





-
生物多样性及保护的研究是倍受关注的生态学问题[1]。森林作为陆地最重要的生态系统类型,其生物多样性一直是生态学研究的热点[2]。林下草本植物作为森林的重要组成部分,在维持森林生态系统功能方面起着重要的作用[3-4]。不同类型的森林群落具有不同的物种多样性,如阔叶林、泰加林、热带雨林及季节性干旱森林等均表现了物种多样性的差异[5-8]。林分密度对林下草本植物多样性有明显影响[9];也有研究表明,林分密度对草本植物多样性影响不明显[10]。林分密度对林下草本植物多样性的影响还需进一步研究。山西省关帝山庞泉沟自然保护区是华北地区天然次生林重要的分布区,其中云杉Picea是该地区主要树种,以云杉为主的森林群落对该地区生态系统的维持起着重要的作用。曾有文献采用方差均值比,Morisita指数Iq,负二项参数K等测定了寒温性针叶林物种的分布格局[11];采用数量分析的方法研究了关帝山庞泉沟主要森林群落华北落叶松林和云杉林及其交错区的植物多样性[12];采用Rao指数和群落加权平均数(CWM)指数计算群落的功能多样性指数[13]。本研究以华北山区典型的云杉林为研究对象,选择不同密度的云杉林分,调查林下草本植物物种组成,分析林下草本植物多样性,旨在了解在以云杉为代表的寒温性针叶林中,林分密度对林下草本植物多样性是否有明显的影响,为深入了解云杉林森林生态系统生物多样性提供基础,同时为云杉林可持续经营管理提供参考。
-
研究地在山西省庞泉沟国家级自然保护区,地处吕梁山脉中段,位于山西省交城县西北部和方山县东北部,37°45′~37°55′N,111°22′~111°33′E,海拔为1 600~2 831 m。该区南北长15.0 km,东西长14.5 km,总面积为10 443.5 hm2。研究地气候属于暖温带大陆性季风气候,1月平均气温为-10.2 ℃,7月平均气温为17.5 ℃,年平均气温为4.2 ℃;年平均降水量822.6 mm,全年降水量极不均匀,多集中于7-8月,占全年降水量的75%以上;年平均蒸发量1 268.0 mm,相对湿度71%。土壤为山地棕壤,平均土层厚度70 cm,腐殖质层厚8 cm[12]。
庞泉沟自然保护区植被茂盛,植被覆盖率高达95%。云杉(包括青扦Picea wisonii和白扦Picea meyeri)和华北落叶松Larix principis-rupprechtii为主要的建群种。此外还有油松Pinus tabuliformis,山杨Populus davidiana,白桦Betula platyphylla,红桦Betula albo-sinensis,辽东栎Quercus wutaishannica等。灌木主要有沙棘Hippophae rhamnoides,土庄绣线菊Spiraea pubescens,忍冬Lonicera japonica,灰栒子Cotoneaster acutifolius等。草本植物有舞鹤草Maianthemum bifolium,小红菊Dendranthem achaneti,唐松草Thalictrum aquilegifolium,细叶薹草Carex stenophylla和茜草Rubia cordifolia等。
-
在研究区选择不同密度的云杉林,各密度的云杉林设3个样地,样地的面积为20 m×20 m,设置1 m×1 m的草本植物调查样方9个·样地-1,共设81个草本植物的调查样方。样地林分基本特征见表 1。
-
在2013年6月对每个样地进行调查。对样地内乔木进行每木检尺,测定其胸径、树高、枝下高、郁闭度(表 1);对样地内的草本样方进行调查,记录草本样方内出现的所有草的种类、株数(丛数)、盖度、高度等。
表 1 不同密度林分的基本特征
Table 1. Basic characteristics of stands in different densities
样地 密度/(株·hm-2) 海拔/m 坡向(°) 树高/m 胸径/cm 枝下高/m 郁闭度 1 475 1 900~1 950 NE 23.8 36.2 4.8 0.65 2 500 1 900~1 950 NE 25.8 36.8 6.7 0.70 3 525 1 900~1 950 NE 23.9 29 4.9 0.70 4 675 1 800~1 850 NW 21.1 24.4 7.1 0.85 5 700 1 800~1 850 NW 20.4 24.4 5.7 0.83 6 800 1 800~1 850 NW 21.3 23.8 6 0.87 7 900 1 800~1 850 NW 20.4 23 7.6 0.90 8 925 1 800~1 850 NW 21.8 26.3 7.8 0.89 9 1 000 1 800~1 850 NW 23.2 23.6 9.7 0.91 -
本研究的物种多样性采用多样性指数、物种丰富度、均匀度指数等来表征。Simpson多样性指数(D):$D=1 - \sum\limits_{i=1}^s {P_i^2} $。Shannon-Wiener多样性指数(H):$H=- \sum\limits_{i=1}^s {{P_i}} \ln {p_i}$。Patrick丰富度(R):R=S。Pielou均匀度指数(Js):${J_{\rm{s}}}=\frac{H}{{\ln S}}$。其中:Pi=ni/N, ni为样方内某物种的个体数,N为样方内所有物种的个体数;S为样地内物种数。
-
在研究区的云杉林中共调查到37种草本植物,隶属于18个科35个属。既有一些百合科Liliaceae,菊科Compositae和毛茛科Ranunculaceae等世界性大科,又有一些少种科。其中百合科和菊科的物种数最多,都为6种,占草本植物种的16.2%,有10个属只有1个物种,其属、种分别占样地的28.6%和27.0%。其详细信息见表 2。
表 2 云杉林下草本植物物种组成
Table 2. Composition of herbs in spruce forest

-
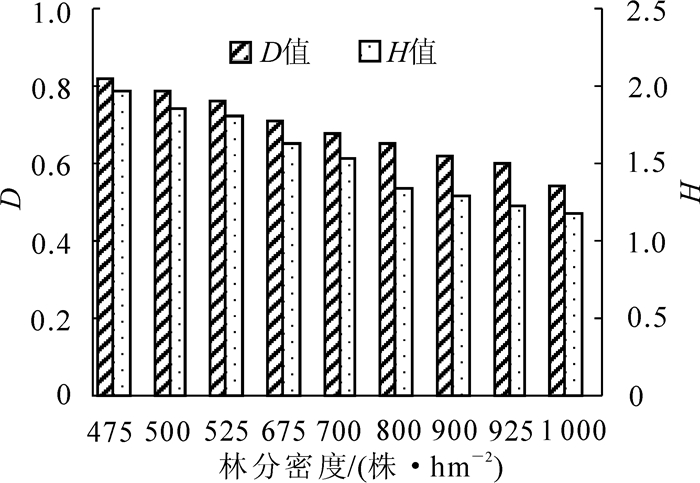
物种多样性是群落结构的重要指标,通过多样性指数可以间接反映出群落功能的特征。本研究选用Simpson指数(D)和Shannon-Wiener指数(H)进行物种多样性测度。由图 1可知:草本植物的Simpson指数和Shannon-Wiener指数随着林分密度的增加而减小。当林分密度为475株·hm-2时,多样性指数(D和H)分别为0.82和1.97;当林分密度为1 000株·hm-2时,多样性指数(D和H)分别为0.54和1.18。

图 1 不同林分密度云杉林下草本植物的Simpson(D值)和Shannon-Wiener(H值)
Figure 1. Simpson index and Shannon-Wiener index of herbs in spruce forest of different densities
-
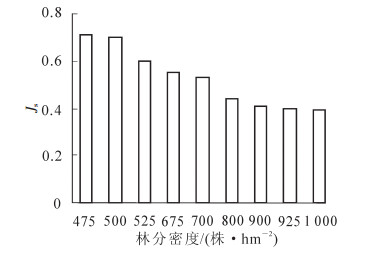
Pielou均匀度指数(Js)表示群落均匀度的指标,它可以反映群落中个体数量分布的均匀程度。由图 2可知,随着林分密度的增加,云杉林下草本植物的均匀度指数逐渐减小。当林分密度为475株·hm-2时,均匀度指数为0.71;当林分密度为1 000株·hm-2时,均匀度指数为0.39。

图 2 不同林分密度云杉林下草本植物的均匀度指数(Js值)
Figure 2. Pielou index of herbs in spruce forest of different densities
-
丰富度是指群落中物种数目的多少。一般情况,当物种数目越多,丰富度就越大;相反,物种数目越少,丰富度越小。本研究中物种的丰富度与林分密度之间没有一定的变化规律(图 3)。

图 3 不同林分密度下草本植物的丰富度(S)
Figure 3. Patrick index of herbs in pruce forest of different densities
-
林分密度与草本植物多样性指数的相关性分析表明(表 3):林分密度与Shannon-Wiener多样性指数(H),Pielou均匀度指数(Js)和Simpson多样性指数(D)呈极显著的负相关,其中Simpson多样性指数(D)受林分密度的影响最大;Shannon-Wiener多样性指数(H)与Pielou均匀度指数(Js)和Simpson多样性指数(D)呈极显著的正相关;Pielou均匀度指数(Js)和Simpson多样性指数(D)呈极显著正相关;物种丰富度(S)与Shannon-Wiener多样性指数(H),Pielou均匀度指数(Js)和Simpson多样性指数(D)都呈负相关,而与林分密度呈显著正相关。
表 3 林分密度与草本植物多样性指数的相关系数
Table 3. Correlation coefficients between stand density and diversity index of herbaceous plants
林分密度 H Js D S 林分密度 1 -0.9875** -0.9697** -0.9886** -0.7503* H 1 0.9835** 0.9835** -0.7630* Js 1 0.9683** -0.8608** D 1 -0.7283* S 1 *表示在0.05水平下显著,**表示在0.01水平下极显著。 -
在本研究区,不同密度云杉林下草本植物的多样性存在明显差异。随着林分密度的增加,草本植物的Simpson多样性指数(D值)、Shannon-wiener多样性指数(H值)和均匀度指数(Js)都逐渐减小。此外,林分密度与草本植物多样性指数都具有一定的相关性。
已有研究表明[16-17],随着林分密度的增加,不同密度林下草本植物的多样性指数(Simpson多样性指数和Shannon-Wiener指数)逐渐减小,这与本研究结果一致。但是,李民义等[9]和吕婧娴等[10]对不同密度林下草本植物多样性的研究表明:随着林分密度的增加,草本植物多样性指数表现为先增大后减小的趋势。出现上述不同结果的原因可能是随着林分密度的增加,林下微环境(如光照、温度、水分等)受到较复杂的影响,从而引起草本植物多样性指数出现上述复杂的变化。一些学者研究表明:随着林分密度的增加,不同密度林下草本植物的均匀度指数逐渐减小[15-17],这与本研究结果一致。可能随着林分密度的增加,林隙减少,从而导致均匀度指数减小。吕婧娴等[10]对不同密度林下草本植物多样性的研究表明:随着林分密度的增加,物种丰富度没有一定的变化规律,这与本研究结果一致。其可能原因是物种丰富度指数受多种因素(如海拔、坡向、土壤条件等)的共同作用。Risser等[18]认为只有Shannon-Wiener指数与林分密度呈负相关,其他物种多样指数的变化并无一定的规律。本研究的云杉林为寒温性常绿针叶林,为亚高山暗针叶林的一个组成部分,枝冠稠密常绿,林木自然整枝不良,林下潮湿阴暗,灌木稀少,盖度小于20%。尤其在密度大的林分中,林下光照不足,草本植物种类和数量较少,草本植物多样性指数较小,均匀度指数也相对小。在密度相对小的林分,光照条件相对好,草本植物多样性指数和均匀度指数相对高。本研究中不同密度的林分,丰富度没有规律性变化,这表明林下草本植物个体数量和物种数不仅受光照的影响,而是多种因素综合作用的结果。此外,林木密度对林下草本植物个体数量和物种数的影响作用不能明显地表现出来。
草本植物多样性受多种因素的影响,例如地形、光照、林冠层的分布、土壤肥力等。曹云生等[21]对不同的森林群落的地形因子进行典范对应分析(CCA)排序,认为坡向是影响草本植物多样性差异的最主要地形制约因子,海拔与草本植物多样性指数、均匀度指数和丰富度指数呈现正相关。有研究指出,林下总光照和林冠开阔度对草本植物的影响最大,并且林隙光照对林下植物的分布有明显的指示作用,大林隙内草本植物的丰富度明显高于小林隙的[20]。林冠层的分布又影响光照环境和净降水量的分布,从而影响草本植物的多样性[21]。白永飞等[22]对锡林河流域和沙坡头人工植被植物群落的研究表明:物种丰富度和多样性指数与土壤有机碳及全氮含量呈正相关。文海燕等[23]对草本植物的多样性研究表明:草本植物物种多样性与土壤有机碳、全氮、全磷和有效氮呈正相关。草本植物多样性受多种环境因子的制约,各种环境因子又相互作用共同影响植物多样性。要全面了解不同密度云杉林下草本植物的多样性。需将草本植物的生理生态学特征与林分微环境利用状况以及群落的种间关系结合起来做进一步研究。
草本植物是森林生态系统生物多样性的重要组成成分。促进林下草本植物的良好发育,必然大大提高森林生态系统的物种多样性。在森林抚育管理中,不仅要考虑乔木的保留密度,同时要兼顾林下植物的多样性,通过合理调控林分密度,适当提高林下草本植物多样性,可增加林分的稳定性。
Diversity of understory herbs in Picea forests with different stand densities
-
摘要: 为了解寒温性针叶林林分密度与林下草本植物多样性的关联性, 在山西省庞泉沟国家级自然保护区, 选择以云杉Picea林为代表的寒温性针叶林, 在不同密度的林分中设置样地, 调查林下草本植物的物种组成, 分析林下草本植物的多样性, 并对林分密度和多样性指数进行相关分析。结果表明:①在研究区的云杉林中共调查到37种草本植物, 隶属于18个科35个属。②云杉林下草本植物多样性指数(Simpson指数、Shannon-Wiener指数和Pielou均匀度指数)随着林分密度的增加而减小。当林分密度为475株·hm-2时, Simpson指数、Shannon-Wiener多样性指数和Pielou均匀度指数分别为0.82, 1.97和0.71;当林分密度为1 000株·hm-2时, Simpson指数、Shannon-Wiener多样性指数和Pielou均匀度指数分别为0.54, 1.18和0.39。③不同密度的云杉林中, 草本植物丰富度指数没有明显的变化规律。研究认为:在以云杉为主要树种的寒温性针叶林中, 林分密度对林下草本植物的多样性指数有明显的影响, 随着林分密度的增加多样性指数减小。Abstract: To explain the relevance of stand density and understory vegetation diversity in cold, temperate coniferous forests, the Picea (spruce) forests in the Pangquangou Nature Reserve of Guandi Mountain, Shanxi Province were selected as a representative study. In the experiment, twenty seven sample plots were set in stands with nine stand densities (475, 500, 525, 675, 700, 800, 900, 925 and 1 000 trees·hm-2) and three replicates. The sample plot area is 400 m2 (20 m×20 m). Species composition of understory herbaceous plants was determined in all replicates and the relevance between stand density and diversity index was analyzed using Simpson, Shannon-Wiener Diversity, Pielou Evenness Indexes and Richness. Results showed (1) 37 species of herbs belonging to 35 genera in 18 families.(2) The diversity indexes of herbaceous plants decreased with an increase in stand density:with a stand density of 475 trees·hm-2 the Simpson Index was 0.82, the Shannon-Wiener Index was 1.97, and the Pielou Index was 0.71; whereas, at 1 000 trees·hm-2 the Simpson Index was 0.54, the Shannon-Wiener Index was 1.18, and the Pielou Index was 0.39.(3) With different stand densities, the Richness of herbaceous plants ranged from 14 to 23. Thus, in cold, temperate coniferous forests with Picea as the main species, the diversity index of understory herbs decreased with a stand density increase.
-
Key words:
- forest ecology /
- Picea forests /
- stand density /
- plant composition /
- diversity of herbaceous plants
-
图 1 不同林分密度云杉林下草本植物的Simpson(D值)和Shannon-Wiener(H值)
Figure 1 Simpson index and Shannon-Wiener index of herbs in spruce forest of different densities
图 2 不同林分密度云杉林下草本植物的均匀度指数(Js值)
Figure 2 Pielou index of herbs in spruce forest of different densities
图 3 不同林分密度下草本植物的丰富度(S)
Figure 3 Patrick index of herbs in pruce forest of different densities
表 1 不同密度林分的基本特征
Table 1. Basic characteristics of stands in different densities
样地 密度/(株·hm-2) 海拔/m 坡向(°) 树高/m 胸径/cm 枝下高/m 郁闭度 1 475 1 900~1 950 NE 23.8 36.2 4.8 0.65 2 500 1 900~1 950 NE 25.8 36.8 6.7 0.70 3 525 1 900~1 950 NE 23.9 29 4.9 0.70 4 675 1 800~1 850 NW 21.1 24.4 7.1 0.85 5 700 1 800~1 850 NW 20.4 24.4 5.7 0.83 6 800 1 800~1 850 NW 21.3 23.8 6 0.87 7 900 1 800~1 850 NW 20.4 23 7.6 0.90 8 925 1 800~1 850 NW 21.8 26.3 7.8 0.89 9 1 000 1 800~1 850 NW 23.2 23.6 9.7 0.91  下载: 导出CSV
下载: 导出CSV
表 2 云杉林下草本植物物种组成
Table 2. Composition of herbs in spruce forest
表 3 林分密度与草本植物多样性指数的相关系数
Table 3. Correlation coefficients between stand density and diversity index of herbaceous plants
林分密度 H Js D S 林分密度 1 -0.9875** -0.9697** -0.9886** -0.7503* H 1 0.9835** 0.9835** -0.7630* Js 1 0.9683** -0.8608** D 1 -0.7283* S 1 *表示在0.05水平下显著,**表示在0.01水平下极显著。
下载: 导出CSV
-
[1] 岳天祥.生物多样性研究及其问题[J].生态学报, 2001, 21(3):462-467. YUE Tianxiang. Studies and questions of biological diversity[J]. Acta Ecol Sin, 2001, 21(3):462-467. [2] 李步杭, 张健, 姚晓琳, 等.长白山阔叶红松林草本植物多样性季节动态及空间分布格局[J].应用生态学报, 2008, 19(3):467-473. LI Buhang, ZHANG Jian, YAO Xiaolin, et al. Seasonal dynamics and spatial distribution patterns of herbs diversity in broadleaved Korean pine (Pinus koraiensis) mixed forest in Changbai Mountains[J]. Chin J Appl Ecol, 2008, 19(3):467-473. [3] 巍强, 张秋良, 代海燕, 等.大青山不同林地类型土壤特性及其水源涵养功能[J].水土保持学报, 2008, 22(2):111-115. WEI Qiang, ZHANG Qiuliang, DAI Haiyan, et al. Soil characteristics and water conservation of different forest types in Daqing Mountain[J]. J Soil Water Conserv, 2008, 22(2):111-115. [4] 张佳, 李生宇, 靳正忠, 等.防护林下草本植物层片物种多样性与环境因子的关系[J].干旱区研究, 2011, 28(1):118-125. ZHANG Jia, LI Shengyu, JIN Zhengzhong, et al. Relationship between species diversity of herbaceous plants in the shelterbelt and environment factors[J]. Arid Zone Res, 2011, 28(1):118-125. [5] 钱迎倩, 马克平.生物多样性研究理论与方法[M].北京:中国科学技术出版社, 1994:36-54. [6] 郝占庆, 于德永, 杨晓明, 等.长白山北坡植物群落α多样性及其随海拔梯度的变化[J].应用生态学报, 2002, 13(7):785-789. HAO Zhanqing, YU Deyong, YANG Xiaoming, et al. α diversity of communities and their variety along altitude gradient on northern slope of Changbai Mountain[J]. Chin J Appl Ecol, 2002, 13(7):785-789. [7] 马克平, 黄建辉, 于顺利, 等.北京东灵山地区植物群落多样性的研究(Ⅱ)丰富度、均匀度和物种多样性指数[J].生态学报, 1995, 15(3):268-277. MA Keping, HUANG Jianhui, YU Shunli, et al. Plant community diversity in Dongling mountain, Beijing, China (Ⅱ) species richness, evenness and species diversities[J]. Acta Ecol Sin, 1995, 15(3):268-277. [8] 臧润国, 杨彦承, 蒋有绪.海南岛霸王岭热带山地雨林群落结构及树种多样性特征的研究[J].植物生态学报, 2001, 25(3):270-275. ZHANG Runguo, YANG Yancheng, JIANG Youxu. Community structure and tree species diversity characteristics in a tropical mountain rain forest in Bawangling nature reserve, Hainnan Island[J]. Acta Phytoecol Sin, 2001, 25(3):270-275. [9] 李民义, 张建军, 郭宝妮, 等.晋西黄土区不同密度油松人工林林下植物多样性及水文效应[J].生态学杂志, 2013, 32(5):1083-1090. LI Minyi, ZHANG Jianjun, GUO Baoni, et al. Understory plant species diversity and hydrological effect of Pinus tabulaeformis plantations with different stand densities in Loess Plateau of Western Shanxi, China[J]. Chin J Ecol, 2013, 32(5):1083-1090. [10] 吕婧娴, 王得祥, 张宋智, 等.小陇山林区不同密度油松人工林林下物种多样性研究[J].西北农林科技大学学报:自然科学版, 2010, 38(11):49-56. LÜ Jingxian, WANG Dexiang, ZHANG Songzhi, et al. Study on the species diversity of undergrowth in the different stand densities of Pinus tabulaeformis plantation of Xiaolongshan area[J]. J Northwest A & F Univ Nat Sci Ed, 2010, 38(11):49-56. [11] 石晓东, 韩有志.庞泉沟自然保护区寒温性针叶林的种群结构与分布格局[J].武汉植物学研究, 2008, 26(5):489-494. SHI Xiaodong, HAN Youzhi. Population structure and distribution pattern of cold temperate coniferous forest in the Pangquangou National Nature Reserve[J]. J Wuhan Bot Res, 2008, 26(5):489-494. [12] 李毳.山西庞泉沟针叶林群落植物多样性研究[J].水土保持学报, 2001, 15(5):95-107. LI Cui. Study on plant diversity of conifer forest communities in Pangquangou[J]. J Soil & Water Conserv, 2001, 15(5):95-107. [13] 郭玉永.山西庞泉沟自然保护区典型森林群落功能多样性研究[J].山西林业科技, 2012, 41(2):16-18. GUO Yuyong. Research on functional diversity of typical forest communities in Pangquangou natural reserve in Shanxi[J]. J Shanxi For Sci Technol, 2012, 41(2):16-18. [14] 陶福禄, 李树人, 冯宗炜, 等.豫西山区日本落叶松林下植物物种多样性的研究[J].生态学杂志, 1998, 17(4):1-6. TAO Fulu, LI Shuren, FENG Zongwei, et al. Studies on plant species diversity under the canopy of Larix kaempferi community in west mountain area of Henan Province[J]. Chin J Ecol, 1998, 17(4):1-6. [15] 李俊清.森林生态学[M].北京:高等教育出版社, 2006:299-304. [16] 鲁绍伟, 刘凤芹, 余新晓, 等.北京山区不同密度油松结构与功能研究[J].水土保持研究, 2008, 15(1):117-121. LU Shaowei, LIU Fengqin, YU Xinxiao, et al. Studies on the configuration and function of different density pine in rocky mountain area of Beijing[J]. Res Soil & Water Conserv, 2008, 15(1):117-121. [17] 侯磊, 张硕新, 陈云明, 等.林分密度对人工油松林下植物的影响[J].西北林学院学报, 2013, 28(3):46-52. HOU Lei, ZHANG Shuoxin, CHEN Yunming, et al. Impact of stand density on the vegetation under artificial Pinus tabulaeformis[J]. J Northwest For Univ, 2013, 28(3):46-52. [18] RISSER P G, RICE E L. Diversity in tree species in Oklahoma upland forest[J]. Ecology, 1971, 52(5):876-880. [19] 曹云生, 杨新兵, 张伟, 等.冀北山区森林群落草本多样性及其与地形关系研究[J].生态环境学报, 2010, 19(12):2840-2844. CAO Yunsheng, YANG Xinbing, ZHANG Wei, et al. Herbaceous diversity and the relationship with terrain in forest communities in northern Hebei mountain[J]. Ecol Environ Sci, 2010, 19(12):2840-2844. [20] 段文标, 王丽霞, 陈立新, 等.红松阔叶混交林林隙大小及光照对草本植物的影响[J].应用生态学报, 2013, 24(3):614-620. DUAN Wenbiao, WANG Lixia, CHEN Lixin, et al. Effects of forest gap size and light intensity on herbaceous plants in Pinus koraiensis-dominated broadleaved mixed forest[J]. Chin J Appl Ecol, 2013, 24(3):614-620. [21] 夏富才, 潘春芳, 赵秀海, 等.长白山原始阔叶红松林林下草本植物多样性格局及其影响因素[J].西北植物学报, 2012, 32(2):370-376. XIA Fucai, PAN Chunfang, ZHAO Xiuhai, et al. Influence of overstory on seasonal variability of understory herbs in primary broad-leaved Korean pine forest of Changbai mountain[J]. Acta Bot Boreali-Occident Sin, 2012, 32(2):370-376. [22] 白永飞, 李陵浩, 王其兵, 等.锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究[J].植物生态学报, 2000, 24(6):667-673. BAI Yongfei, LI Linghao, WANG Qibing, et al. Changes in plant species diversity and productivity along gradients of precipitation and elevation in the Xilin River Basin, Inner Mongolia[J]. Acta Phytoecol Sin, 2000, 24(6):667-673. [23] 文海燕, 傅华, 赵哈林.退化沙质草地植物群落物种多样性与土壤肥力的关系[J].草业科学, 2008, 25(10):6-9. WEN Haiyan, FU Hua, ZHAO Halin. The relationship between plant species biodiversity and soil fertility in degraded sandy grassland[J]. Pratac Sci, 2008, 25(10):6-9. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2014.05.003
 点击查看大图
点击查看大图
计量
- 文章访问数: 4356
- HTML全文浏览量: 850
- PDF下载量: 567
- 被引次数: 0
