




-
山蜡梅Chimonanthus nitens又称亮叶蜡梅、毛山茶、香风茶,为蜡梅科Calycanthaceae蜡梅属Chimonanthus常绿灌木,是中国特有植物,主产于浙江、江西、安徽、湖南、广西、贵州等长江以南省(自治区)。山蜡梅含有挥发油、生物碱、黄酮类和酚性成分,具有抗病毒、消炎、镇痛、提高免疫力等功效,是茶药两用的野生珍稀植物[1]。随着对山蜡梅化学成分和药理活性研究的深入,山蜡梅茶、山蜡梅叶颗粒等制剂的临床应用,山蜡梅需求量不断增加,种群遭到破坏,野生资源不足。为了保证山蜡梅的可持续利用,应开展山蜡梅的资源现状调查和保护工作。湖南[2]、安徽[3]、江西[4]等省先后开展了野生山蜡梅的群落和资源调查。2020年,在对浙江省丽水市遂昌县乌溪江流域公益林典型森林群落多功能经营改造调查中,首次发现浙江省天然山蜡梅种群分布。野生山蜡梅喜湿耐荫,多分布于溪流沟谷和山区林缘。乌溪江流域公益林内山蜡梅种群原生境保存状态完整,是研究浙江居群生境群落特征和多样性的良好群落。从植物群落物种组成和结构探寻山蜡梅种群现状及自我更新能力对于指导山蜡梅资源保护和恢复具有重要理论和实践价值。群落物种多样性研究则可以反映群落中物种的丰富度、均匀度以及群落与环境因子的适合度,是判断群落稳定性和资源利用可持续性的重要依据,对保护和恢复山蜡梅种群,合理开发利用山蜡梅资源具有重要意义。
-
浙江省丽水市遂昌县境内的乌溪江流域(28°37′13″~28°21′21″N,118°55′36″~119°00′32″E)全长约56 km,属于亚热带季风气候,温暖湿润,四季分明,雨量充沛,土壤营养丰富。林分类型有常绿阔叶林、杉木Cunninghamia lanceolata林、针阔混交林等,流域内蕴藏着丰富的森林资源、水资源和生物资源[5],生态环境优越。乌溪江源头部分样地植物有25科42属50种[6]。以山蜡梅为主的蜡梅属植物分布于乌溪江流域林地的中坡和下坡以及林缘和溪流边坡等地。本研究调查样地所在的林分类型均为次生常绿阔叶林。
-
采用典型取样法沿乌溪江流域两侧,自北向南在遂昌县湖山林场孟淤(P1)、左肩(P2)、沙会(P3)、焦滩(P4)和蟠龙(P5)公益林内山蜡梅分布较为集中的区域分别设置400 m2样地各1个。将所有样地分割为4个100 m2的样方进行调查,记录样方内所有胸径≥1 cm的树种名称、位置、胸径、树高、冠幅和枝下高。各样地内均匀设置3个5 m×5 m的小样方,调查灌木层、草本层植物名称、平均高度、株(丛)数和盖度。进行山蜡梅种群年龄结构调查时,记录样地内所有山蜡梅的树高、基径和冠幅。各样地取4份土壤样品,检测氮、磷、钾、有机质质量分数和pH。样地情况和土壤检测结果如表1所示。
表 1 乌溪江流域各样地基本信息
Table 1. Basic information of test plots in Wuxi River Basin
样地 北纬(N) 东经(E) 海拔/m 坡向 坡度/(°) 郁闭度 土壤 氮/(mg·kg−1) 磷/(mg·kg−1) 钾/(mg·kg−1) 有机质/(g·kg−1) pH P1 28°31′58″ 118°56′03″ 250 西 19 0.7 352.5±22.2 a 1.2±0.6 b 99.8±50.1 b 54.3±15.2 a 4.8 P2 28°32′10″ 118°55′44″ 271 西北 45 0.5 281.4±55.0 b 0.8±0.7 b 97.6±25.3 b 57.3±14.3 a 4.8 P3 28°30′57″ 118°58′38″ 260 东北 30 0.7 323.6±43.1 ab 1.5±1.1 b 138.6±35.5 b 55.3±8.4 a 4.9 P4 28°23′51″ 118°58′22″ 261 西南 25 0.6 368.2±45.9 a 3.1±0.4 a 251.7±45.7 a 64.9±14.8 a 4.8 P5 28°24′03″ 118°58′51″ 326 西南 50 0.6 292.3±32.8 b 0.7±0.6 b 105.1±23.0 b 44.6±8.4 a 5.0 说明:不同小写字母表示不同样地间差异显著(P<0.05)。 -
参考《中国植物志》网站(
http://www.iplant.cn/frps )命名和分类标准[7],统计样地内所有植物的科、属、种以及生活型。按世界种子植物科、属分布区类型划分标准[8−9]划分样地群落中科、属的分布区类型。 -
采用空间代替时间的方法,以径级代替龄级进行年龄划分。结合各样方乔木层种类胸径的调查数据,分别以胸径(DBH)和树高(H)为指标,绘制乔木层各种类种群的径级垂直结构图。以样地内1 cm≤DBH≤5 cm的乔木作第Ⅰ径级,以5 cm为级长,5 cm<DBH≤10 cm的乔木作第Ⅱ径级,……,DBH>25 cm作为第Ⅵ径级,分级统计株数,计算百分率。其中,Ⅰ~Ⅱ为小径木,Ⅲ~Ⅳ为中径木,Ⅴ~Ⅵ为大径木。山蜡梅为灌木,测量其基径(DB),以0.5 cm为1级,1.0 cm<DB≤1.5 cm为第1级,……, DB>4.5 cm为第8级,绘制山蜡梅径级结构图。采用桂旭君等[10]的方法,精准测量样地内全部DBH≥1 cm的个体树高,对树高进行K-means聚类分析,以不同分类数下组内平方和的变化情况确定最佳分类数,代入K-means分类法中获得乔木层各层高度范围。
-
山蜡梅生活型为灌木,但其整体高度与所处林分乔木层的高度未有明显分层,与乔木层分别统计会明显影响同一群落层间物种重要值的可比性[11],因此,在样地调查和计算重要值时,将山蜡梅、檵木Loropetalum chinense等与乔木层未有明显分层的灌木或小乔木按乔木层计算。计算样地内各层组成树种的相对密度、相对频度、相对优势度和相对盖度,并计算重要值,具体参考文献[12−14]。
参考杨泉光等[15]的分析方法,采用Drude 7级制多度估测灌木层和草本层物种个体数目:Soe表示植株地上部密闭,覆盖度为76%~100%;Cop3表示植株很多,但个体未完全衔接,覆盖度为51%~75%;Cop2表示植株多,覆盖度为26%~50%;Cop1表示植株尚多,覆盖度为6%~26%;Sp表示植株不多而分散,覆盖度为1%~5%;Sol表示植株数很少而稀疏,覆盖度<1%;Un表示个别或单株。
-
参考汪殿蓓等[16]的植物群落多样性研究指数选择标准,采用ForStat 2.0 (统计之林)计算山蜡梅群落各种多样性指数。指数包括:物种丰富度指数[样地内物种数(N)和Margalef指数(R)];物种多样性指数[Shannon-Wienner指数(H)和Simpson 指数(D)];Pielou均匀度指数(J);优势度指数(C)。参照袁丛军等[17]的方法,将乔木层、灌木层、草本层权重系数分别设定为0.5、0.3、0.2,计算各样地整体多样性指数。
-
如表2所示:乌溪江山蜡梅生境群落中的种子植物包含42科71属,涉及12个分布区类型。属于世界分布和中国特有分布的各3属,均占总属数的4.23 %,其中本研究中的山蜡梅即属于中国特有分布中的蜡梅属;热带分布(2~7)的有36属,占总属数的50.70%,其中,泛热带分布有15属,占21.13%;温带分布(8~11)的有29属,占总属数的40.85%,多数属集中在东亚和北美洲间断分布(12属)和北温带分布(10属),分别占总属数的16.90%和14.08%。未见温带亚洲分布,地中海区、西亚至中亚分布和中亚分布等类型。由此可见,山蜡梅植物地理成分有热带向温带的过渡趋势,具有典型的中亚热带森林植物区系特点。
表 2 乌溪江山蜡梅群落种子植物属的分布区类型
Table 2. Areal-types of spermatophytic genera of C. nitens community in the Wuxi River Basin
分布类型 编号 分布区类型 属数/属 属的比例/% 小计/% 1 世界分布 3 4.23 4.23 热带分布 2 泛热带分布 15 21.13 50.70 3 热带亚洲和热带美洲间断分布 2 2.82 4 旧世界热带分布 6 8.45 5 热带亚洲至热带大洋州分布 2 2.82 6 热带亚洲至热带非洲分布 5 7.04 7 热带亚洲分布 6 8.45 温带分布 8 北温带分布 10 14.08 40.85 9 东亚和北美洲间断分布 12 16.90 10 旧世界温带分布 1 1.41 11 东亚分布 6 8.45 12 中国特有分布 3 4.23 4.23 合计 71 100.00 100.00 -
在乌溪江5个山蜡梅集中分布样地内,共有维管束植物105种,隶属47科80属。其中,被子植物39科68属93种,裸子植物2科3属3种,蕨类植物6科9属9种(表3)。105种植物中乔木为30种,乔木或灌木为32种,灌木为21种,藤本为5种,草本为17种,分别占群落物种总数的 28.57%、30.48%、20.00%、4.76% 和16.19%。组成群落的优势科有:樟科Lauraceae 4属10种;蔷薇科Rosaceae 4属7种;壳斗科Fagaceae 4属6种;豆科Fabaceae 5属5种;茜草科Rubiaceae 5属5种;有2~4个种的有17科,占科总数的36.17%,其余25科均只含1属1种,占总科数的53.19%。
表 3 乌溪江山蜡梅生境群落的维管束植物统计
Table 3. Statistics of vascular plants in the C. nitens community in the Wuxi River Basin
科 属 种 类群 科 属 种 类群 豆科 Fabaceae 5 5 被子植物 虎耳草科 Saxifragaceae 1 1 被子植物 茜草科 Rubiaceae 5 5 被子植物 夹竹桃科 Apocynaceae 1 1 被子植物 樟科 Lauraceae 4 10 被子植物 金缕梅科 Hamamelidaceae 1 1 被子植物 蔷薇科 Rosaceae 4 7 被子植物 堇菜科 Violaceae 1 1 被子植物 壳斗科 Fagaceae 4 6 被子植物 菊科 Asteraceae 1 1 被子植物 禾本科 Poaceae 4 4 被子植物 鳞始蕨科 Lindsaeaceae 1 1 被子植物 大戟科 Euphorbiaceae 3 4 被子植物 马鞭草科 Verbenaceae 1 1 被子植物 报春花科 Primulaceae 3 3 被子植物 葡萄科 Vitaceae 1 1 被子植物 漆树科 Anacardiaceae 3 3 被子植物 山茱萸科 Cornaceae 1 1 被子植物 杜鹃花科 Ericaceae 2 4 被子植物 柿科 Ebenaceae 1 1 被子植物 山茶科 Theaceae 2 4 被子植物 鼠李科 Rhamnaceae 1 1 被子植物 安息香科 Styracaceae 2 2 被子植物 五加科 Araliaceae 1 1 被子植物 桑科 Moraceae 2 2 被子植物 蕈树科 Altingiaceae 1 1 被子植物 冬青科 Aquifoliaceae 1 3 被子植物 杨梅科 Myricaceae 1 1 被子植物 莎草科 Cyperaceae 1 3 被子植物 叶下珠科 Phyllanthaceae 1 1 被子植物 五列木科 Pentaphylacaceae 1 3 被子植物 柏科 Cupressaceae 2 2 裸子植物 蜡梅科 Calycanthaceae 1 2 被子植物 松科 Pinaceae 1 1 裸子植物 木犀科 Oleaceae 1 2 被子植物 里白科 Gleicheniaceae 2 2 蕨类植物 五福花科 Adoxaceae 1 2 被子植物 鳞毛蕨科 Dryopteridaceae 2 2 蕨类植物 菝葜科 Smilacaceae 1 1 被子植物 海金沙科 Lygodiaceae 1 1 蕨类植物 唇形科 Lamiaceae 1 1 被子植物 金星蕨科 Thelypteridaceae 1 1 蕨类植物 大麻科 Cannabaceae 1 1 被子植物 卷柏科 Selaginellaceae 1 1 蕨类植物 杜英科 Elaeocarpaceae 1 1 被子植物 乌毛蕨科 Blechnaceae 1 1 蕨类植物 胡桃科 Juglandaceae 1 1 被子植物 -
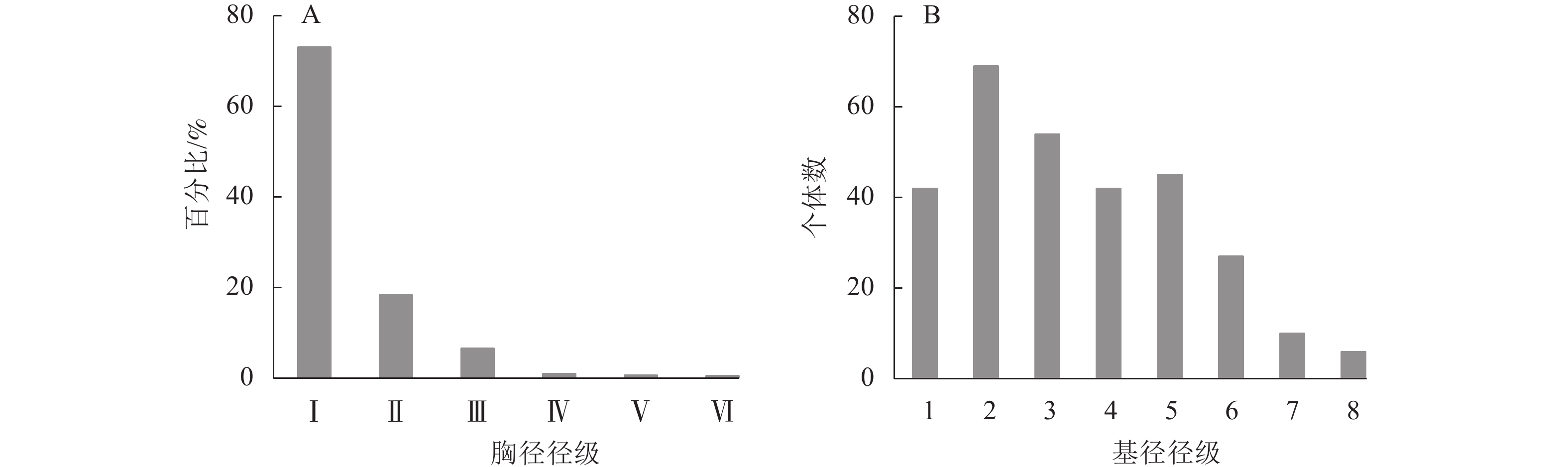
图1A显示:样地内DBH≥1 cm的木本植物共有1276株,其中:小径木的个体占绝大多数,为总株数的91.46%;其次为中径木的个体,占总株数的7.68%;大径木仅有11株,占0.86%,其中水杉Metasequoia glyptostroboides、马尾松Pinus masonniana各3株,杉木2株,苦槠Castanopsis sclerophylla 2株,化香树Platycarya strobilacea 1株,最大径级为水杉(44.0 cm)。样地内所有木本植物个体径级结构呈倒“J”型,整体群落结构为增长型,更新状态良好。山蜡梅主要位于第Ⅰ径级,占总株数的23.12%,为调查样地内的优势种。山蜡梅种群的径级结构同所在群落整体径级结构一致,山蜡梅幼龄和中龄个体(第1~6级)占整个种群的94.58%,为增长型种群(图1B)。
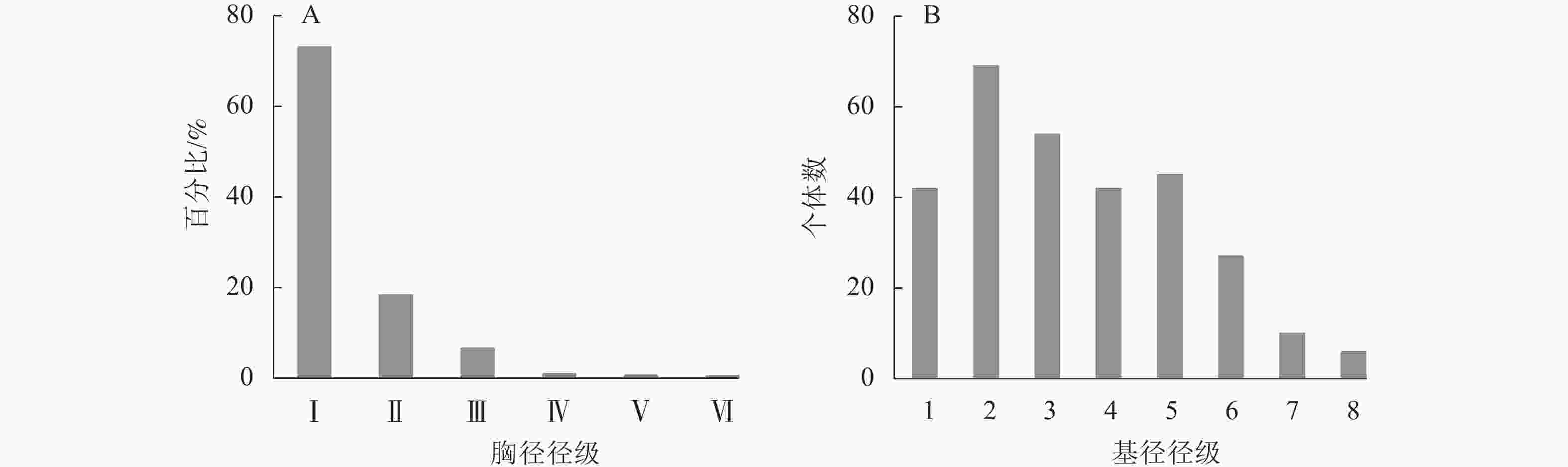
图 1 山蜡梅生境群落(A)和种群(B)径级结构分布
Figure 1. Community (A) and population (B) diameter class structure distribution of C. nitens
乌溪江山蜡梅各样地乔木层植物有76种,重要值如表4所示。山蜡梅所在群落内乔木层重要值总体排序杉木列第1位(11.67%),山蜡梅次之(11.63%),苦槠列第3位(9.05%)。重要值大于2的树种还包括檵木(7.28%)、石栎Lithocarpus glaber (7.08%)、青冈Cyclobalanopsis glauca (5.41%)、峨眉鼠刺Itea oblonga (4.97%)、黄瑞木Adinandra millettii (4.04%)、水杉(3.32%)和木蜡树Toxicodendron sylvestre (2.93%)。不同样地内,山蜡梅所占的重要值位置不同,优势度也不相同。在P1样地内,山蜡梅重要值列第1位(15.89%),P2样地内居第2位(19.36%),P3和P4样地内居第4位(7.59%和7.81%),P5样地内居第3位(13.98%)。由重要值分析亦可知:无论是总体排序还是各样地分别排序,前5位种群的重要值总和均大于50%,说明该群落具有明显的优势群,且山蜡梅种群重要值均排在前4位,表明其为群落的优势种。
表 4 乌溪江山蜡梅生境群落乔木层主要物种的重要值
Table 4. Importance values of the main species in the arbor layer of C. nitens community in the Wuxi River Basin
树种 重要值/% P P1 P2 P3 P4 P5 杉木 Cunninghamia lanceolata 11.67 9.77 4.29 14.91 16.16 − 山蜡梅 Chimonanthus nitens 11.63 15.89 19.36 7.59 7.81 13.98 苦槠 Castanopsis sclerophylla 9.05 5.44 3.16 17.71 9.94 − 檵木 Loropetalum chinense 7.28 12.17 7.82 4.33 3.31 11.94 石栎 Lithocarpus glaber 7.08 − 3.26 8.61 14.07 2.90 青冈 Cyclobalanopsis glauca 5.41 − 20.32 6.52 − 14.43 峨眉鼠刺 Itea oblonga 4.97 3.01 4.01 5.66 7.05 3.79 黄瑞木 Adinandra millettii 4.04 4.92 − 3.74 6.94 − 水杉Metasequoia glyptostroboides 3.32 10.71 − − − − 木蜡树Toxicodendron sylvestre 2.93 3.62 11.68 2.30 − 3.72 马尾松Pinus massoniana 5.63 − − − − 牡荆Vitex negundo var. cannabifolia 3.84 − − − − 拟赤杨Alniphyllum fortunei 2.22 − 4.15 − − 格药柃Eurya muricata 2.12 − − − − 化香树 Platycarya strobilacea 2.08 − − − − 红楠Machilus thunbergii − − − 2.71 − 黄绒润楠Machilus grijsii − − − − 2.19 柳叶蜡梅Chimonanthus salicifolius − − − − 3.10 马银花Rhododendron ovatum − 5.92 − − 3.25 毛冬青Ilex pubescens − − − 2.07 − 毛花连蕊茶Camellia fraterna − − 2.00 3.07 2.75 密花树Rapanea neriifolia − − − − 17.05 山乌桕Triadica cochinchinensis − 7.60 − − − 水团花 Adina pilulifera − − − − 7.94 盐肤木Rhus chinensis − 7.94 − − − 迎春樱Cerasus discoidea − − − 2.02 − 硬斗石栎Lithocarpus hancei − − − − 2.57 中华杜英Elaeocarpus chinensis − 3.17 − − − 说明:48种重要值小于2的树种省略; −表示未分布。P为所有样地树种重要值综合排序;P1~P5为各样地树种重要值排序。 从表5可见:灌木层植物有45种,包括青冈、绒毛润楠Machilus velutina等乔木树种的幼树和杜茎山Maesa japonica、檵木、峨眉鼠刺、香花崖豆藤Millettia dielsiana、白花苦灯笼Tarenna mollissima、山蜡梅、水团花Adina pilulifera等灌木和小乔木,多度大多为Cop1和Cop2。
表 5 乌溪江山蜡梅灌木层主要物种重要值和多度
Table 5. Importance values and abundance of C. nitens community under growth layer in the Wuxi River Basin
树种 株数或丛数 盖度/% 频率 重要值/% 多度 杜茎山 Maesa japonica 29 74.0 0.2 10.17 Cop3 檵木 Loropetalum chinense 14 78.0 0.2 7.58 Soe 青冈 Cyclobalanopsis glauca 10 61.0 0.2 5.91 Cop3 峨眉鼠刺 Itea oblonga 6 43.0 0.2 4.19 Cop2 香花崖豆藤 Millettia dielsiana 8 32.0 0.2 3.97 Cop2 白花苦灯笼 Tarenna mollissima 7 33.4 0.2 3.86 Cop2 山蜡梅 Chimonanthus nitens 8 26.0 0.2 3.65 Cop2 水团花Adina pilulifera 5 30.0 0.2 3.30 Cop2 窄基红褐柃Eurya rubiginosa var. attenuata 6 20.0 0.2 2.95 Cop1 毛花连蕊茶Camellia fraterna 9 9.0 0.2 2.91 Cop1 黄瑞木Adinandra millettii 5 17.0 0.2 2.60 Cop1 绒毛润楠 Machilus velutina 2 26.0 0.2 2.52 Cop2 网脉酸藤子Embelia rudis 5 14.0 0.2 2.43 Cop1 小槐花Ohwia caudata 4 15.0 0.2 2.30 Cop1 映山红Rhododendron simsii 5 9.0 0.2 2.16 Cop1 说明:30种重要值小于2的树种省略。 从表6可知:草本层植物有18种,其中狗脊蕨Woodwardia japonica、美丽复叶耳蕨Arachniodes speciosa、里白Diplopterygium glaucum、芒萁Dicranopteris pedata、山类芦Neyraudia montana和淡竹叶Lophatherum gracile的重要值之和占64.88%。
表 6 乌溪江山蜡梅草本层主要物种重要值和多度
Table 6. Importance values and abundance of of C. nitens community herbal layer in the Wuxi River Basin
树种 株数或丛数 盖度/% 频率 重要值/% 多度 狗脊蕨 Woodwardia japonica 12 87.0 0.6 13.53 Soe 美丽复叶耳蕨 Arachniodes speciosa 14 66.0 0.4 11.38 Cop3 里白 Diplopterygium glaucum 11 77.0 0.4 11.21 Soe 芒萁 Dicranopteris pedata 14 41.0 0.4 9.69 Cop2 山类芦 Neyraudia montana 8 86.0 0.2 9.58 Soe 淡竹叶 Lophatherum gracile 20 31.0 0.2 9.49 Cop2 青绿薹草Carex breviculmis 5 27.0 0.4 6.01 Cop2 珠穗薹草Carex ischnostachya 4 12.0 0.4 4.69 Cop1 微糙三脉紫菀Aster ageratoides 3 10.0 0.2 2.92 Cop1 芒Miscanthus sinensis 2 12.0 0.2 2.75 Cop1 紫花堇菜Viola grypoceras 4 2.0 0.2 2.68 Sp 异穗卷柏Selaginella heterostachys 3 6.0 0.2 2.65 Cop1 三穗薹草Carex tristachya 1 14.0 0.2 2.58 Cop1 黑足鳞毛蕨Dryopteris fuscipes 2 7.0 0.2 2.41 Cop1 乌蕨Odontosoria chinensis 3 1.0 0.2 2.32 Sp 金星蕨Parathelypteris glanduligera 2 4.0 0.2 2.21 Sp 显子草Phaenosperma globosum 1 8.0 0.2 2.18 Cop1 海金沙Lygodium japonicum 1 1.0 0.2 1.70 Sp -
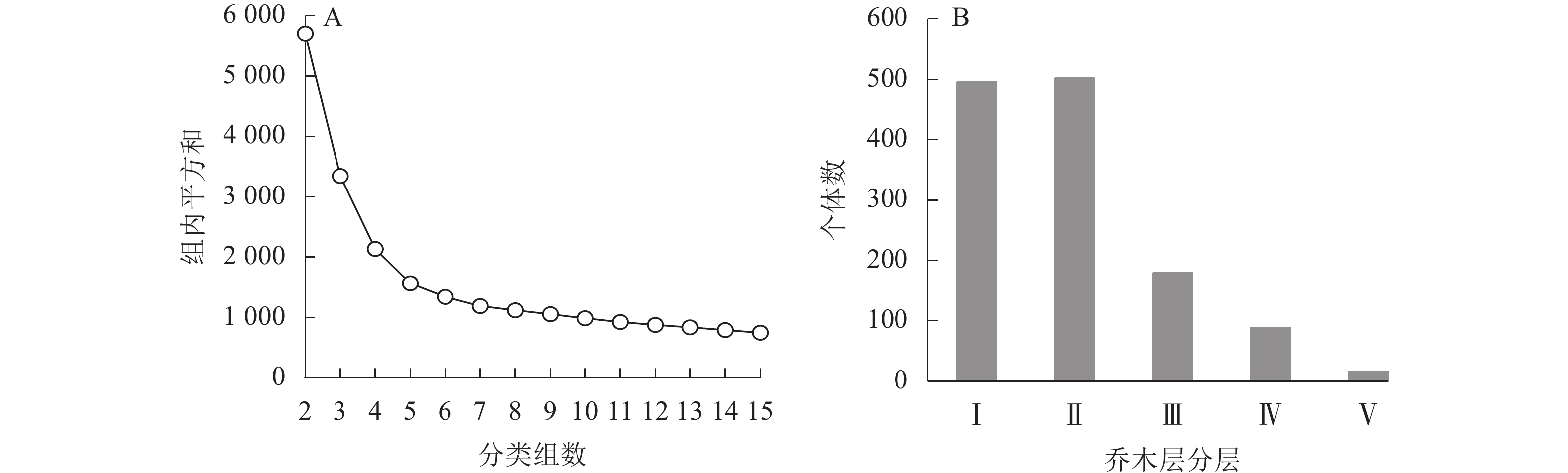
乌溪江山蜡梅群落可分为乔木层、灌木层和草本层。乔木层树种丰富,垂直结构较为复杂,根据不同分类组数下平方和的变化情况对DBH≥1 cm的乔木层再次进行分类。由图2A可知:当分类组数为5时,平方和变化趋势减缓,可确定最优树高聚类组数为5类,代入K-means分类法中,获得山蜡梅所在群落乔木层垂直结构:从下到上依次为乔木Ⅰ层[1.0,4.5) m、乔木Ⅱ层[4.5,7.0) m、乔木Ⅲ层[7.0,10.0) m、乔木Ⅳ层[10.0,14.0) m和乔木Ⅴ层[14.0,18.0] m。各层乔木数量见图2B。乔木Ⅰ层个体占38.87%,全部为小径木,山蜡梅、檵木、青冈、密花树、峨眉鼠刺等47种乔灌木集中于该层,该层中35.08%为山蜡梅。乔木Ⅱ层的个体占39.34%,除2.00%为中径木外,其余均为小径木,主要为山蜡梅、檵木、黄瑞木、峨眉鼠刺、杉木和苦槠等49种,有50.00% 以上的树种与灌木层重合,山蜡梅在该层中占23.71%;乔木Ⅲ层的个体占14.03%,由75.42%的小径木和24.58%的中径木组成,主要为杉木、石栎、苦槠、青冈、木蜡树和拟赤杨等24种;乔木Ⅳ层的个体占6.50%,小径木占43.37%,中径木的比例提高至54.22%,并有少量大径木,主要为杉木和石栎,还包括木蜡树、拟赤杨、红楠等13种;乔木Ⅴ层的个体仅占1.25%,包含小径木、中径木各3株,大径木10株,主要为水杉、马尾松、杉木、拟赤杨和苦槠。灌木层垂直高度为0.80~1.50 m,草本层垂直高度为0.02~0.78 m,种类、株数和相对盖度见表5~6。此外,还有少量层间植物,如流苏子Coptosapelta diffusa、紫藤Wisteria sinensis、络石Trachelospermum jasminoides和异叶爬山虎Parthenocissus dalzielii等。
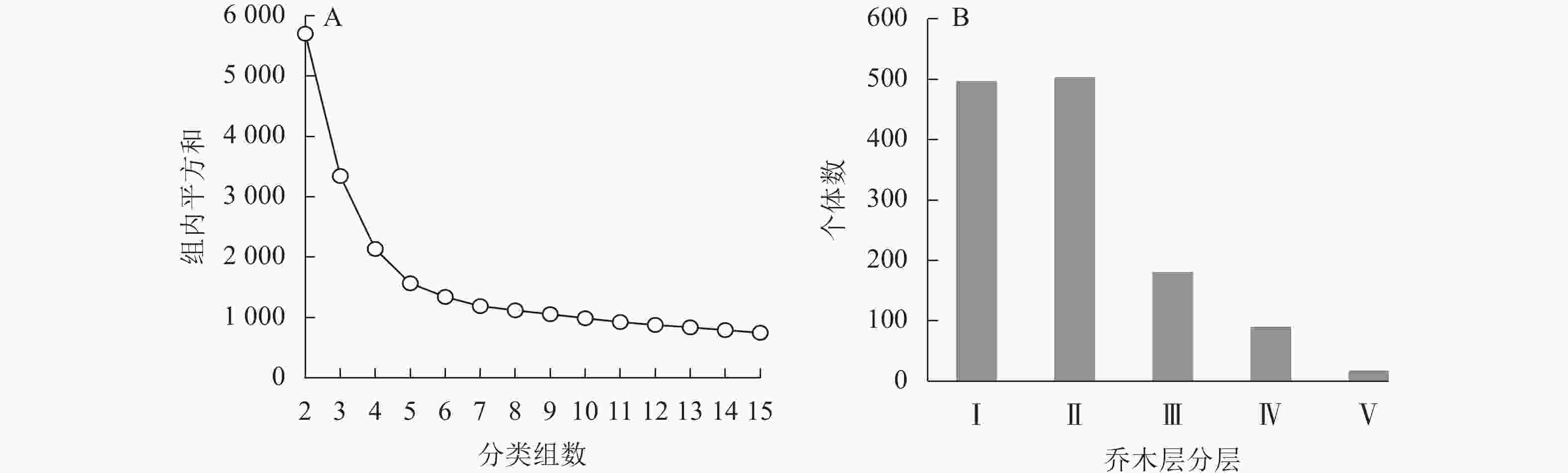
图 2 乔木层最优分类组数判断(A)及各层林木个体数(B)
Figure 2. Judgment index partitioning optimal groups (A) and the number of individuals at each level of the arbor layer (B)
山蜡梅主要位于群落乔木层的乔木Ⅰ层和乔木Ⅱ层,其中,57.84%的山蜡梅位于乔木Ⅰ层,41.46%的山蜡梅位于乔木Ⅱ层,还有少量幼苗位于灌木层。山蜡梅的生活型和灌木特质决定了其在群落中的层次位置,而其所在群落高层植物(乔木Ⅲ层至乔木Ⅴ层)数量显著减少,郁闭度为0.5~0.7时更有利于林下灌层植物如山蜡梅、檵木等的生长。
-
如表7所示:不同样地乔木层、灌木层、草本层的Margalef丰富度指数分别为2.99~5.71、0.93~3.66、1.00~1.86,P1乔木层和草本层丰富度指数最高,P4灌木层丰富度指数最高,整体丰富度指数从大到小依次为P4 (3.93)、P1 (3.84)、P3 (3.73)、P5 (2.68)、P2 (1.98)。不同样地乔木层、灌木层、草本层的Shannon-Wiener指数分别为1.97~2.71、1.09~2.50、1.16~1.61,P3乔木层的Shannon-Wiener指数最高,P4灌木层的Shannon-Wiener指数最高,P1草本层的Shannon-Wiener指数最高,整体Shannon-Wiener指数从大到小依次为P4 (2.37)、P3 (2.20)、P1 (1.94)、P5 (1.83)、P2 (1.56);各样地乔木层、灌木层、草本层的的Simpson指数分别为0.80~0.91、0.60~0.90、0.63~0.75,P3乔木层的Simpson指数最高,P4灌木层和草本层的Simpson指数最高,整体Simpson指数从大到小依次为P4 (0.87)、P3 (0.84)、P5 (0.77)、P1 (0.75)、P2 (0.72)。各样地乔木层、灌木层、草本层的Pielous指数分别为0.67~0.80、0.67~0.92、0.77~0.99,P3乔木层的Pielous指数最高,P4灌木层和草本层的Pielous指数最高,整体均匀度指数从大到小依次为P4 (0.86)、P3 (0.84)、P2 (0.77)、P5 (0.76)、P1 (0.69)。各样地乔木层、灌木层、草本层的优势度指数分别为2.99~5.71、0.93~3.66、1.00~1.86,P1乔木层和草本层的优势度指数最高,P4灌木层的优势度指数最高,整体优势度指数从大到小依次为P4 (3.85)、P1 (3.84)、P3 (3.73)、P5 (2.68)、P2 (1.98)。
表 7 乌溪江山蜡梅生境群落物种多样性指数
Table 7. Diversity indices of C. nitens community in the Wuxi River Basin
样地 乔木层 灌木层 草本层 N R H D J C N R H D J C N R H D J C P1 34 5.71 2.35 0.82 0.67 5.71 9 2.03 1.47 0.64 0.67 2.03 8 1.86 1.61 0.72 0.77 1.86 P2 16 2.99 1.97 0.80 0.71 2.99 4 0.93 1.09 0.60 0.79 0.93 4 1.04 1.26 0.69 0.91 1.04 P3 30 5.35 2.71 0.91 0.80 5.35 10 2.67 2.05 0.84 0.89 2.67 4 1.25 1.16 0.64 0.84 1.25 P4 31 5.27 2.67 0.90 0.78 5.10 15 3.66 2.50 0.90 0.92 3.66 4 1.00 1.38 0.75 0.99 1.00 P5 21 3.71 2.23 0.85 0.73 3.71 7 1.82 1.54 0.73 0.79 1.82 5 1.38 1.24 0.63 0.77 1.38 说明:N. 物种数(种);R. Margalef丰富度指数;H. Shannon-Wiener指数; D. Simpson指数; J. Pielou指数; C. 优势度指数。 乌溪江山蜡梅群落Margalef丰富度指数、Shannon-Wiener指数、Simpson指数和优势度指数从大到小总体表现为乔木层、灌木层、草本层,Pielou指数则呈相反趋势。P4样地的综合多样性指数均最高,P2样地除均匀度指数居中外,各指数均表现较低。
-
乌溪江流域的山蜡梅种群主要分布在海拔250~350 m河流两岸林缘、溪流边坡和林地中、下坡,半阴的缓坡分布数量最多。山蜡梅生境群落中共有维管束植物105种,隶属47科80属。地理区系组成物种以泛热带分布、东亚和北美洲间断分布、北温带分布为主,热带分布属与温带分布属比例分别为50.70%和40.85%,优势科为樟科、蔷薇科、壳斗科、豆科和茜草科等,具有典型中亚热带常绿阔叶林植物区系特点。乌溪江流域属于亚热带季风气候,温暖湿润,地理位置所处气候区与区系性质相符,群落植物对环境有较强的适应性,这与湖南[2]、安徽[3]和江西[4]等地发现的山蜡梅种群所处气候区及生境群落植物区系相一致。
乌溪江山蜡梅所在群落的径级结构与垂直结构表现出与其他亚热带常绿阔叶林样地的一致性,如:样地内所有木本植物个体及山蜡梅种群的径级结构均呈倒“J”型,群落与种群结构为增长型[18−20];不同冠层的径级分布,低冠层(乔木Ⅰ层和乔木Ⅱ层)绝大部分为小径木,中冠层(乔木Ⅲ层和乔木Ⅳ层)先以小径木为主,后随冠层增高中径木比例成倍增加,高冠层(乔木Ⅴ层)则以大径木为主,但数量在整个群落中占比极少[19, 21]。林冠结构能影响林下光照分布,同时也为林下物种提供更多的生态位空间[22]。山蜡梅种群为低冠层的优势种,上层空间中、大径级林木的低占比及以松、杉类等透光率强的针叶树为主的树种组成结构使得林下郁闭度保持在50%~70%的适宜范围,为山蜡梅等耐荫物种早期生长发育和更新提供了蔽护条件。
很多学说认为:群落物种多样性增加可以提高生态系统的稳定性[23−25],但对有些群落而言则为负相关[16, 26]。山蜡梅种群适生冠层的植物数量占林木总数的78.21%,因此,群落多样性高的同时意味着种间竞争强度增大,如P4样地的多样性指数最高,山蜡梅在此样地中的重要值就比在其他样地中低;而在立地条件差、多样性较低的P2样地,山蜡梅的重要值则较高。由此也可知山蜡梅具有极强的生存能力。当条件适宜时,如在P1样地,半阴坡,坡度低于20°,物种多样性较为丰富的条件下,山蜡梅的绝对数量是其他样地的2倍以上。在没有外部人为干扰的情况下群落稳定。建议在保护现有群落的同时,对山蜡梅种群进行遗传多样性研究,选择核心资源,在乌溪江流域开展山蜡梅仿野生栽培试验,合理开发利用山蜡梅野生资源。
Habitat community characteristics and species diversity of Chimonanthus nitens in Wuxi River Basin, Suichang
-
摘要:
目的 山蜡梅Chimonanthus nitens是中国特有的茶药两用野生珍稀植物。随着对山蜡梅药用功能的开发和市场需求的增加,野生资源受到严重破坏。研究山蜡梅种群生境群落的结构特征及物种多样性,探讨山蜡梅的适生环境,可为山蜡梅种群的保护和开发利用提供科学依据。 方法 在实地踏查的基础上,于遂昌乌溪江流域选取山蜡梅典型群落,设置5块400 m2样地,调查分析该群落的植物区系特征、物种组成、群落结构及物种多样性。 结果 ①山蜡梅生境群落区系组成物种以泛热带分布、东亚和北美洲间断分布、北温带分布为主,热带与温带分布属比例分别为50.70%和40.85%,具有典型中亚热带植物区系特点。② 2 000 m2样地内共有维管植物47科80属105种,科属种类丰富、组成分散,优势科为樟科Lauraceae、蔷薇科Rosaceae、壳斗科Fagaceae、豆科Fabaceae和茜草科Rubiaceae。③山蜡梅种群和生境群落径级结构均呈倒“J”型,为增长型结构,乔木层重要值前5位的树种为杉木Cunninghamia lanceolata (11.67%)、山蜡梅(11.63%)、苦槠Castanopsis sclerophylla (9.05%)、檵木 Loropetalum chinense (7.28%)和石栎Lithocarpus glaber (7.08%)。群落垂直结构层次丰富。运用K-means聚类,乔木层胸径≥1 cm的个体根据树高可分为5个林层,山蜡梅主要位于乔木Ⅰ层[1.0, 4.5) m和乔木Ⅱ层[4.5, 7.0) m。④不同样地物种多样性指数差异较大,Margalef指数、Shannon-Wiener指数、Simpson指数、Pielou均匀度指数及优势度指数分别为1.98~3.84、1.56~2.37、0.72~0.87、0.69~0.86、1.98~3.85。 结论 遂昌乌溪江流域山蜡梅生境群落物种组成丰富,垂直结构复杂,小、中径级林木占绝大多数,为增长型群落,物种多样性整体较高。建议在保护现有群落生境前提下,在乌溪江流域开展山蜡梅仿野生栽培试验,合理开发利用山蜡梅资源。图2表7参26 Abstract:Objective Chimonanthus nitens is a rare wild plant unique to China both for tea and medicine. The wild resources have been seriously damaged with the development of its medicinal function and the increase of market demand. This study aims to explore the structure characteristics and species diversity of the habitat community of C. nitens population, as well as its suitable living environment, so as to provide a scientific basis for the protection, development and utilization of C. nitens population. Method Based on the field survey, the typical community of C. nitens was selected in Wuxi River Basin of Suichang, and five 400 m2 plots were set up to investigate and analyze the flora, species composition, community structure and species diversity of the community. Result (1) The flora of C. nitens community was dominated by pantropical distributions, disjuncted distribution in East Asia and North America, and northern temperate distribution. The proportions of tropical and temperate genera were 50.70% and 40.85%, respectively, with typical characteristics of mid-subtropical flora. (2) There were 105 species in the 2000 m2 sample plots, belonging to 80 genera and 47 families. The families and genera were rich in species and dispersed in composition. The dominant families were Lauraceae, Rosaceae, Fagaceae, Fabaceae and Rubiaceae. (3) The diameter class structure of C. nitens population and habitat community was an inverted “J” type, indicating a growth type. The top 5 tree species with important values of the arbor layer were Cunninghamia lanceolata (11.67%), Chimonanthus nitens (11.63%), Castanopsis sclerophylla (9.05%), Loropetalum chinense (7.28%) and Lithocarpus glaber (7.08%). The vertical structure of the community was rich. K-means clustering showed that individuals with DBH≥1 cm in the tree layer could be divided into five forest layers according to the tree height. The main location of the community was in the tree layer Ⅰ [1.0, 4.5) m and tree layer Ⅱ [4.5, 7.0) m. (4) The species diversity index varied greatly in different regions. Margalef index, Shannon-Wiener index, Simpson index, Pielou index and Dominance index were 1.98−3.84, 1.56−2.37, 0.72−0.87, 0.69−0.86 and 1.98−3.85, respectively. Conclusion The community of wild Chimonanthus nitens in Wuxi River Basin of Suichang has rich species composition and complex vertical structure. Small and medium diameter trees account for the vast majority, which is an increasing community with high species diversity as a whole. Under the premise of protecting the existing community habitat, it is suggested to carry out experiments imitating wild cultivation to rationally develop and utilize the wild resources of Chimonanthus nitens. [Ch, 2 fig. 7 tab. 26 ref.] -
Key words:
- Chimonanthus nitens /
- flora /
- species composition /
- community structure /
- species diversity /
- Suichang /
- Wuxi River Basin
-
图 1 山蜡梅生境群落(A)和种群(B)径级结构分布
Figure 1 Community (A) and population (B) diameter class structure distribution of C. nitens
图 2 乔木层最优分类组数判断(A)及各层林木个体数(B)
Figure 2 Judgment index partitioning optimal groups (A) and the number of individuals at each level of the arbor layer (B)
表 1 乌溪江流域各样地基本信息
Table 1. Basic information of test plots in Wuxi River Basin
样地 北纬(N) 东经(E) 海拔/m 坡向 坡度/(°) 郁闭度 土壤 氮/(mg·kg−1) 磷/(mg·kg−1) 钾/(mg·kg−1) 有机质/(g·kg−1) pH P1 28°31′58″ 118°56′03″ 250 西 19 0.7 352.5±22.2 a 1.2±0.6 b 99.8±50.1 b 54.3±15.2 a 4.8 P2 28°32′10″ 118°55′44″ 271 西北 45 0.5 281.4±55.0 b 0.8±0.7 b 97.6±25.3 b 57.3±14.3 a 4.8 P3 28°30′57″ 118°58′38″ 260 东北 30 0.7 323.6±43.1 ab 1.5±1.1 b 138.6±35.5 b 55.3±8.4 a 4.9 P4 28°23′51″ 118°58′22″ 261 西南 25 0.6 368.2±45.9 a 3.1±0.4 a 251.7±45.7 a 64.9±14.8 a 4.8 P5 28°24′03″ 118°58′51″ 326 西南 50 0.6 292.3±32.8 b 0.7±0.6 b 105.1±23.0 b 44.6±8.4 a 5.0 说明:不同小写字母表示不同样地间差异显著(P<0.05)。  下载: 导出CSV
下载: 导出CSV
表 2 乌溪江山蜡梅群落种子植物属的分布区类型
Table 2. Areal-types of spermatophytic genera of C. nitens community in the Wuxi River Basin
分布类型 编号 分布区类型 属数/属 属的比例/% 小计/% 1 世界分布 3 4.23 4.23 热带分布 2 泛热带分布 15 21.13 50.70 3 热带亚洲和热带美洲间断分布 2 2.82 4 旧世界热带分布 6 8.45 5 热带亚洲至热带大洋州分布 2 2.82 6 热带亚洲至热带非洲分布 5 7.04 7 热带亚洲分布 6 8.45 温带分布 8 北温带分布 10 14.08 40.85 9 东亚和北美洲间断分布 12 16.90 10 旧世界温带分布 1 1.41 11 东亚分布 6 8.45 12 中国特有分布 3 4.23 4.23 合计 71 100.00 100.00
下载: 导出CSV
表 3 乌溪江山蜡梅生境群落的维管束植物统计
Table 3. Statistics of vascular plants in the C. nitens community in the Wuxi River Basin
科 属 种 类群 科 属 种 类群 豆科 Fabaceae 5 5 被子植物 虎耳草科 Saxifragaceae 1 1 被子植物 茜草科 Rubiaceae 5 5 被子植物 夹竹桃科 Apocynaceae 1 1 被子植物 樟科 Lauraceae 4 10 被子植物 金缕梅科 Hamamelidaceae 1 1 被子植物 蔷薇科 Rosaceae 4 7 被子植物 堇菜科 Violaceae 1 1 被子植物 壳斗科 Fagaceae 4 6 被子植物 菊科 Asteraceae 1 1 被子植物 禾本科 Poaceae 4 4 被子植物 鳞始蕨科 Lindsaeaceae 1 1 被子植物 大戟科 Euphorbiaceae 3 4 被子植物 马鞭草科 Verbenaceae 1 1 被子植物 报春花科 Primulaceae 3 3 被子植物 葡萄科 Vitaceae 1 1 被子植物 漆树科 Anacardiaceae 3 3 被子植物 山茱萸科 Cornaceae 1 1 被子植物 杜鹃花科 Ericaceae 2 4 被子植物 柿科 Ebenaceae 1 1 被子植物 山茶科 Theaceae 2 4 被子植物 鼠李科 Rhamnaceae 1 1 被子植物 安息香科 Styracaceae 2 2 被子植物 五加科 Araliaceae 1 1 被子植物 桑科 Moraceae 2 2 被子植物 蕈树科 Altingiaceae 1 1 被子植物 冬青科 Aquifoliaceae 1 3 被子植物 杨梅科 Myricaceae 1 1 被子植物 莎草科 Cyperaceae 1 3 被子植物 叶下珠科 Phyllanthaceae 1 1 被子植物 五列木科 Pentaphylacaceae 1 3 被子植物 柏科 Cupressaceae 2 2 裸子植物 蜡梅科 Calycanthaceae 1 2 被子植物 松科 Pinaceae 1 1 裸子植物 木犀科 Oleaceae 1 2 被子植物 里白科 Gleicheniaceae 2 2 蕨类植物 五福花科 Adoxaceae 1 2 被子植物 鳞毛蕨科 Dryopteridaceae 2 2 蕨类植物 菝葜科 Smilacaceae 1 1 被子植物 海金沙科 Lygodiaceae 1 1 蕨类植物 唇形科 Lamiaceae 1 1 被子植物 金星蕨科 Thelypteridaceae 1 1 蕨类植物 大麻科 Cannabaceae 1 1 被子植物 卷柏科 Selaginellaceae 1 1 蕨类植物 杜英科 Elaeocarpaceae 1 1 被子植物 乌毛蕨科 Blechnaceae 1 1 蕨类植物 胡桃科 Juglandaceae 1 1 被子植物
下载: 导出CSV
表 4 乌溪江山蜡梅生境群落乔木层主要物种的重要值
Table 4. Importance values of the main species in the arbor layer of C. nitens community in the Wuxi River Basin
树种 重要值/% P P1 P2 P3 P4 P5 杉木 Cunninghamia lanceolata 11.67 9.77 4.29 14.91 16.16 − 山蜡梅 Chimonanthus nitens 11.63 15.89 19.36 7.59 7.81 13.98 苦槠 Castanopsis sclerophylla 9.05 5.44 3.16 17.71 9.94 − 檵木 Loropetalum chinense 7.28 12.17 7.82 4.33 3.31 11.94 石栎 Lithocarpus glaber 7.08 − 3.26 8.61 14.07 2.90 青冈 Cyclobalanopsis glauca 5.41 − 20.32 6.52 − 14.43 峨眉鼠刺 Itea oblonga 4.97 3.01 4.01 5.66 7.05 3.79 黄瑞木 Adinandra millettii 4.04 4.92 − 3.74 6.94 − 水杉Metasequoia glyptostroboides 3.32 10.71 − − − − 木蜡树Toxicodendron sylvestre 2.93 3.62 11.68 2.30 − 3.72 马尾松Pinus massoniana 5.63 − − − − 牡荆Vitex negundo var. cannabifolia 3.84 − − − − 拟赤杨Alniphyllum fortunei 2.22 − 4.15 − − 格药柃Eurya muricata 2.12 − − − − 化香树 Platycarya strobilacea 2.08 − − − − 红楠Machilus thunbergii − − − 2.71 − 黄绒润楠Machilus grijsii − − − − 2.19 柳叶蜡梅Chimonanthus salicifolius − − − − 3.10 马银花Rhododendron ovatum − 5.92 − − 3.25 毛冬青Ilex pubescens − − − 2.07 − 毛花连蕊茶Camellia fraterna − − 2.00 3.07 2.75 密花树Rapanea neriifolia − − − − 17.05 山乌桕Triadica cochinchinensis − 7.60 − − − 水团花 Adina pilulifera − − − − 7.94 盐肤木Rhus chinensis − 7.94 − − − 迎春樱Cerasus discoidea − − − 2.02 − 硬斗石栎Lithocarpus hancei − − − − 2.57 中华杜英Elaeocarpus chinensis − 3.17 − − − 说明:48种重要值小于2的树种省略; −表示未分布。P为所有样地树种重要值综合排序;P1~P5为各样地树种重要值排序。
下载: 导出CSV
表 5 乌溪江山蜡梅灌木层主要物种重要值和多度
Table 5. Importance values and abundance of C. nitens community under growth layer in the Wuxi River Basin
树种 株数或丛数 盖度/% 频率 重要值/% 多度 杜茎山 Maesa japonica 29 74.0 0.2 10.17 Cop3 檵木 Loropetalum chinense 14 78.0 0.2 7.58 Soe 青冈 Cyclobalanopsis glauca 10 61.0 0.2 5.91 Cop3 峨眉鼠刺 Itea oblonga 6 43.0 0.2 4.19 Cop2 香花崖豆藤 Millettia dielsiana 8 32.0 0.2 3.97 Cop2 白花苦灯笼 Tarenna mollissima 7 33.4 0.2 3.86 Cop2 山蜡梅 Chimonanthus nitens 8 26.0 0.2 3.65 Cop2 水团花Adina pilulifera 5 30.0 0.2 3.30 Cop2 窄基红褐柃Eurya rubiginosa var. attenuata 6 20.0 0.2 2.95 Cop1 毛花连蕊茶Camellia fraterna 9 9.0 0.2 2.91 Cop1 黄瑞木Adinandra millettii 5 17.0 0.2 2.60 Cop1 绒毛润楠 Machilus velutina 2 26.0 0.2 2.52 Cop2 网脉酸藤子Embelia rudis 5 14.0 0.2 2.43 Cop1 小槐花Ohwia caudata 4 15.0 0.2 2.30 Cop1 映山红Rhododendron simsii 5 9.0 0.2 2.16 Cop1 说明:30种重要值小于2的树种省略。
下载: 导出CSV
表 6 乌溪江山蜡梅草本层主要物种重要值和多度
Table 6. Importance values and abundance of of C. nitens community herbal layer in the Wuxi River Basin
树种 株数或丛数 盖度/% 频率 重要值/% 多度 狗脊蕨 Woodwardia japonica 12 87.0 0.6 13.53 Soe 美丽复叶耳蕨 Arachniodes speciosa 14 66.0 0.4 11.38 Cop3 里白 Diplopterygium glaucum 11 77.0 0.4 11.21 Soe 芒萁 Dicranopteris pedata 14 41.0 0.4 9.69 Cop2 山类芦 Neyraudia montana 8 86.0 0.2 9.58 Soe 淡竹叶 Lophatherum gracile 20 31.0 0.2 9.49 Cop2 青绿薹草Carex breviculmis 5 27.0 0.4 6.01 Cop2 珠穗薹草Carex ischnostachya 4 12.0 0.4 4.69 Cop1 微糙三脉紫菀Aster ageratoides 3 10.0 0.2 2.92 Cop1 芒Miscanthus sinensis 2 12.0 0.2 2.75 Cop1 紫花堇菜Viola grypoceras 4 2.0 0.2 2.68 Sp 异穗卷柏Selaginella heterostachys 3 6.0 0.2 2.65 Cop1 三穗薹草Carex tristachya 1 14.0 0.2 2.58 Cop1 黑足鳞毛蕨Dryopteris fuscipes 2 7.0 0.2 2.41 Cop1 乌蕨Odontosoria chinensis 3 1.0 0.2 2.32 Sp 金星蕨Parathelypteris glanduligera 2 4.0 0.2 2.21 Sp 显子草Phaenosperma globosum 1 8.0 0.2 2.18 Cop1 海金沙Lygodium japonicum 1 1.0 0.2 1.70 Sp
下载: 导出CSV
表 7 乌溪江山蜡梅生境群落物种多样性指数
Table 7. Diversity indices of C. nitens community in the Wuxi River Basin
样地 乔木层 灌木层 草本层 N R H D J C N R H D J C N R H D J C P1 34 5.71 2.35 0.82 0.67 5.71 9 2.03 1.47 0.64 0.67 2.03 8 1.86 1.61 0.72 0.77 1.86 P2 16 2.99 1.97 0.80 0.71 2.99 4 0.93 1.09 0.60 0.79 0.93 4 1.04 1.26 0.69 0.91 1.04 P3 30 5.35 2.71 0.91 0.80 5.35 10 2.67 2.05 0.84 0.89 2.67 4 1.25 1.16 0.64 0.84 1.25 P4 31 5.27 2.67 0.90 0.78 5.10 15 3.66 2.50 0.90 0.92 3.66 4 1.00 1.38 0.75 0.99 1.00 P5 21 3.71 2.23 0.85 0.73 3.71 7 1.82 1.54 0.73 0.79 1.82 5 1.38 1.24 0.63 0.77 1.38 说明:N. 物种数(种);R. Margalef丰富度指数;H. Shannon-Wiener指数; D. Simpson指数; J. Pielou指数; C. 优势度指数。
下载: 导出CSV
-
[1] 吴艳秋, 刘宇灵, 林龙飞, 等. 山蜡梅叶化学成分、药理作用及临床应用研究进展[J]. 中华中医药杂志, 2021, 36(11): 6599 − 6607. WU Yanqiu, LIU Yuling, LIN Longfei, et al. Research progress on chemical composition, preparation, pharmacological action and clinical application of Chimonanthus nitens Folium [J]. China Journal of Traditional Chinese Medicine and Pharmacy, 2021, 36(11): 6599 − 6607. [2] 邱如文, 喻勋林, 蓝肖. 湖南双牌县打鼓坪林场亮叶腊梅群落研究[J]. 广西林业科学, 2008, 37(1): 8 − 12. QIU Ruwen, YU Xunlin, LAN Xiao. Study on the Chimonanthus nitens community in Daguping Forest Farm of Shuangpai County, Hunan Province [J]. Guangxi Forestry Science, 2008, 37(1): 8 − 12. [3] 方建新, 胡长玉, 程国栋. 黄山市蜡梅属植物资源的调查研究与利用[J]. 资源开发与市场, 2009, 25(5): 418 − 420. FANG Jianxin, HU Changyu, CHENG Guodong. Investigation and development of plant resources of Chimonanthus genus in Huangshan City [J]. Resource Development &Market, 2009, 25(5): 418 − 420. [4] 孙延军, 张冰, 林石狮, 等. 江西三清山优势种群钩锥-山蜡梅群落研究[J]. 广西植物, 2011, 31(1): 70 − 74. SUN Yanjun, ZHANG Bing, LIN Shishi, et al. Study on the dominant population Castanopsis tibetana-Chimonanthus nitens community in Mount Sanqingshan, China [J]. Guihaia, 2011, 31(1): 70 − 74. [5] 张小林, 张靖天, 迟春娟, 等. 乌溪江梯级水库的营养特征及水生态健康评价[J]. 环境工程技术学报, 2018, 8(5): 502 − 509. ZHANG Xiaolin, ZHANG Jingtian, CHI Chunjuan, et al. Nutritional characteristics and aquatic ecosystem health assessment in cascade reservoirs of Wuxijiang [J]. Journal of Environmental Engineering Technology, 2018, 8(5): 502 − 509. [6] 夏春平, 季新良, 彭辉, 等. 乌溪江源头水源涵养林空间结构二元分布特征[J]. 福建林业科技, 2021, 48(3): 10 − 13. XIA Chunping, JI Xinliang, PENG Hui, et al. Binary distribution spatial structure parameters of water conservation forest in the headstream area of Wuxijiang River [J]. Journal of Fujian Forestry Science and Technology, 2021, 48(3): 10 − 13. [7] 中国科学院中国植物志编辑委员会. 中国植物志: 第30卷第2册[M]. 北京: 科学出版社, 1979: 5 − 10. Flora of China Editorial Committee of Chinese Academy of Sciences. Flora of China: Vol 30 Issue 2 [M]. Beijing: Science Press, 1979: 5 − 10. [8] 吴征镒. 中国种子植物属的分布区类型[J]. 云南植物研究, 1991, 13(增刊4): 1 − 139. WU Zhengyi. The areal-types of chinese genera of seed plants[J]. Acta Botanica Yunnanica, 1991, 13(suppl 4): 1 − 139. [9] 吴征镒, 周浙昆, 李德铢, 等. 世界种子植物科的分布区类型系统[J]. 云南植物研究, 2003, 25(3): 245 − 257. WU Zhengyi, ZHOU Zhekun, LI Dezhu, et al. The areal-types of the world families of seed plants [J]. Acta Botanica Yunnanica, 2003, 25(3): 245 − 257. [10] 桂旭君, 练琚愉, 张入匀, 等. 鼎湖山南亚热带常绿阔叶林群落垂直结构及其物种多样性特征[J]. 生物多样性, 2019, 27(6): 619 − 629. GUI Xujun, LIAN Juyu, ZHANG Ruyun, et al. Vertical structure and its biodiversity in a subtropical evergreen broad-leaved forest at Dinghushan in Guangdong Province, China [J]. Biodiversity Science, 2019, 27(6): 619 − 629. [11] 王育松, 上官铁梁. 关于重要值计算方法的若干问题[J]. 山西大学学报(自然科学版), 2010, 33(2): 312 − 316. WANG Yusong, SHANGGUAN Tieliang. Discussion on calculating method of important values [J]. Journal of Shanxi University (Natural Science Edition), 2010, 33(2): 312 − 316. [12] 张金屯. 数量生态学[M]. 2版. 北京: 科学出版社, 2011: 248 − 249. ZHANG Jintun. Quantitative Ecology [M]. 2nd ed. Beijing: Science Press, 2011: 248 − 249. [13] 孙儒泳, 李庆芬, 牛翠娟, 等. 基础生态学[M]. 北京: 高等教育出版社, 2002: 140 − 143. SUN Ruyong, LI Qingfen, NIU Cuijuan, et al. Basic Ecology[M]. Beijing: Higher Education Press, 2002: 140 − 143. [14] 芦伟, 唐战胜, 郑振杰, 等. 浙江古田山濒危植物香果树群落组成和结构特征[J]. 生态环境学报, 2018, 27(6): 1052 − 1059. LU Wei, TANG Zhansheng, ZHENG Zhenjie, et al. The community composition and structural characteristics of Emmenopterys henryi forest, an endangered species in Gutianshan, Zhejiang Province [J]. Ecology and Environmental Sciences, 2018, 27(6): 1052 − 1059. [15] 杨泉光, 柴胜丰, 吴儒华, 等. 濒危植物东兴金花茶伴生群落及其种群结构特征[J]. 广西林业科学, 2020, 49(4): 492 − 497. YANG Quanguang, CHAI Shengfeng, WU Ruhua, et al. Characteristics of concomitant community and population structure of endangered plant Camellia tunghinensis [J]. Guangxi Forestry Science, 2020, 49(4): 492 − 497. [16] 汪殿蓓, 暨淑仪, 陈飞鹏. 植物群落物种多样性研究综述[J]. 生态学杂志, 2001, 20(4): 55 − 60. WANG Dianbei, JI Shuyi, CHEN Feipeng. A review on the species diversity of plant community [J]. Chinese Journal of Ecology, 2001, 20(4): 55 − 60. [17] 袁丛军, 刘娜, 谢涛, 等. 贵州青钱柳群落结构及物种多样性特征[J]. 江苏农业科学, 2022, 50(4): 131 − 138. YUAN Congjun, LIU Na, XIE Tao, et al. Community structure and species diversity characteristics of Cyclocarya paliurus in Guizhou Province [J]. Jiangsu Agricultural Sciences, 2022, 50(4): 131 − 138. [18] 叶万辉, 曹洪麟, 黄忠良, 等. 鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究[J]. 植物生态学报, 2008, 32(2): 274 − 286. YE Wanhui, CAO Hongling, HUANG Zhongliang, et al. Community structure of a 20 hm2 lower subtropical evergreen broadleaved forest plot in Dinghushan, China [J]. Journal of Plant Ecology, 2008, 32(2): 274 − 286. [19] 陈斌, 赵家豪, 关庆伟, 等. 江西武夷山中亚热带南方铁杉针阔混交林群落组成与结构[J]. 生态学报, 2018, 38(20): 7359 − 7372. CHEN Bin, ZHAO Jiahao, GUAN Qingwei, et al. Community composition and structure of a mid-subtropical coniferous (Tsuga chinensis var. tchekiangensis) and broadleaf mixed forest in Jiangxi Wuyishan, China [J]. Acta Ecologica Sinica, 2018, 38(20): 7359 − 7372. [20] 丁晖, 陈水飞, 徐辉, 等. 2013—2018年武夷山亚热带常绿阔叶林乔木层动态[J]. 生态学报, 2022, 42(8): 3458 − 3469. DING Hui, CHEN Shuifei, XU Hui, et al. Dynamics of arbor layer in the subtropical evergreen broad-leaved forest in the Wuyi Mountains, Fujian Province, southeastern China in 2013−2018 [J]. Acta Ecologica Sinica, 2022, 42(8): 3458 − 3469. [21] 祝燕, 赵谷风, 张俪文, 等. 古田山中亚热带常绿阔叶林动态监测样地—群落组成与结构[J]. 植物生态学报, 2008, 32(2): 262 − 273. ZHU Yan, ZHAO Gufeng, ZHANG Liwen, et al. Community composition and structure of Gutianshan forest dynamic plot in a mid-subtropical evergreen broad-leaved forest, east China [J]. Journal of Plant Ecology, 2008, 32(2): 262 − 273. [22] CHEN Xiongwen, NIU Jianzhi. Relationships between tree height and tree species richness at small scales[J/OL]. Acta Oecologica, 2020, 109: 103668[2022-08-09]. doi: 10.1016/j.actao.2020.103668. [23] 郑超超, 伊力塔, 余树全, 等. 浙江江山公益林群落生物多样性与稳定性[J]. 东北林业大学学报, 2013, 41(11): 31 − 35. ZHENG Chaochao, YI Lita, YU Shuquan, et al. Biodiversity and stability of ecological service forest of Jiangshan City in Zhejiang Province [J]. Journal of Northeast Forestry University, 2013, 41(11): 31 − 35. [24] 庞圣江, 张培, 杨保国, 等. 广西大青山西南桦人工林林下植物多样性与稳定性[J]. 中南林业科技大学学报, 2018, 38(2): 103 − 107, 113. PANG Shengjiang, ZHANG Pei, YANG Baoguo, et al. Species diversity and community stability of Betula alnoides plantations in Daqingshan Mountain in areas of Guangxi [J]. Journal of Central South University of Forestry &Technology, 2018, 38(2): 103 − 107, 113. [25] 巫翠华, 蔡建军, 李华, 等. 紫柏山壳斗科植物群落物种多样性和稳定性研究[J]. 中南林业科技大学学报, 2021, 41(8): 108 − 115. WU Cuihua, CAI Jianjun, LI Hua, et al. Studies on species diversity and stability of the fagaceae community in the Zibai Mountain [J]. Journal of Central South University of Forestry &Technology, 2021, 41(8): 108 − 115. [26] 朱杰, 吴安驰, 邹顺, 等. 南亚热带常绿阔叶林树木多样性与生物量和生产力的关联及其影响因素[J]. 生物多样性, 2021, 29(11): 1435 − 1446. ZHU Jie, WU Anchi, ZOU Shun, et al. Relationships between tree diversity and biomass/productivity and their influence factors in a lower subtropical evergreen broad-leaved forest [J]. Biodiversity Science, 2021, 29(11): 1435 − 1446. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220570
 点击查看大图
点击查看大图
计量
- 文章访问数: 1854
- HTML全文浏览量: 434
- PDF下载量: 39
- 被引次数: 0
