




-
水稻Oryza sativa是世界上最重要的粮食作物之一,全球超过一半的人以稻米为粮食[1]。近年来,随着中国水稻种植面积的提升,以及水稻的逐年丰收,水稻贮藏问题越来越突出。在世界范围内,每年因贮藏期间虫害和霉变等问题造成巨大的粮食损失,损失量超过总量的5%[2],严重影响粮食安全和食品安全。同时,贮藏过程中引起水稻种子发芽率下降,对水稻生产和育种过程也会产生不良影响。目前,水稻种子通常采用低温冷库贮藏,通过控制温度和湿度来改善其贮藏环境[3−4]。但这种方法成本高,也会造成环境污染。因此,筛选耐贮的水稻种质用于育种,是目前解决水稻贮藏问题最经济有效的途径之一,对保障国家粮食安全和用种安全,以及为企业和农户提供高发芽率的优质种质具有重大的意义。
水稻的耐贮性主要包括两方面内容:作为稻米消费,长时间贮藏后仍能保持原有风味;作为种子或种质保存,长时间贮藏后仍具有较强的活力[5]。目前学者对于水稻耐贮特性的研究集中在后者[6−10]。研究表明:水稻的耐贮性是一个受到遗传、生长发育、成熟以及收获后的环境等许多因素影响的一个复杂性状。目前,水稻种子耐贮性的评价指标通常采用发芽率、发芽势、发芽指数、活力指数和半衰期表示。由于发芽率、发芽势的测定较为简单,同时也更贴近生产实际,因此一般将发芽率、发芽势作为评价水稻耐贮性的指标。水稻种子耐贮性的研究方法主要有自然老化法和人工老化法[11−12]。自然老化法是指种子在自然贮藏或种质保存的条件下,发芽力逐渐丧失的过程;人工老化是指采用高温高湿的环境来加速种子老化速度研究其耐贮藏性的方法[13]。王仪春等[14]研究发现:人工加速老化后种子劣变程度增加,表现为丙二醛(MDA)含量显著上升,抗氧化物酶活性均下降。张瑛等[15]连续6 a测定入库的水稻种子电导率,结果显示随着贮藏时间的延长,种子电导率先急速增长,再缓慢增长,之后迅速下降,说明电解质渗漏导致电导率大幅增加,膜透性增强。
由于自然老化最符合种子贮藏的自然特征,因此本研究采用自然老化法对不同品种的水稻种子耐贮性进行筛选鉴定,通过对耐贮品种与不耐贮品种的淀粉酶活性、抗氧化酶活性、MDA质量摩尔浓度以及脯氨酸(Pro)质量分数等生理指标之间的差异进行分析,为水稻耐贮种质筛选评价及耐贮特性研究提供理论依据。
-
以220份水稻品种为材料,其中常规粳稻34份,常规籼稻11份,杂交稻175份。供试水稻种子在2015年由浙江省种子管理总站提供,所有品种均于黄熟期时收获。按照常规方法贮藏于浙江农林大学冷库中,贮藏温度为10~15 ℃,湿度为45%~65%。
采用自然老化处理,2015年测定其原始发芽率、原始发芽势(未经自然老化);分别于2020、2021年再次测定其发芽率、发芽势,并对筛选的耐贮和不耐贮品种的生理指标进行测定。
-
参照ISTA标准[16],每个品种随机数取100粒种子,3次重复,用质量浓度为1.0%次氯酸钠消毒10 min后冲洗干净。将种子均匀放置在湿润的无菌发芽纸上,然后将种子置于30 ℃光照8 h (光照强度为12 000 lx),20 ℃黑暗16 h的培养箱中培养14 d,每天记载发芽数并适当补充水分。计算公式为:发芽率=14 d内正常发芽幼苗数/供试种子数×100%;发芽势=5 d内正常发芽幼苗数/供试种子数×100%。将发芽率下降幅度<5%且发芽势下降幅度<15%的品种作为耐贮品种,将发芽率下降幅度≥5%且发芽势下降幅度≥15%的品种作为不耐贮品种。
-
采用3,5-二硝基水杨酸比色法测定淀粉酶活性[17],过氧化氢酶(CAT)活性采用紫外吸光法测定[18],过氧化物酶(POD)活性采用愈创木酚法测定[19],超氧化物歧化酶(SOD)活性采用氮蓝四唑法测定[20],丙二醛采用硫代巴比妥酸显色法测定[21],脯氨酸采用磺基水杨酸提取酸性茚三酮显色法测定[22]。
-
使用Excel 2010和GraphPad Prism 8.0进行数据的整理与作图,使用SPSS 19.0软件进行单因素方差(one-way ANOVA)分析(Duncan test)。
-
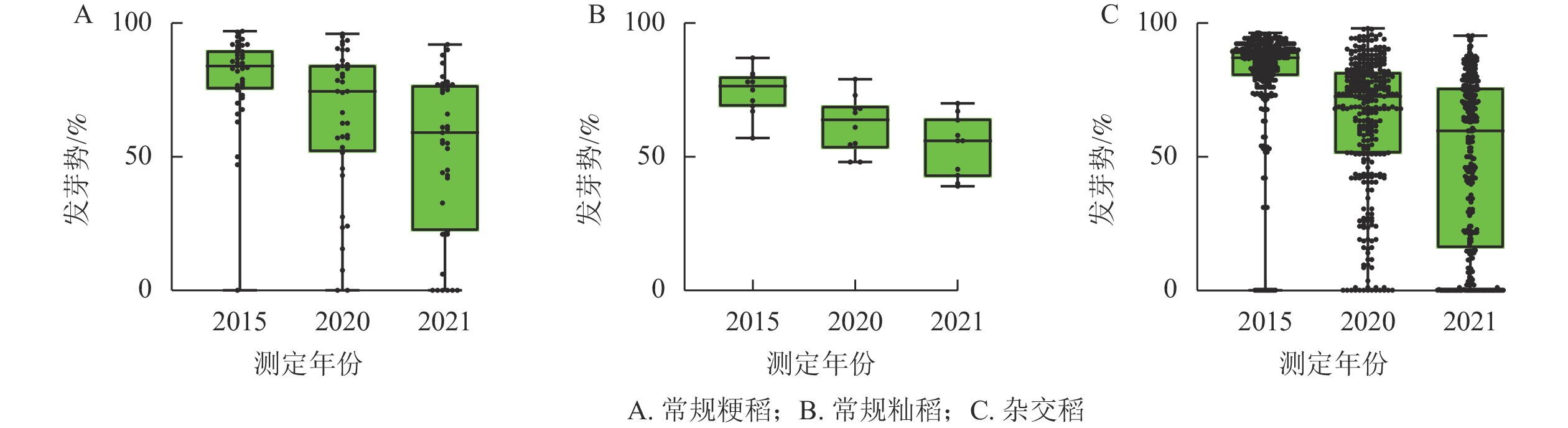
随着贮藏时间的延长,不同品种的种子活力相关参数均呈现下降趋势(图1和图2)。常规粳稻的平均原始发芽率为87.6%,常规籼稻的平均原始发芽率为86.4%,杂交稻的平均原始发芽率为88.9%;贮藏5 a之后,常规粳稻、常规籼稻、杂交稻的平均发芽率分别下降了14.1%、10.2%、16.4%;平均发芽势分别下降了24.2%、19.6%、25.4%。贮藏6 a之后,常规粳稻、常规籼稻、杂交稻的平均发芽率分别下降了62.2%、23.4%、45.3%;平均发芽势分别降低了55.0%、38.1%、50.7%。不同类型水稻间种子活力的变化趋势保持一致。
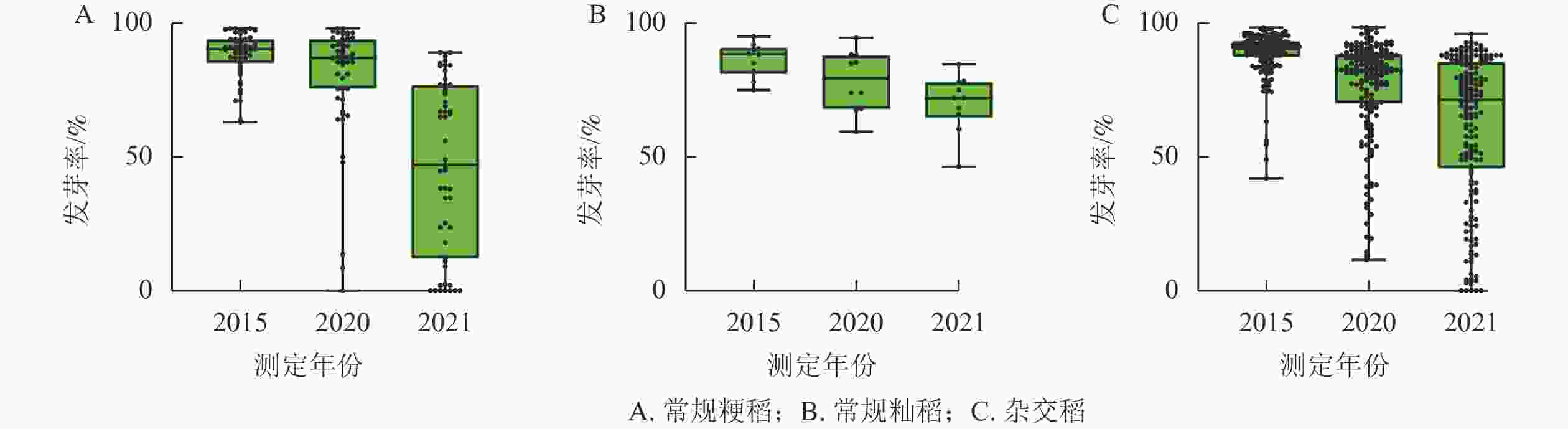
图 1 不同类型水稻种子在不同贮藏时期的发芽率变化
Figure 1. Changes of germination rate of different types of rice seeds in different storage periods

图 2 不同类型水稻种子在不同贮藏时期的发芽势变化
Figure 2. Changes of germination potential of different types of rice seeds in different storage periods
-
由表1可见:随着贮藏时间的延长,常规粳稻的发芽率不断下降。贮藏5 a,‘南粳44’‘Nanjing 44’、‘沈稻18’‘Shendao 18’、‘天稻丰’‘Tiandaofeng’、‘常农粳7号’‘Changnongjing 7’、‘秀水134’‘Xiushui 134’和‘平安粳稻11号’‘Pinganjingdao 11’的发芽率分别下降了2.4%、3.2%、3.3%、27.1%、23.9%、17.7%,其中发芽率‘沈稻18’的下降幅度最小,‘常农粳7号’的下降幅度最大,达显著差异(P<0.05)。贮藏6 a,发芽率的下降趋势与贮藏5 a一致。随着贮藏时间的延长,常规粳稻的发芽势不断下降。贮藏5 a,‘南粳44’‘沈稻18’‘天稻丰’‘平安粳稻11号’‘常农粳7号’‘秀水134’的发芽势分别下降了4.2%、10.7%、13.6%、20.5%、34.7%、54.0%。其中‘南粳44’发芽势的下降幅度最小,‘秀水134’的下降幅度最大,达显著差异(P<0.05)。贮藏6 a时的发芽势与5 a的下降趋势一致。在常规粳稻类型中,筛选出‘沈稻18’‘南粳44’‘天稻丰’等为耐贮品种,‘常农粳7号’‘秀水134’‘平安粳稻11号’等为不耐贮品种。
表 1 不同常规粳稻品种在不同贮藏时期发芽率和发芽势的变化
Table 1. Variation of germination rate and germination potential of different conventional japonica rice cultivars in different storage periods
类型 品种名 2015年 2020年 2021年 发芽率/% 发芽势/% 发芽率/% 发芽势/% 发芽率/% 发芽势/% 耐贮品种 ‘南粳44’ 96.3±0.6 a 91.7±0.6 a 93.3±0.6 a 88.0±2.0 b 84.3±4.2 b 66.0±1.0 c ‘沈稻18’ 97.0±1.0 a 90.0±2.0 a 94.7±1.2 a 81.3±1.5 b 82.3±5.5 b 59.0±2.0 c ‘天稻丰’ 94.7±1.5 a 86.7±1.5 a 91.7±2.3 a 76.3±2.5 b 73.7±5.8 b 59.3±6.4 c 不耐贮品种 ‘常农粳7号’ 97.0±1.0 a 89.3±0.6 a 76.3±2.1 b 66.3±1.5 b 38.0±5.3 c 33.3±2.1 c ‘秀水134’ 97.0±1.0 a 91.3±1.2 a 78.3±2.5 b 59.3±1.5 b 37.3±0.6 c 32.7±1.5 c ‘平安粳稻11号’ 95.7±0.6 a 88.0±1.0 a 81.3±2.5 b 73.0±3.6 b 25.7±1.2 c 22.7±2.1 c 说明:数据为平均值±标准差。不同小写字母表示同一品种在不同贮藏时期间差异显著(P<0.05) -
由表2可见:随着贮藏时间的延长,常规籼稻的发芽率不断下降。贮藏5 a,‘赣晚籼35号’‘Ganwanxian 35’、‘甬籼69’‘Yongxian 69’、‘赣晚糯7号’‘Ganwannuo 7’、‘温926’‘Wen 926’、‘扬稻6号’‘Yangdao 6’和‘甬籼15号’‘Yongxian 15’的发芽率分别下降4.3%、4.1%、3.2%、17.1%、17.4%、22.0%;其中,‘赣晚糯7号’的下降幅度最小,‘甬籼15号’的下降幅度最大,达显著差异(P<0.05)。贮藏6 a,发芽率的下降与5 a趋势一致。随着贮藏时间的延长,常规籼稻的发芽势不断下降。贮藏5 a‘甬籼69’‘ 赣晚糯7号’‘赣晚籼35号’‘温926’‘扬稻6号’‘甬籼15号’的发芽势分别下降2.9%、7.4%、8.8%、23.2%、30.5%、50.6%;其中‘甬籼69’的下降幅度最小,‘甬籼15号’的下降幅度最大,达显著差异(P<0.05)。贮藏6 a,发芽势的下降与5 a趋势一致。常规籼稻筛选出‘赣晚籼35号’‘赣晚糯7号’‘甬籼69’等为耐贮品种,‘温926’‘甬籼15号’‘扬稻6号’等为不耐贮品种。
表 2 不同常规籼稻品种在不同贮藏时期发芽率和发芽势变化
Table 2. Variation of germination rate and germination potential of different conventional indica rice cultivars in different storage periods
类型 品种名 2015年 2020年 2021年 发芽率/% 发芽势/% 发芽率/% 发芽势/% 发芽率/% 发芽势/% 耐贮品种 ‘赣晚籼35号’ 97.3±2.1 a 90.0±1.0 a 93.3±3.1 a 82.7±1.2 b 84.7±2.5 b 81.3±2.5 b ‘赣晚糯7号’ 96.0±1.0 a 91.3±1.2 a 93.0±1.7 a 85.0±3.6 a 78.3±5.9 b 70.0±4.0 b ‘甬籼69’ 95.0±1.0 a 83.0±2.6 a 91.3±3.2 a 80.7±3.1 a 72.0±7.5 b 63.3±2.1 b 不耐贮品种 ‘温926’ 91.7±1.5 a 88.7±2.1 a 78.3±0.6 b 72.0±2.0 b 52.3±5.1 c 41.3±1.5 c ‘甬籼15号’ 88.7±1.2 a 83.3±0.6 a 72.7±5.0 b 55.3±1.2 b 46.3±4.0 c 37.0±1.0 c ‘扬稻6号’ 87.7±2.3 a 81.3±1.5 a 74.7±2.5 b 62.3±2.5 b 29.7±7.1 c 26.0±1.0 c 说明:数据为平均值±标准差。不同小写字母表示同一品种在不同贮藏时期间差异显著(P<0.05) -
由表3所示:随着贮藏时间的延长,杂交稻的发芽率不断下降。贮藏5 a,‘两优培九’‘Liangyou Peijiu’、‘中浙优1号’‘Zhongzhenyou 1’、‘晶两优华占’‘Jingliangyou Huazhan’、‘深两优865’‘Shenliangyou 865’、‘两优1号’‘Liangyou 1’和‘隆香优130’‘Longxiangyou 130’的发芽率分别下降1.5%、2.1%、2.9%、21.5%、25.3%、35.6%;其中‘两优培九’的下降幅度最小,‘隆香优130’的下降幅度最大,达显著差异(P<0.05)。贮藏6 a,发芽率的下降趋势与5 a保持一致。随着贮藏时间的延长,杂交稻的发芽势不断下降。贮藏5 a,‘中浙优1号’‘晶两优华占’‘两优培九’‘深两优865’‘两优1号’‘隆香优130’的发芽势分别下降3.4%、3.5%、6.0%、25.2%、37.7%、46.3%;其中,‘中浙优1号’的下降幅度最小,‘隆香优130’的下降幅度最大,达显著差异(P<0.05)。贮藏6 a,发芽势的下降与5 a趋势一致。杂交稻品种筛选出‘两优培九’‘中浙优1号’‘晶两优华占’等为耐贮品种,‘隆香优130’‘深两优865’‘两优1号’等为不耐贮品种。
表 3 不同杂交稻品种在不同贮藏时期发芽率变化
Table 3. Changes of germination rate of different hybrid rice cultivars in different storage periods
类型 品种名 2015年 2020年 2021年 发芽率/% 发芽势/% 发芽率/% 发芽势/% 发芽率/% 发芽势/% 耐贮品种 ‘两优培九’ 97.7±1.5 a 89.0±3.0 a 96.3±1.5 a 84.0±2.0 a 92.7±2.1 a 78.0±1.0 b ‘中浙优1号’ 96.3±0.6 a 90.3±1.5 a 94.3±8.6 a 87.3±2.3 a 91.3±3.2 a 82.3±1.5 b ‘晶两优华占’ 96.0±1.0 a 89.0±1.0 a 93.3±0.6 a 86.0±1.0 a 92.3±0.6 a 77.0±1.0 b 不耐贮品种 ‘隆香优130’ 96.3±1.5 a 90.7±1.5 a 71.0±1.7 b 62.0±2.0 b 38.7±2.1 c 28.3±2.5 c ‘深两优865’ 96.0±2.0 a 86.0±1.7 a 79.0±2.0 b 68.7±3.1 b 24.7±3.1 c 18.3±1.5 c ‘两优1号’ 94.0±1.0 a 91.3±2.1 a 75.0±1.0 b 66.3±2.1 b 29.0±1.7 c 20.3±2.1 c 说明:数据为平均值±标准差。不同小写字母表示同一品种在不同贮藏时期间差异显著(P<0.05) -
常规粳稻不同耐贮品种间种子淀粉酶活性、抗氧化酶活性、MDA质量摩尔浓度和Pro质量分数存在明显差异(表4)。耐贮品种的α-淀粉酶、β-淀粉酶、CAT、POD、SOD活性和Pro质量分数的平均值比不耐贮品种分别高650%、74.5%、20.7%、23.3%、18.9%、25.0%;耐贮品种的MDA质量摩尔浓度平均值比不耐贮品种低了48.6%。其中,α-淀粉酶活性最强的依次为耐贮品种‘沈稻18’‘南粳44’‘秀水134’,均显著高于不耐贮品种(P<0.05);‘沈稻18’的α-淀粉酶活性比不耐贮品种‘天稻丰’显著高200.5% (P<0.05)。β-淀粉酶活性最强的依次为耐贮品种‘秀水134’‘沈稻18’‘南粳44’,其中‘秀水134’‘沈稻18’均显著高于不耐贮品种(P<0.05),‘秀水134’的β-淀粉酶活性比不耐贮品种‘常农粳7号’显著高167.4% (P<0.05)。抗氧化酶活性最强、Pro质量分数最高的为耐贮品种‘南粳44’,均显著高于不耐贮品种(P<0.05)。MDA质量摩尔浓度最低的为耐贮品种‘南粳44’,最高的为不耐贮品种‘天稻丰’。
表 4 不同品种常规粳稻生理指标变化
Table 4. Changes of physiological indexes of different cultivars of conventional japonica rice
类型 品种 α-淀粉酶活性/
(mg·g−1·min−1)β-淀粉酶活性/
(mg·g−1·min−1)CAT活性/
(×16.67 nkat·g−1)POD活性/
(×16.67 nkat·g−1)SOD活性/
(×16.67 nkat·g−1)MDA/
(μmol·kg−1)Pro/
(μg·g−1)耐贮品种 ‘南粳44’ 2.9±0.5 b 5.8±0.6 c 85.7±1.4 a 305.1±1.9 a 212.2±9.7 a 2.7±0.7 d 14.7±2.1 a ‘沈稻18’ 4.3±0.3 a 8.6±0.9 b 63.7±3.3 b 224.3±10.3 b 211.8±1.2 a 4.7±0.2 b 12.3±1.0 b ‘秀水134’ 1.8±0.1 c 12.3±1.8 a 62.4±1.8 b 133.7±3.6 d 118.9±3.1 d 4.0±0.1 bc 10.6±0.5 bc 平均值 3.0±1.3 A 8.9±3.3 A 70.6±13.1 A 221.0±85.7 A 181.0±53.8 A 3.8±1.0 A 12.5±2.1 A 不耐贮品种 ‘平安粳稻11号’ 0.4±0.2 d 5.5±0.3 c 62.9±1.3 b 181.5±2.1 c 122.7±3.5 d 3.3±0.1 cd 9.8±0.5 c ‘常农粳7号’ 0.6±0.1 d 4.6±0.3 c 64.2±0.5 b 182.1±1.7 c 159.6±8.7 c 9.2±0.9 a 9.9±0.4 c ‘天稻丰’ 0.2±0.2 d 5.3±0.1 c 48.4±1.2 c 174.1±3.1 c 174.3±2.9 b 9.7±0.8 a 10.5±1.4 bc 平均值 0.4±0.2 A 5.1±0.5 A 58.5±8.8 A 179.2±4.4 A 152.2±26.6 A 7.4±3.6 A 10.0±0.4 A 说明:数据为平均值±标准差。同列不同小写字母表示各品种间差异显著,同列不同大写字母表示不同耐贮性的平均值差异显著(P<0.05) -
常规籼稻不同耐贮品种间种子淀粉酶活性、抗氧化酶活性、MDA质量摩尔浓度和Pro质量分数存在明显差异(表5)。耐贮品种的α-淀粉酶、β-淀粉酶、CAT、POD、SOD活性和Pro质量分数的平均值比不耐贮品种分别高200%、137.5%、82.4%、80.5%、9.7%、11.4%;耐贮品种MDA质量摩尔浓度的平均值比不耐贮品种低了52.8%。其中,α-淀粉酶活性、POD活性达到显著差异(P<0.05)。耐贮品种‘赣晚籼35号’‘赣晚糯7号’‘甬籼69’的α-淀粉酶活性显著高于不耐贮品种‘农乐1号’‘甬籼15号’和 ‘扬稻6号’。β-淀粉酶活性和CAT活性最高的是耐贮品种‘甬籼69’,其中,β-淀粉酶活性分别比不耐贮品种高156.1%、252.1%、213.0%,差异显著(P<0.05)。POD、SOD活性最强的是耐贮品种‘赣晚糯7号’,显著高于不耐贮品种(P<0.05)。Pro质量分数最高的为耐贮品种‘甬籼69’,显著高于其他5个品种(P<0.05),但其他5个品种之间差异不显著。
表 5 不同品种常规籼稻生理指标变化
Table 5. Changes of physiological indexes of different cultivars of conventional indica rice
类型 品种 α-淀粉酶活性/
(mg·g−1·min−1)β-淀粉酶活性/
(mg·g−1·min−1)CAT活性/
(×16.67 nkat·g−1)POD活性/
(×16.67 nkat·g−1)SOD活性/
(×16.67 nkat·g−1)MDA/
(μmol·kg−1)Pro/
(μg·g−1)耐贮品种 ‘赣晚籼35号’ 4.5±0.5 a 11.2±1 b 54.9±2.6 c 328.1±11.6 b 210.8±6.7 b 2.3±0.2 e 10.1±0.6 b ‘赣晚糯7号’ 3.7±0.3 b 11.7±1 b 77.1±4.4 b 468.4±10.1 a 224.8±0.3 a 2.4±0.3 e 10.8±1.3 b ‘甬籼69’ 4.3±0.3 a 16.9±1.6 a 104.5±3.0 a 271.8±8.3 c 212.9±5.7 b 5.6±0.2 d 14.1±1.0 a 平均值 4.2±0.4 A 13.3±3.2 A 78.8±24.8 A 356.1±101.2 A 216.2±7.6 A 3.4±1.9 A 11.7±2.1 A 不耐贮品种 ‘农乐1号’ 1.2±0.2 d 6.6±0.4 c 46.3±1.7 d 127.7±10.8 e 211.7±4.8 b 7.2±0.3 b 10.3±1.2 b ‘甬籼15号’ 0.6±0.1 e 4.8±0.4 c 28.2±2.4 e 279.8±4.9 c 176.8±3.9 d 7.8±0.2 a 11.1±0.4 b ‘扬稻6号’ 2.3±0.2 c 5.4±0.5 c 55.0±1.2 c 184.4±1.9 d 202.9±1.7 c 6.7±0.2 c 10.2±0.2 b 平均值 1.4±0.9 B 5.6±0.9 A 43.2±13.7 A 197.3±76.9 B 197.1±18.2 A 7.2±0.6 A 10.5±0.5 A 说明:数据为平均值±标准差。同列不同小写字母表示各品种间差异显著,同列不同大写字母表示不同耐贮性的平均值差异显著(P<0.05) -
杂交稻不同耐贮品种间种子淀粉酶活性、抗氧化酶活性、MDA质量摩尔浓度和Pro质量分数存在明显差异(表6)。耐贮品种的α-淀粉酶、β-淀粉酶、CAT、POD、SOD活性和Pro质量分数的平均值比不耐贮品种分别高357.1%、186.0%、142.1%、161.9%、50.0%、83.3%;耐贮品种MDA质量摩尔浓度的平均值比不耐贮品种低了82.7%。其中,α-淀粉酶活性、CAT活性、SOD活性均达到显著差异(P<0.05)。其中,α-淀粉酶、β-淀粉酶活性最高的为耐贮品种‘两优培九’,显著高于不耐贮品种‘隆香优130’‘深两优865’‘两优1号’(P<0.05)。CAT活性最强、Pro质量分数最高的为耐贮品种‘中浙优1号’,均显著高于不耐贮品种(P<0.05);POD活性最强的为‘晶两优华占’,分别显著高出不耐贮品种‘隆香优130’‘深两优865’‘两优1号’308.6%、182.5%、113.7%。SOD活性最强,同时MDA质量摩尔浓度最低的为耐贮品种‘两优培九’,MDA质量摩尔浓度最高的为不耐贮品种‘隆香优130’。
表 6 不同品种杂交稻生理指标变化
Table 6. Changes of physiological indexes of different cultivars of hybrid rice
类型 品种 α-淀粉酶活性/
(mg·g−1·min−1)β-淀粉酶活性/
(mg·g−1·min−1)CAT活性/
(×16.67 nkat·g−1)POD活性/
(×16.67 nkat·g−1)SOD活性/
(×16.67 nkat·g−1)MDA/
(μmol·kg−1)Pro/
(μg·g−1)耐贮品种 ‘两优培九’ 8.5±0.2 a 22.4±0.4 a 133.5±5.7 b 201.4±3.7 c 229.9±7.6 a 2.0±0.6 e 12.0±0.4 c ‘中浙优1号’ 4.0±0.7 c 12.9±1.1 b 169.4±3.0 a 376.4±9.5 b 229.6±3.1 a 2.7±0.2 d 31.4±1.3 a ‘晶两优华占’ 6.6±0.2 b 13.5±0.1 b 119.9±1.3 c 430.3±11.3 a 206.9±1.6 b 2.2±0.5 e 16.1±1.8 b 平均值 6.4±2.3 A 16.3±5.3 A 140.9±25.6 A 336.0±119.7 A 222.1±13.2 A 2.3±0.4 A 19.8±10.2 A 不耐贮品种 ‘隆香优130’ 2.7±0.4 d 3.6±0.1 e 60.6±3.3 d 105.3±4.6 f 144.8±3.1 d 14.5±0.5 a 12.6±1.2 c ‘深两优865’ 0.6±0.1 e 8.7±0.2 c 53.8±2.2 d 152.3±4.8 d 175.8±3.7 c 7.5±0.3 c 10.6±0.7 cd ‘两优1号’ 0.9±0.1 e 4.7±0.4 d 60.2±3.4 d 127.5±7.2 e 123.8±2.5 e 12.2±0.4 b 9.4±0.7 d 平均值 1.4±1.1 B 5.7±2.7 A 58.2±3.8 B 128.3±23.5 A 148.1±26.2 B 11.4±3.6 A 10.8±1.6 A 说明:数据为平均值±标准差。同列不同小写字母表示各品种间差异显著,同列不同大写字母表示不同耐贮性的平均值差异显著(P<0.05) -
种子是遗传物质的载体和农业生产最基本的资料,是一个活的生命有机体。在水稻种子贮藏过程中会遇到各种逆境胁迫,胁迫因子会对种子的品质造成很大影响,进而对作物生产造成不可逆的影响[23]。同时种子内部在贮藏过程中的一系列代谢活动会引起贮藏养分的衰竭、化学组成的改变、细胞膜结构的破坏以及酶活性的丧失等变化。所有这些变化最终导致种子遗传完整性丧失,表现性状为种子活力的下降或丧失,对农业生产、储运和经营带来重大损失。因此从水稻种质资源中选取耐贮藏的优异品种进行育种和培育,是解决水稻种子贮藏问题的根本途径。
从本研究可以看出:不同水稻品种间耐贮性存在巨大的差异。常规粳稻、常规籼稻、杂交稻在贮藏5和6 a时,发芽率分别平均下降了12.8%、16.2%、14.8%;发芽势分别平均下降了23.0%、20.6%、20.4%。RAO等[24]研究认为:在耐贮性上籼稻>爪哇稻>粳稻。曾大力等[25]研究表明:籼粳指数与耐贮特性呈显著的正相关,籼稻比粳稻更耐贮藏。但不管是粳稻或籼稻,都存在耐贮性好或差的极端情况。本研究中,常规粳稻、常规籼稻、杂交稻的种子活力下降趋势基本一致,但下降幅度不同。水稻种子耐贮性与种子的寿命和种子活力紧密相关。种子活力受到种子发育、成熟期间或生长环境条件等多因素影响,尤其是温度、水分、光照、土壤和气候条件等。因此不同栽培条件以及不同田间管理的水稻品种其耐贮性有着很大不同[25],收获过早或过晚的种子其活力都大大降低[26−27]。侯文平等[28]研究表明:收获过早,种胚未完全成熟,干物质未完全充实籽粒,会造成种子产量降低;收获过晚,则导致种子自然老化,种子发芽率下降。因此适时收获是保持种子高活力,提高耐贮性的有效措施。
大量文献报道:常规稻种子耐贮性较杂交稻种子好[29−30],并认为杂交稻种子易萌发的特点是导致种子耐贮性差的根本原因[31]。但在本研究中,杂交稻的发芽率、发芽势和生理指标优于常规粳稻、常规籼稻。但耐贮性较强的品种也具有较强的休眠特性,在育种过程中休眠性强的品系会被淘汰,因此就间接淘汰了耐贮性强的品系。闫蕴韬等[6]研究发现:水稻种子的休眠性和耐贮性呈正相关,但同时也发现不少休眠性弱而耐贮性强的种质材料,因此可以从水稻种质群体材料中筛选出耐贮性强但休眠性一般或较弱的种质资源,从而选育耐贮性强的水稻新品种。
α-淀粉酶与β-淀粉酶都是必不可少的分解淀粉的酶类。种子的萌发与淀粉酶呈高度正相关,淀粉酶活性越高,种子的萌发力越强[32],这与本研究结果一致:耐贮品种发芽率越高,淀粉酶活性越强;不耐贮品种发芽率低,淀粉酶活性也越低。在植株内,由于萌发需要启动一系列程序性的复杂的生理生化反应,许多活性物质如蛋白质、酶、激素等需要合成,这些合成反应都需要能量,因此较高的淀粉酶活性可以快速水解种子中储存的淀粉,从而提供充分的呼吸底物,以保障种子萌发中的能量供应。随着自然老化造成逆境胁迫加重,引起植株内质膜功能改变,质膜损伤加重,淀粉酶的活性受到抑制,不耐贮品种发芽率、发芽势也呈现出降低的趋势。
种子贮藏过程中受环境的诱导,活性氧(ROS)生成增加,为了消除ROS对蛋白质的损伤而造成的危害[33−35],植物发展了较为复杂的抗氧化体系调节蛋白质的氧化还原状态,以保持其正常的生理功能。其中CAT、SOD和POD等几种主要的酶一起构成了生物的抗氧化酶系统,这几种酶的活性高有利于清除活性氧、自由基的毒害,可以保持种子活力,增强其耐贮性。因此它们的活性高低与植物代谢及抗逆性密切相关[36]。本研究中,不耐贮品种的CAT活性均显著低于耐贮品种,说明不耐贮品种的各类生化反应受到抑制,导致过氧化氢的产量降低,因此该酶活性也逐步降低。不耐贮品种的POD活性均显著低于耐贮品种,这可能是随着逆境胁迫加深细胞代谢活动受到一定抑制,产生的过氧化物量减少,需要此酶的活性降低。不耐贮品种的SOD酶活性均显著低于耐贮品种,说明在不耐贮品种中,水稻种子内生化反应变化剧烈,产生的自由基和超氧阴离子较多,需要此酶较高的活性来应对。
水稻种子在贮藏期间受到逆境胁迫时,MDA质量摩尔浓度急剧增加,植物体通过调节体内脯氨酸含量来适应逆境[37−40]。本研究发现不耐贮品种的MDA质量摩尔浓度均显著高于耐贮品种;不耐贮品种的Pro质量分数均显著低于耐贮品种。说明在贮藏过程中,耐贮品种产生的MDA质量摩尔浓度较少,其膜脂过氧化程度较低,其抗氧化能力强,能减少有害物质的积累,有利于保持种子活力。同时耐贮品种能够通过调节体内Pro质量分数来适应逆境,有效抵御逆境对种子带来的伤害。这可能也是其耐贮藏的重要原因之一。
本研究结果表明:自然老化的水稻品种的耐贮性与淀粉酶、抗氧化酶、丙二醛、脯氨酸等生理指标密切相关。结合发芽数据与生理指标,筛选出一批耐贮品种,即‘南粳44’‘沈稻18’‘天稻丰’‘赣晚籼35号’‘赣晚糯7号’‘甬籼69’‘两优培九’‘中浙优1号’‘晶两优华占’。在今后的育种工作中,可增加以上品种的繁种和制种工作。不管是在常规粳稻、常规籼稻或是杂交稻中,均存在耐贮性好的材料和耐贮性差的材料,这种分布的多样性可以为发掘、利用和聚合耐贮藏基因,选育耐贮藏的水稻品种提供物质与遗传基础。
Screening and physiological characteristics of storable Oryza sativa germplasm resources
-
摘要:
目的 水稻Oryza sativa种子的耐贮性是与种用价值、种源和粮食安全密切相关的重要性状。研究水稻种子耐贮特性,对水稻种质资源保存以及保障国家粮食安全具有重要意义。 方法 基于220份水稻品种,经过5~6 a的自然老化,通过室内标准发芽实验筛选出耐贮水稻品种,并进一步测定不同耐贮品种间的生理特性。 结果 随着自然老化时间的延长,不同水稻品种的发芽率和发芽势均呈现逐步降低的趋势。与种子初始质量相比,常规粳稻、常规籼稻、杂交稻的发芽率平均降低了62.2%、23.4%、45.3%;发芽势平均降低了55.0%、38.1%、50.7%。以发芽率下降幅度<5%且发芽势下降幅度<15%作为耐贮品种的评价标准,筛选出以‘南粳44’‘Nanjing 44’、‘沈稻18’‘Shendao 18’、‘天稻丰’‘Tiandaofeng’为代表的耐贮常规粳稻;‘赣晚籼35号’‘Ganwanxian 35’、‘赣晚糯7号’‘Ganwannuo 7’、‘甬籼69’‘Yongxian 69’为代表的耐贮常规籼稻;‘两优培九’‘Liangyou Peijiu’、‘中浙优1号’‘Zhongzheyou 1’、‘晶两优华占’‘Jingliangyou Huazhan’为代表的耐贮杂交稻。进一步研究发现:以上9个品种具有淀粉酶活性高、抗氧化酶活性高、脯氨酸质量分数高、丙二醛质量摩尔浓度低的特点。 结论 基于耐贮能力与生理特性,推荐‘南粳44’‘沈稻18’‘天稻丰’‘赣晚籼35号’‘赣晚糯7号’‘甬籼69’‘两优培九’‘中浙优1号’‘晶两优华占’等为耐贮种质,这为今后选育耐贮藏的水稻优良品种提供了新资源。图2表6参40 Abstract:Objective This study, with an investigation of the seed storability, an important trait closely related to seed value, provenance and food security, of rice (Oryza sativa), is aimed to survey the storability characteristics of rice seeds so as to promote the conservation of rice germplasm resources and the protection of national food security. Method Based on 220 rice cultivars, after 5−6 years of natural aging, rice varieties with good storage tolerance were selected employing laboratory standard germination experiment before the physiological characteristics of different cultivars were determined. Result (1) With the prolongation of natural aging time, the germination rate and germination potential of different rice cultivars showed a trend of gradual decrease. (2) Compared with the initial seed quality, the germination rates of conventional japonica rice, conventional indica rice and hybrid rice decreased by 62.2%, 23.4% and 45.3% on average, respectively whereas the germination potential decreased by 55.0%, 38.1% and 50.7% on average. (3) Based on the evaluation criteria (varies with a decrease in germination rate <5% and a decrease in germination potential <15% are regarded as storable cultivars), storable conventional japonica rice represented by ‘Nanjing 44’ ‘Shendao 18’ and ‘Tiandaofeng’ were screened out along with storable conventional indica rice represented by ‘Wanxian 35’ ‘Ganwannuo 7’ and ‘Yongxian 69’; the storable hybrid rice represented by ‘Liangyoupeijiu’ ‘Zhongzheyou 1’ and ‘Jingliangyou Huazhan’. (4) It was further found that ‘Nanjing 44’ ‘Shendao 18’ ‘Tiandaofeng’ ‘Ganwanxian 35’ ‘Ganwanuo 7’ ‘Yongxian 69’ ‘Liangyou Peijiu’ ‘Zhongzhenyou 1’ and ‘Jingliangyou Huazhan’ had the characteristics of high amylase activity, high antioxidant enzyme activity, high proline content and low MDA content. Conclusion In terms of the storability and physiological characteristics, ‘Nanjing 44’ ‘Shendao 18’ ‘Tiandaofeng’ ‘Ganwanxian 35’ ‘Ganwannuo 7’ ‘Yongxian 69’ ‘Liangyoupeijiu’ ‘Zhongzheyou 1’ ‘Jingliangyou Huazhan’ are recommended and storable germplasms, providing new resources for the future selection of rice cultivars with storage tolerance. [Ch, 2 fig. 6 tab. 40 ref.] -
Key words:
- Oryza sativa /
- storability /
- seed vigor
-
图 1 不同类型水稻种子在不同贮藏时期的发芽率变化
Figure 1 Changes of germination rate of different types of rice seeds in different storage periods
图 2 不同类型水稻种子在不同贮藏时期的发芽势变化
Figure 2 Changes of germination potential of different types of rice seeds in different storage periods
表 1 不同常规粳稻品种在不同贮藏时期发芽率和发芽势的变化
Table 1. Variation of germination rate and germination potential of different conventional japonica rice cultivars in different storage periods
类型 品种名 2015年 2020年 2021年 发芽率/% 发芽势/% 发芽率/% 发芽势/% 发芽率/% 发芽势/% 耐贮品种 ‘南粳44’ 96.3±0.6 a 91.7±0.6 a 93.3±0.6 a 88.0±2.0 b 84.3±4.2 b 66.0±1.0 c ‘沈稻18’ 97.0±1.0 a 90.0±2.0 a 94.7±1.2 a 81.3±1.5 b 82.3±5.5 b 59.0±2.0 c ‘天稻丰’ 94.7±1.5 a 86.7±1.5 a 91.7±2.3 a 76.3±2.5 b 73.7±5.8 b 59.3±6.4 c 不耐贮品种 ‘常农粳7号’ 97.0±1.0 a 89.3±0.6 a 76.3±2.1 b 66.3±1.5 b 38.0±5.3 c 33.3±2.1 c ‘秀水134’ 97.0±1.0 a 91.3±1.2 a 78.3±2.5 b 59.3±1.5 b 37.3±0.6 c 32.7±1.5 c ‘平安粳稻11号’ 95.7±0.6 a 88.0±1.0 a 81.3±2.5 b 73.0±3.6 b 25.7±1.2 c 22.7±2.1 c 说明:数据为平均值±标准差。不同小写字母表示同一品种在不同贮藏时期间差异显著(P<0.05)  下载: 导出CSV
下载: 导出CSV
表 2 不同常规籼稻品种在不同贮藏时期发芽率和发芽势变化
Table 2. Variation of germination rate and germination potential of different conventional indica rice cultivars in different storage periods
类型 品种名 2015年 2020年 2021年 发芽率/% 发芽势/% 发芽率/% 发芽势/% 发芽率/% 发芽势/% 耐贮品种 ‘赣晚籼35号’ 97.3±2.1 a 90.0±1.0 a 93.3±3.1 a 82.7±1.2 b 84.7±2.5 b 81.3±2.5 b ‘赣晚糯7号’ 96.0±1.0 a 91.3±1.2 a 93.0±1.7 a 85.0±3.6 a 78.3±5.9 b 70.0±4.0 b ‘甬籼69’ 95.0±1.0 a 83.0±2.6 a 91.3±3.2 a 80.7±3.1 a 72.0±7.5 b 63.3±2.1 b 不耐贮品种 ‘温926’ 91.7±1.5 a 88.7±2.1 a 78.3±0.6 b 72.0±2.0 b 52.3±5.1 c 41.3±1.5 c ‘甬籼15号’ 88.7±1.2 a 83.3±0.6 a 72.7±5.0 b 55.3±1.2 b 46.3±4.0 c 37.0±1.0 c ‘扬稻6号’ 87.7±2.3 a 81.3±1.5 a 74.7±2.5 b 62.3±2.5 b 29.7±7.1 c 26.0±1.0 c 说明:数据为平均值±标准差。不同小写字母表示同一品种在不同贮藏时期间差异显著(P<0.05)
下载: 导出CSV
表 3 不同杂交稻品种在不同贮藏时期发芽率变化
Table 3. Changes of germination rate of different hybrid rice cultivars in different storage periods
类型 品种名 2015年 2020年 2021年 发芽率/% 发芽势/% 发芽率/% 发芽势/% 发芽率/% 发芽势/% 耐贮品种 ‘两优培九’ 97.7±1.5 a 89.0±3.0 a 96.3±1.5 a 84.0±2.0 a 92.7±2.1 a 78.0±1.0 b ‘中浙优1号’ 96.3±0.6 a 90.3±1.5 a 94.3±8.6 a 87.3±2.3 a 91.3±3.2 a 82.3±1.5 b ‘晶两优华占’ 96.0±1.0 a 89.0±1.0 a 93.3±0.6 a 86.0±1.0 a 92.3±0.6 a 77.0±1.0 b 不耐贮品种 ‘隆香优130’ 96.3±1.5 a 90.7±1.5 a 71.0±1.7 b 62.0±2.0 b 38.7±2.1 c 28.3±2.5 c ‘深两优865’ 96.0±2.0 a 86.0±1.7 a 79.0±2.0 b 68.7±3.1 b 24.7±3.1 c 18.3±1.5 c ‘两优1号’ 94.0±1.0 a 91.3±2.1 a 75.0±1.0 b 66.3±2.1 b 29.0±1.7 c 20.3±2.1 c 说明:数据为平均值±标准差。不同小写字母表示同一品种在不同贮藏时期间差异显著(P<0.05)
下载: 导出CSV
表 4 不同品种常规粳稻生理指标变化
Table 4. Changes of physiological indexes of different cultivars of conventional japonica rice
类型 品种 α-淀粉酶活性/
(mg·g−1·min−1)β-淀粉酶活性/
(mg·g−1·min−1)CAT活性/
(×16.67 nkat·g−1)POD活性/
(×16.67 nkat·g−1)SOD活性/
(×16.67 nkat·g−1)MDA/
(μmol·kg−1)Pro/
(μg·g−1)耐贮品种 ‘南粳44’ 2.9±0.5 b 5.8±0.6 c 85.7±1.4 a 305.1±1.9 a 212.2±9.7 a 2.7±0.7 d 14.7±2.1 a ‘沈稻18’ 4.3±0.3 a 8.6±0.9 b 63.7±3.3 b 224.3±10.3 b 211.8±1.2 a 4.7±0.2 b 12.3±1.0 b ‘秀水134’ 1.8±0.1 c 12.3±1.8 a 62.4±1.8 b 133.7±3.6 d 118.9±3.1 d 4.0±0.1 bc 10.6±0.5 bc 平均值 3.0±1.3 A 8.9±3.3 A 70.6±13.1 A 221.0±85.7 A 181.0±53.8 A 3.8±1.0 A 12.5±2.1 A 不耐贮品种 ‘平安粳稻11号’ 0.4±0.2 d 5.5±0.3 c 62.9±1.3 b 181.5±2.1 c 122.7±3.5 d 3.3±0.1 cd 9.8±0.5 c ‘常农粳7号’ 0.6±0.1 d 4.6±0.3 c 64.2±0.5 b 182.1±1.7 c 159.6±8.7 c 9.2±0.9 a 9.9±0.4 c ‘天稻丰’ 0.2±0.2 d 5.3±0.1 c 48.4±1.2 c 174.1±3.1 c 174.3±2.9 b 9.7±0.8 a 10.5±1.4 bc 平均值 0.4±0.2 A 5.1±0.5 A 58.5±8.8 A 179.2±4.4 A 152.2±26.6 A 7.4±3.6 A 10.0±0.4 A 说明:数据为平均值±标准差。同列不同小写字母表示各品种间差异显著,同列不同大写字母表示不同耐贮性的平均值差异显著(P<0.05)
下载: 导出CSV
表 5 不同品种常规籼稻生理指标变化
Table 5. Changes of physiological indexes of different cultivars of conventional indica rice
类型 品种 α-淀粉酶活性/
(mg·g−1·min−1)β-淀粉酶活性/
(mg·g−1·min−1)CAT活性/
(×16.67 nkat·g−1)POD活性/
(×16.67 nkat·g−1)SOD活性/
(×16.67 nkat·g−1)MDA/
(μmol·kg−1)Pro/
(μg·g−1)耐贮品种 ‘赣晚籼35号’ 4.5±0.5 a 11.2±1 b 54.9±2.6 c 328.1±11.6 b 210.8±6.7 b 2.3±0.2 e 10.1±0.6 b ‘赣晚糯7号’ 3.7±0.3 b 11.7±1 b 77.1±4.4 b 468.4±10.1 a 224.8±0.3 a 2.4±0.3 e 10.8±1.3 b ‘甬籼69’ 4.3±0.3 a 16.9±1.6 a 104.5±3.0 a 271.8±8.3 c 212.9±5.7 b 5.6±0.2 d 14.1±1.0 a 平均值 4.2±0.4 A 13.3±3.2 A 78.8±24.8 A 356.1±101.2 A 216.2±7.6 A 3.4±1.9 A 11.7±2.1 A 不耐贮品种 ‘农乐1号’ 1.2±0.2 d 6.6±0.4 c 46.3±1.7 d 127.7±10.8 e 211.7±4.8 b 7.2±0.3 b 10.3±1.2 b ‘甬籼15号’ 0.6±0.1 e 4.8±0.4 c 28.2±2.4 e 279.8±4.9 c 176.8±3.9 d 7.8±0.2 a 11.1±0.4 b ‘扬稻6号’ 2.3±0.2 c 5.4±0.5 c 55.0±1.2 c 184.4±1.9 d 202.9±1.7 c 6.7±0.2 c 10.2±0.2 b 平均值 1.4±0.9 B 5.6±0.9 A 43.2±13.7 A 197.3±76.9 B 197.1±18.2 A 7.2±0.6 A 10.5±0.5 A 说明:数据为平均值±标准差。同列不同小写字母表示各品种间差异显著,同列不同大写字母表示不同耐贮性的平均值差异显著(P<0.05)
下载: 导出CSV
表 6 不同品种杂交稻生理指标变化
Table 6. Changes of physiological indexes of different cultivars of hybrid rice
类型 品种 α-淀粉酶活性/
(mg·g−1·min−1)β-淀粉酶活性/
(mg·g−1·min−1)CAT活性/
(×16.67 nkat·g−1)POD活性/
(×16.67 nkat·g−1)SOD活性/
(×16.67 nkat·g−1)MDA/
(μmol·kg−1)Pro/
(μg·g−1)耐贮品种 ‘两优培九’ 8.5±0.2 a 22.4±0.4 a 133.5±5.7 b 201.4±3.7 c 229.9±7.6 a 2.0±0.6 e 12.0±0.4 c ‘中浙优1号’ 4.0±0.7 c 12.9±1.1 b 169.4±3.0 a 376.4±9.5 b 229.6±3.1 a 2.7±0.2 d 31.4±1.3 a ‘晶两优华占’ 6.6±0.2 b 13.5±0.1 b 119.9±1.3 c 430.3±11.3 a 206.9±1.6 b 2.2±0.5 e 16.1±1.8 b 平均值 6.4±2.3 A 16.3±5.3 A 140.9±25.6 A 336.0±119.7 A 222.1±13.2 A 2.3±0.4 A 19.8±10.2 A 不耐贮品种 ‘隆香优130’ 2.7±0.4 d 3.6±0.1 e 60.6±3.3 d 105.3±4.6 f 144.8±3.1 d 14.5±0.5 a 12.6±1.2 c ‘深两优865’ 0.6±0.1 e 8.7±0.2 c 53.8±2.2 d 152.3±4.8 d 175.8±3.7 c 7.5±0.3 c 10.6±0.7 cd ‘两优1号’ 0.9±0.1 e 4.7±0.4 d 60.2±3.4 d 127.5±7.2 e 123.8±2.5 e 12.2±0.4 b 9.4±0.7 d 平均值 1.4±1.1 B 5.7±2.7 A 58.2±3.8 B 128.3±23.5 A 148.1±26.2 B 11.4±3.6 A 10.8±1.6 A 说明:数据为平均值±标准差。同列不同小写字母表示各品种间差异显著,同列不同大写字母表示不同耐贮性的平均值差异显著(P<0.05)
下载: 导出CSV
-
[1] GODFRAY H C, BEDDINGTON J R, CRUTE I R, et al. Food security: the challenge of feeding 9 billion people [J]. Science, 2010, 327(5967): 812 − 818. [2] 吴跃进, 卢义宣, 吴敬德, 等. 耐储藏专用型水稻选育及相关技术研究[J]. 中国稻米, 2004, 10(3): 6 − 7. WU Yuejin, LU Yixuan, WU Jingde, et al. Research on the breeding of storage-tolerant rice and related technologies [J]. China Rice, 2004, 10(3): 6 − 7. [3] 曾大力. 水稻耐贮藏种质的筛选及遗传育种研究[D]. 北京: 中国农业科学院, 2002. ZENG Dali. Genetic Analysis and Improvement of Long-storability in Rice (Oryza sativa L. ) [D]. Beijing: Chinese Academy of Agricultural Sciences, 2002. [4] HANG N T, LIN Qiuyun, LIU Linglong, et al. Mapping QTLs related to rice seed storability under natural and artificial aging storage conditions [J]. Euphytica, 2015, 203(3): 673 − 681. [5] 柳武革, 王丰, 刘振荣, 等. 水稻耐储藏特性研究进展[J]. 生物技术通报, 2006(增刊 1): 50 − 52, 62. LIU Wuge, WANG Feng, LIU Zhenrong, et al. Progress of research on storability of rice [J]. Biotechnology Bulletin, 2006(suppl 1): 50 − 52, 62. [6] 闫蕴韬, 何兮, 张海清, 等. 水稻种子耐贮性研究进展[J]. 中国农学通报, 2022, 38(5): 1 − 8. YAN Yuntao, HE Xi, ZHANG Haiqing, et al. Advances in research on the storability of rice seeds [J]. Chinese Agricultural Science Bulletin, 2022, 38(5): 1 − 8. [7] 熊豆. 水稻种子耐贮性全基因组关联分析[D]. 长沙: 湖南农业大学, 2019. XIONG Dou. Genome-wide Association Study of Rice Seed Storability [D]. Changsha: Hunan Agricultural University, 2019. [8] 李春生. 水稻种子耐贮藏相关性状QTL分析及育种利用评价[D]. 南京: 南京农业大学, 2016. LI Chunsheng. Analysis of QTLs Related to Storability and Their Evaluation of Breeding Utilization in Rice [D]. Nanjing: Nanjing Agricultural University, 2016. [9] 刁丽荣, 魏毅东, 魏林艳, 等. 水稻种子活力和耐储性研究进展[J]. 福建农业学报, 2016, 31(4): 437 − 443. DIAO Lirong, WEI Yidong, WEI Linyan, et al. Research progress of rice seed vigor and storage tolerance [J]. Fujian Journal of Agricultural Sciences, 2016, 31(4): 437 − 443. [10] 江川, 李书柯, 李清华, 等. 水稻耐储藏性研究进展[J]. 江西农业学报, 2011, 23(10): 39 − 43. JIANG Chuan, LI Shuke, LI Qinghua, et al. Research progress in storability of rice [J]. Acta Agriculturae Jiangxi, 2011, 23(10): 39 − 43. [11] 李稳香, 颜启传. 杂交水稻自然老化种子与人工老化种子性能差异研究[J]. 杂交水稻, 1997(3): 29 − 31. LI Wenxiang, YAN Qichuan. Study on the performance difference between natural and artificially aged seeds of hybrid rice [J]. Hybrid Rice, 1997(3): 29 − 31. [12] 董国军, 胡兴明, 曾大力, 等. 水稻种子人工老化和自然老化的比较研究[J]. 浙江农业科学, 2004(1): 29 − 31. DONG Guojun, HU Xingming, ZENG Dali, et al. A comparative study on artificial aging and natural aging of rice seeds [J]. Journal of Zhejiang Agricultural Sciences, 2004(1): 29 − 31. [13] CAO Dongdong, CHEN Shanyu, HUANG Yutao, et al. Effects of artificial aging on physiological characteristics of rice seeds with different dormancy characteristics [J]. Agricultural Biotechnology, 2019, 8(1): 52 − 56. [14] 王仪春, 王洋, 陆敏, 等. 人工老化处理对不同休眠特性水稻种子生理特性的影响[J]. 种子, 2018, 37(6): 15 − 19. WANG Yichun, WANG Yang, LU Min, et al. Effects of artificial aging on physiological characteristics of rice seeds in different dormancy properties [J]. Seed, 2018, 37(6): 15 − 19. [15] 张瑛, 吴先山, 吴敬德, 等. 稻谷储藏过程中理化特性变化的研究[J]. 中国粮油学报, 2003, 18(6): 20 − 24, 28. ZHANG Ying, WU Xianshan, WU Jingde, et al. Study on the change of physical and chemical properties of rice during storage [J]. Journal of the Chinese Cereals and Oils Association, 2003, 18(6): 20 − 24, 28. [16] International Seed Testing Association. International rules for seed testing [J]. Seed Science and Technology, 1985, 31(1): 1 − 152. [17] 刘小丽, 宋保军, 侯睿林. 测定α-淀粉酶活性的两种方法的比较研究[J]. 农业科技与信息, 2006(9): 36 − 38. LIU Xiaoli, SONG Baojun, HOU Ruilin. A comparative study of two methods for the determination of α-amylase activity [J]. Agricultural Science-Technology and Information, 2006(9): 36 − 38. [18] AEBI H. Catalase in vitro [J]. Methods in Enzymology, 1984, 105(C): 121 − 126. [19] KOCHBA J, LAVEE S, SPIEGEL-ROY P. Differences in peroxidase activity and isoenzymes in embryogenic ane non-embryogenic ‘Shamouti’ orange ovular callus lines [J]. Plant and Cell Physiology, 1977, 18(2): 463 − 467. [20] CONSTANTINE N, GIANNOPOLITIS, RIES S K. Superoxide dismutases [J]. Plant Physiology, 1977, 59(2): 309 − 314. [21] HODGES D M, DELONG J M, FORNEY C F, et al. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds [J]. Planta, 1999, 207(4): 604 − 611. [22] 陈丽珍, 叶剑秋, 王荣香. 水稻盐胁迫的研究进展[J]. 热带农业科学, 2011, 31(3): 87 − 90. CHEN Lizhen, YE Jianqiu, WANG Rongxiang. Research progress on salt-stress in rice [J]. Chinese Journal of Tropical Agricultural, 2011, 31(3): 87 − 90. [23] KRANNER I, MINIBAYEVA F V, BECKETT R P, et al. What is stress? concepts, definitions and applications in seed science [J]. New Phytologist, 2010, 188(3): 655 − 673. [24] RAO N K, JACKSON M T. Variation in seed longevity of rice cultivars belonging to different isozyme groups [J]. Genetic Resources and Crop Evolution, 1997, 44(2): 159 − 164. [25] 曾大力, 钱前, 国广泰史, 等. 稻谷储藏特性及其与籼粳特性的关系研究[J]. 作物学报, 2002, 28(4): 551 − 554. ZENG Dali, QIAN Qian, KUNIHIRO Y, et al. Study on storability and morphological index in rice (Oryza sativa L. ) under artificial ageing [J]. Acta Agronomica Sinica, 2002, 28(4): 551 − 554. [26] RAJA K, PONNUSWAMY A S. Shelf life of hybrid rice (CORH-2) seed as influenced by stage of harvest [J]. Journal Seed Research, 2009, 37(1): 81 − 87. [27] ARUMUGAM M, RAJANNA M P, GOWDA R. Seed dormancy and seedling vigour as influenced by planting time environment and date of harvest in rice (Oryza sativa L. ) [J]. Caspian Journal of Environmental Sciences, 2008, 6(1): 1 − 9. [28] 侯文平, 王成瑷, 赵磊, 等. 收获时期对水稻种子发芽率的影响[J]. 种子世界, 2014(12): 35 − 37. HOU Wenping, WANG Cheng’ai, ZHAO Lei, et al. Effect of harvest time on germination rate of rice seeds [J]. Seed World, 2014(12): 35 − 37. [29] KRISHNA A, BIRADARPATIL N K, MANJAPPA K, et al. Influence of methods of cultivation and nutrition on seed storability of BPT-5204 rice variety [J]. Karnataka Journal of Agricultural Sciences, 2009, 22(1): 192 − 193. [30] RAJJOU L, LOVIGNY Y, GROOT S P C, et al. Proteome-wide characterization of seed aging in Arabidopsis: a comparison between artificial and natural aging protocols [J]. Plant Physiology, 2008, 148(1): 620 − 641. [31] ELLIS R H, HONG T D, IACHSON M T. Seed production environment, time of harvest, an the potential longetivity of seeds of three cultivars of rice [J]. Annals of Botany, 1993, 72: 583 − 590. [32] 汪晓峰, 丛滋金. 种子活力的生物学基础及提高和保持种子活力的研究进展[J]. 种子, 1997, 16(6): 36 − 39. WANG Xiaofeng, CONG Zijin. Biological basis of seed vigor and research progress of improving and maintaining seed vigor [J]. Seed, 1997, 16(6): 36 − 39. [33] 高家东. 杂交水稻种子耐贮藏生理基础和蛋白质组学研究[D]. 长沙: 湖南农业大学, 2012. GAO Jiadong. Study on the Physiological Characteristics and Proteomics of Seed Storability of Hybrid Rice [D]. Changsha: Hunan Agricultural University, 2012. [34] KOORNNEEF M, BENTSINK L, HILHORST L. Seed dormancy and germination [J]. Current Opinion in Plant Biology, 2002, 5(1): 33 − 36. [35] 丁海东, 刘慧, 朱晓红, 等. 植物细胞蛋白质氧化及其蛋白质组学研究进展[J]. 中国农学通报, 2011, 27(33): 187 − 193. DING Haidong, LIU Hui, ZHU Xiaohong, et al. Research progress of protein oxidation in plant cells and its proteomics [J]. Chinese Agricultural Science Bulletin, 2011, 27(33): 187 − 193. [36] HAYAT E, CHRISTOPHE B. Oxidative signaling in seed germination and dormancy [J]. Plant Signaling &Behavior, 2008, 3(3): 175 − 182. [37] 南芝润, 范月仙. 植物过氧化氢酶的研究进展[J]. 安徽农学通报, 2008, 14(5): 27 − 29. NAN Zhirun, FAN Yuexian. Research progress of plant catalase [J]. Anhui Agricultural Science Bulletin, 2008, 14(5): 27 − 29. [38] 张冠初, 张智猛, 徐扬, 等. 不同基因型花生抗旱耐盐性评价及鉴定指标筛选[J]. 种子, 2020, 39(8): 1 − 5. ZHANG Guanchu, ZHANG Zhimeng, XU Yang, et al. Evaluation of drought and salt tolerance of peanuts with different genotypes and screening of resistance-related identification indexes [J]. Seed, 2020, 39(8): 1 − 5. [39] 徐宇, 肖化云, 郑能建, 等. 植物组织中游离氨基酸在盐胁迫下响应的研究进展[J]. 环境科学与技术, 2016, 39(7): 40 − 47. XU Yu, XIAO Huayun, ZHENG Nengjian, et al. Progress on responding of free amino acid in plants to salt stress [J]. Environmental Science &Technology, 2016, 39(7): 40 − 47. [40] LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔC t method [J]. Methods, 2001, 25(4): 402 − 408. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220594
 点击查看大图
点击查看大图
计量
- 文章访问数: 2463
- HTML全文浏览量: 377
- PDF下载量: 100
- 被引次数: 0
