生物量。
-
采用SPSS 20.0对数据进行描述统计和正态检验,然后进行单因素方差分析,并用最小显著性差异(LSD)法进行多重比较。
-
施肥促进了杉木苗高和地径的生长(图 2)。与对照相比,4个施肥处理的杉木苗高增长34.1%~60.5%,地径增粗20.5%~40.4%,差异均达显著水平(P<0.05)。与常规施肥(CF)相比,指数施肥处理苗高增长了4.1%~19.8%,地径增粗了4.6%~19.8%,苗高和地径均以EF2处理为最佳,显著高于常规施肥(CF)(P<0.05)。

图 2 不同施肥处理杉木幼苗苗高和地径
Figure 2. Plant height and ground diameter of Chinese firunder different fertilization treatments
-
如表 1所示:不同处理根生物量为10.14~14.13 mg·株-1,差异并不显著(P>0.05)。4个施肥处理的茎、叶和全株生物量与对照相比,增加了113.9%~166.9%,158.0%~230.9%,96.4%~128.9%,差异达显著水平(P<0.05);其中指数施肥EF2处理的茎、叶和全株生物量最高,与常规施肥(CF)相比,分别增加12.2%,27.1%,13.9%。
表 1 不同施肥处理杉木不同器官生物量
Table 1. Biomass of Chinese fir under different fertilization treatments
施肥处理 根/(g·株-1) 茎/(g·株-1) 叶/(g·株-1>) 全株/(g·株-1) 对照 10.14 ± 2.84 a 5.33 ± 1.58 b 7.68 ± 2.11 b 23.15 ± 6.40 b 常规施肥CF 13.84 ± 3.33 a 12.68 ± 1.92 a 19.99 ± 4.79 a 46.51 ± 9.35 a 指数施肥EF1 14.13 " 2.24 a 12.16 ± 0.47 a 19.81 ± 1.86 a 46.10 ± 4.08 a 指数施肥EF2 13.35 ± 2.29 a 14.23 ± 3.61 a 25.41 ± 2.99 a 52.99 ± 8.85 a 指数施肥EF3 12.74 ± 1.38 a 11.40 ± 3.16 a 21.32 ± 2.82 a 45.45 ± 6.97 a -
如表 2所示:不同施肥处理杉木幼苗叶的氮质量分数大小为EF3(23.36 g·kg-1)>EF2(17.45 g·kg-1)>EF1(17.22 g·kg-1)>CF(14.00 g·kg-1)>对照(9.92 g·kg-1)。常规施肥(CF)处理比对照高41.1%;指数施肥EF处理比对照高73.6%~135.5%,显著高于CF处理,其中EF3处理又显著高于EF2和EF1处理(P<0.05)。根中氮质量分数大小为EF2(13.95 g·kg-1)>EF3(13.86 g·kg-1)>CF(10.73 g·kg-1)>EF1(9.43 g·kg-1)>对照(7.69 g·kg-1)。与对照相比,常规施肥CF处理氮质量分数增加了39.6%,指数施肥EF处理增加了22.6%~81.4%,EF2和EF3处理显著高于CF和EF1处理(P<0.05)。茎中氮质量分数大小为EF3(13.77 g·kg-1)>EF2(10.14 g·kg-1)>EF1(6.94 g·kg-1)>CF(6.36 g·kg-1)>对照(5.45 g·kg-1)。与对照相比,常规施肥CF处理茎的氮质量分数减少了16.6%,差异不显著(P>0.05),指数施肥EF处理增加了27.3%~152.6%,达到显著性水平(P<0.05)。指数施肥EF处理显著高于常规施肥CF处理(P<0.05),增加了9.1%~116.5%。
表 2 不同施肥处理杉木幼苗不同器官氮质量分数和氮积累量
Table 2. Nitrogen concentration and accumulation of Chinese fir under different fertilization treatments
施肥处理 氮质量分数/(g·kg-1) 氮积累量/(mg·株-1) 根 茎 叶 根 茎 叶 对照 7.69 ± 0.26 c 6.44 ± 0.35 c 9.92 ± 0.51 d 77.98 ± 2.67 c 34.33 ± 1.88 c 76.22 ± 3.93 e 常规施肥CF 10.73 ± 0.27 b 5.94 ± 0.77 c 14.00 ± 0.27 c 148.57 ± 3.73 b 75.34 ± 9.72 b 279.84 ± 5.39 d 指数施肥EF1 9.43 ± 0.54 b 6.94 ± 0.35 bc 17.22±0.13 b 133.17 ± 7.73 b 84.33 ± 4.21 b 341.19 ± 2.67 c 指数施肥EF2 13.95 ± 0.62 a 10.14 ± 0.86 b 17.45 ± 0.48 b 186.19 ± 8.27 a 144.31 ± 12.23 a 443.49 ± 12.32 b 指数施肥EF3 13.86 ± 0.35 a 13.77 ± 0.14 a 21.31 ± 0.35 a 176.53 ± 4.46 a 156.49 ± 1.54 a 504.35 ± 7.47 a -
不同施肥处理下,杉木无性系幼苗叶的氮积累量大小为EF3(504.4 mg·株-1)>EF2(443.5 mg·株-1)>EF1(341.2 mg·株-1)>CF(279.8 mg·株-1)>对照(76.2 mg·株-1),5个处理之间差异均达到显著性水平(P<0.05)。与对照相比,施肥处理显著增加267.2%~561.7%(P<0.05);其中,指数施肥EF处理比常规施肥CF处理显著高出10.9%~66.2%(P<0.05)。根的氮积累量大小为EF2(186.2 mg·株-1)>EF3(176.5 mg·株-1)>CF(148.6 mg·株-1)>EF1(133.17 mg·株-1)>对照(80.0 mg·株-1)。与对照相比,施肥处理的根氮积累量显著增加了70.8%~138.8%(P<0.05),其中,指数施肥EF2和EF3处理显著高于常规施肥CF处理(P<0.05),分别高出25.3%,18.8%。茎的氮积累量大小为EF3(156.5 mg·株-1)>EF2(144.3 mg·株-1)>EF1(84.3 mg·株-1)>CF(75.3 mg·株-1)>对照(34.33 mg·株-1)。与对照相比,施肥处理增加了119.5%~355.8%,差异达显著性水平(P<0.05)。其中指数施肥EF处理与常规施肥CF处理相比,增加了11.9%~107.7%,EF2和EF3处理显著高于CF处理(P<0.05)。
-
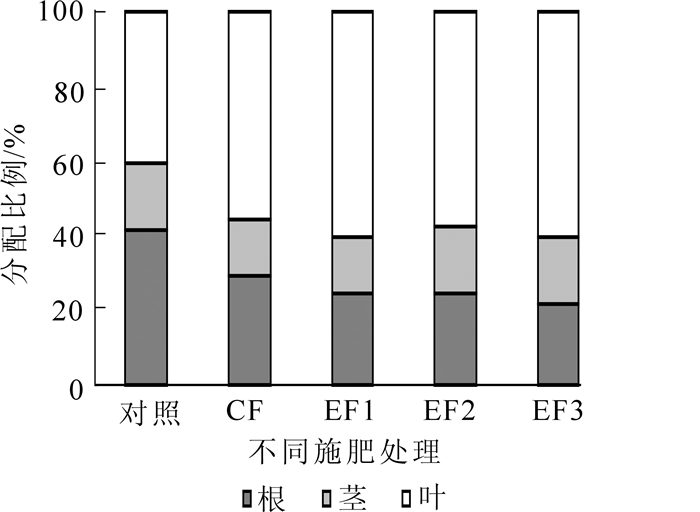
由图 3可知:不同施肥处理下,杉木无性系幼苗各器官的氮素分配率大小顺序总体表现为:叶>根>茎,其中叶氮积累量所占比例大小为EF3(60.2%)>EF1(61.1%)>EF2(57.3%)>CF(55.5%)>对照(40.4%),根中氮所占比例大小为对照(41.4%)>CF(29.5%)>EF2(24.1%)>EF1(23.8%)>EF3(21.1%),茎中氮积累量所占比例大小为EF3(18.7%)>EF2(18.6%)>对照(18.2%)>EF1(15.1%)>CF(15.0%),可以看出氮的主要储存器官是叶片,施氮可显著增加杉木叶片中氮积累量所占比例,降低根的氮积累量所占比例,使杉木幼苗的氮积累量由根向叶转移,而施氮对茎的氮积累量无明显差异。

图 3 不同施肥处理杉木不同器官氮积累的分配
Figure 3. Nitrogen accumulation and distribution of Chinese fir under different fertilization treatments
-
苗高和地径是衡量苗木品质的重要指标。生长结束时,施肥处理的苗高和地径显著高于不施肥处理。与对照相比,常规施肥苗高和地径分别增加34.0%和20.5%;指数施肥处理苗高和地径分别增加40.1%~60.5%和36.0%~40.4%。这与指数施肥显著提高沉香Aquilaria spp.株高和地径的研究结果相似[19]。其中指数施肥EF2处理苗高、地径都达到最佳生长状态,说明适量施氮能促进杉木无性系的生长,过量施氮抑制了杉木无性系的生长。这与王力朋等[13]的指数施肥对楸树Catalpa bungei无性系幼苗的影响研究结果相似。
生物量是衡量苗木生产力高低的重要指标。植物体内生物量分配受到施氮量的影响。不同施肥处理对杉木根生物量的影响并不显著,指数施肥中随着施氮量的增加根生物量略有下降,这是因为氮素缺乏时,植物会加大根系生物量的分配比例以提高根系的氮素吸收能力。指数施肥处理的茎、叶和全株生物量均随施氮总量的增加呈现出先增加后减小的趋势:施氮量为1.0 g·株-1时,生物量最大;施氮量增加至2.0 g·株-1时,生物量减少,说明适量施肥能促进无性系的生长[13],显著促进苗木干物质的积累[4, 12],而过量施氮使苗木受到轻微的毒害[7, 11],苗木的生长和生物量积累受到了抑制[13]。
施肥能提高苗木根、茎、叶的氮含量,而氮含量的提高能促进苗木生长和竞争力,从而提高造林成功的可能性。指数施肥显著提高了杉木无系性根、茎和叶的氮质量分数和氮积累量(P<0.05)。生长结束时,指数施肥处理茎、叶氮质量分数和氮积累量均显著高于常规施肥处理和对照(P<0.05),并随施氮量的增加而增加,这与MALIK等[4]和李双喜等[15]的研究结果相一致,而TIMMER[2]和SALIFU等[7]认为,在养分的添加过程中,当养分供不应求或充足时,苗木养分含量随养分供给的增加而增加;当养分供给超过苗木的最佳需求时,养分含量不会继续增加,甚至会下降。造成研究结果不同的原因可能与实验材料、试验方法不同有关[15]。
-
施肥显著提高了杉木幼苗生物量、氮质量分数及氮积累量。指数施肥中的EF2(施氮总量为1.0 g·株-1)处理杉木无性系表现出较高的生物量积累和养分苗木承载,说明指数施肥有效促进了杉木无性系苗木的生长,改善了苗木营养状态。从苗木生长性状、氮积累量考虑,1.0 g·株-1的施氮量是满足杉木无性系苗木生长的临界点,能获得较高的杉木生物学性状,是杉木无系性苗木温室培育的适宜施氮量。
Growth and N accumulation in seedlings of Cunninghamia lanceolata clones with N exponential fertilization
-
摘要: 杉木Cunninghamia lanceolata是中国特有的速生商品材树种。培育优质无性系苗木是杉木造林成功的关键。探讨了氮素指数施肥对杉木幼苗生长及养分积累的影响,为杉木苗木培育提供基础。以1年生无性系杉木幼苗为材料,采用温室盆栽方法,设定不施肥(对照)、常规施肥(CF)和指数施肥(EF1,EF2,EF3)共5个处理,纯氮施用总量分别为0,0.5,0.5,1.0,2.0 g·株-1,共施氮20次,间隔为10 d。至生长结束后,分别测定杉木的苗高、地径、生物量及氮素质量分数。结果表明:① 施肥显著促进了杉木无性系的苗高、地径和生物量的生长,以指数施肥EF2处理为最佳,分别为59.0 cm,6.0 mm,52.99 g·株-1;② 杉木无性系苗木不同器官的氮质量分数和氮积累量表现为叶>根>茎。与对照相比,常规施肥的根、茎、叶的氮质量分数增加39.6%,16.6%和41.1%,氮积累量增加90.5%,119.5%和267.2%;指数施肥的根、茎、叶的氮质量分数分别增加22.6%~81.4%,27.3%~152.6%和73.6%~135.5%,氮积累量分别增加70.8%~138.8%,145.7%~355.8%和347.6%~561.7%。除EF1处理外,杉木幼苗根、茎和叶的氮质量分数和氮积累量均表现为指数施肥显著大于常规施肥(P < 0.05)。施氮显著提高了杉木无性系幼苗的生长,其中施氮量为1 g·株-1的指数施肥是杉木幼苗温室培育的适宜方法。Abstract: High quality cloned seedlings of Chinese fir (Cunninghamia lanceolata), a fast-growing China-specific plant species dominant in south China, play an important role in Chinese fir forestation. To understand the effects of N exponential fertilization on seedling growth and nutrient content of Chinese fir, pot experiments were conducted in a greenhouse. One-year-old cloned seedlings were used with five N treatments per seedling including no fertilizer (ck) (0 g), conventional fertilization (CF) (0.5 g), and exponential fertilizations of EF1 (0.5 g), EF2 (1.0 g), and EF3 (2.0 g). Replications N fertilization was conducted 20 times at 10 d intervals. Height, ground diameter, biomass, and N absorption were measured after seedling growth. Analyses included state the sampling method and size. Results showed that fertilization of Chinese fir clones significantly promoted seedling height, ground diameter, and biomass (P < 0.05) with the EF2 treatment being optimal having the highest values of 59.0 cm for height, 6.0 mm for diameter (P < 0.05). N concentration and N content in different organs were leaves > roots > stems. Compared with ck, for conventional fertilization groups the N concentration increased in roots 39.6%, stems 16.6%, and leaves 41.1% with N content increasing 90.5% in roots, 119.5% in stems, and 267.2% in leaves; and for exponential fertilization groups the N concentration increased 22.6%-81.4% in roots, 27.3%-152.6% in stems, and 73.6%-135.5% in leaves with N content increasing 70.8%-138.8% in roots, 145.7%-355.8% in stems, and 347.6%-561.7% in leaves. Thus, N application rate had strong effect on growth of Chinese fir with the best method for cultivating Chinese fir cloned seedlings, considering seedling growth, N concentration, and N accumulation, being exponential N of 1 g·seedling-1.
-
图 2 不同施肥处理杉木幼苗苗高和地径
Figure 2 Plant height and ground diameter of Chinese firunder different fertilization treatments
图 3 不同施肥处理杉木不同器官氮积累的分配
Figure 3 Nitrogen accumulation and distribution of Chinese fir under different fertilization treatments
表 1 不同施肥处理杉木不同器官生物量
Table 1. Biomass of Chinese fir under different fertilization treatments
施肥处理 根/(g·株-1) 茎/(g·株-1) 叶/(g·株-1>) 全株/(g·株-1) 对照 10.14 ± 2.84 a 5.33 ± 1.58 b 7.68 ± 2.11 b 23.15 ± 6.40 b 常规施肥CF 13.84 ± 3.33 a 12.68 ± 1.92 a 19.99 ± 4.79 a 46.51 ± 9.35 a 指数施肥EF1 14.13 " 2.24 a 12.16 ± 0.47 a 19.81 ± 1.86 a 46.10 ± 4.08 a 指数施肥EF2 13.35 ± 2.29 a 14.23 ± 3.61 a 25.41 ± 2.99 a 52.99 ± 8.85 a 指数施肥EF3 12.74 ± 1.38 a 11.40 ± 3.16 a 21.32 ± 2.82 a 45.45 ± 6.97 a  下载: 导出CSV
下载: 导出CSV
表 2 不同施肥处理杉木幼苗不同器官氮质量分数和氮积累量
Table 2. Nitrogen concentration and accumulation of Chinese fir under different fertilization treatments
施肥处理 氮质量分数/(g·kg-1) 氮积累量/(mg·株-1) 根 茎 叶 根 茎 叶 对照 7.69 ± 0.26 c 6.44 ± 0.35 c 9.92 ± 0.51 d 77.98 ± 2.67 c 34.33 ± 1.88 c 76.22 ± 3.93 e 常规施肥CF 10.73 ± 0.27 b 5.94 ± 0.77 c 14.00 ± 0.27 c 148.57 ± 3.73 b 75.34 ± 9.72 b 279.84 ± 5.39 d 指数施肥EF1 9.43 ± 0.54 b 6.94 ± 0.35 bc 17.22±0.13 b 133.17 ± 7.73 b 84.33 ± 4.21 b 341.19 ± 2.67 c 指数施肥EF2 13.95 ± 0.62 a 10.14 ± 0.86 b 17.45 ± 0.48 b 186.19 ± 8.27 a 144.31 ± 12.23 a 443.49 ± 12.32 b 指数施肥EF3 13.86 ± 0.35 a 13.77 ± 0.14 a 21.31 ± 0.35 a 176.53 ± 4.46 a 156.49 ± 1.54 a 504.35 ± 7.47 a
下载: 导出CSV
-
[1] 孙慧彦, 刘勇, 马履一, 等.氮磷供给对长白落叶松苗木质量的影响[J].内蒙古农业大学学报, 2011, 32(3):58-62. SUN Huiyan, LIU Yong, MA Lüyi, et al. Effects of supplying of nitrogen phosphorus to Larix olgensis seedling quality[J]. J Inn Mongolia Agric Univ, 2011, 32(3):58-62. [2] TIMMER V R. Exponential nutrient loading:a new fertilization technique to improve seedlings performance on competitive sites[J]. New For, 1996, 13(1):279-299. [3] OLIET J A, PLANELLES R, APTERO F, et al. Field performance of Pinus halepensis planted in Mediterranean arid condition:relative influence of seeding morphology and mineral nutrition[J]. New For, 2009, 37(3):313-331. [4] MALIK V, TIMMER V R. Biomass partitioning and nitrogen retranslocation in black spruce seedlings on competitive mixed wood sites:a bioassay study[J]. Can J For Res, 1998, 28(2):206-215. [5] SALIFU K F, JACOBS D F, BIRGE Z K D. Nursery nitrogen loading improves field performance of bare root oak seedlings planted on abandoned mine lands[J]. Restor Ecol, 2009, 17(3):339-349. [6] 魏红旭, 徐程扬, 马履一, 等.苗木指数施肥技术研究进展[J].林业科学, 2010, 46(7):140-146. WEI Hongxu, XU Chengyang, MA Lüyi, et al. Advances in study on seedling exponential fertilization regime[J]. Sci Silv Sin, 2010, 46(7):140-146. [7] SALIFU K F, TIMMER V R. Nitrogen retranslocation response of young Picea mariana to nitrogen-15 supply[J]. Soil Sci Soc Am J, 2003, 67:309-317. [8] QU Laiye, QUORESHI A M, KOIKE T. Root growth characteristics, biomass and nutrient dynamics of seedlings of two larch species raised under different fertilization regimes[J]. Plant Soil, 2003, 255(1):293-302. [9] HAWKINS B J, BURGESS D, MITCHELLl A K. Growth and nutrient dynamics of western hemlock with conventional or exponential greenhouse fertilization and planting in different fertility conditions[J]. Can J For Res, 2005, 35(4):1002-1016. [10] EVERETT K T, HAWKINS B J, KIISKILA S. Growth and nutrient dynamics of Douglas-fir seedlings raised with exponential or conventional fertilization and planted with or without fertilizer[J]. Can J For Res, 2007, 37(12):2552-2562. [11] 李国雷, 祝燕, 蒋乐, 等.指数施肥对栓皮栎容器苗生长和氮积累的影响[J].东北林业大学学报, 2012, 40(11):6-9. LI Guolei, ZHU Yan, JIANG Le, et al. Effect of exponential fertilization on growth and nitrogen storage of containerized Quercus variabilis seedlings[J]. J Northeast For Univ, 2012, 40(11):6-9. [12] 郝龙飞, 王庆成, 张彦东, 等.指数施肥对山桃稠李播种苗生物量及养分动态的影响[J].林业科学, 2012, 48(6):33-39. HAO Longfei, WANG Qingcheng, ZHANG Yandong, et al. Effect of exponential fertilization on biomass and nutrient dynamics of Padus maackii seedlings[J]. Sci Silv Sin, 2012, 48(6):33-39. [13] 王力朋, 晏紫伊, 李吉跃, 等.指数施肥对楸树无性系生物量分配和根系形态的影响[J].生态学报, 2012, 32(23):7452-7462. WANG Lipeng, YAN Ziyi, LI Jiyue, et al. Effects of exponential fertilization on biomass allocation and root morphology of Catalpa bungei clones[J]. Acta Ecol Sin, 2012, 32(23):7452-7462. [14] 李双喜, 杨曾奖, 徐大平, 等.施氮量对檀香幼苗生长及养分积累的影响[J].植物营养与肥料学报, 2015, 21(3):807-814. LI Shuangxi, YANG Zengjiang, XU Daping, et al. Effects of nitrogen application rate on growth and nutrient accumulation of Santalum album L. seedlings[J]. J Plant Nutr Fert, 2015, 21(3):807-814. [15] 徐嘉科, 陈闻, 王晶, 等.不同施肥方式对红楠生长及营养特性的影响[J].生态学杂志, 2015, 34(5):1241-1245. XU Jiake, CHEN Wen, WANG Jing, et al. Effects of different fertilization strategies on growth and nutrition characteristics of Machilus thunbergii[J]. Chin J Ecol, 2015, 34(5):1241-1245. [16] 陈清堤.杉木无性系造林对比试验及重复力估测[J].林业调查规划, 2012, 35(6):140-144. CHEN Qingti. Comparison test and repeatability estimates on Cunninghamia lanceolata clones afforestation[J]. For Invent Plan, 2012, 35(6):140-144. [17] 齐明.我国杉木无性系选育的成就、问题和对策[J].世界林业研究, 2007, 20(6):50-54. QI Ming. Achievements, problem sand counter measures of clonal selection and breeding of Chinese fir in China[J]. World For Res, 2007, 20(6):50-54. [18] XU Xinjian, TIMMER V R. Biomass and nutrient dynamics of Chinese fir seedlings under conventional and exponential fertilization regimes[J]. Plant Soil, 1998, 203(2):313-322. [19] 王冉, 李吉跃, 张方秋, 等.不同施肥方法对马来沉香和土沉香苗期根系生长的影响[J].生态学报, 2011, 31(1):98-106. WANG Ran, LI Jiyue, ZHANG Fangqiu, et al. Growing dynamic root system of Aquilaria malaccensis and Aquilaria sinensis seedlings in response to different fertilizing methods[J]. Acta Ecol Sin, 2011, 31(1):98-106. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2017.03.011
 点击查看大图
点击查看大图
计量
- 文章访问数: 3730
- HTML全文浏览量: 727
- PDF下载量: 435
- 被引次数: 0
