





-
重组竹的加工利用技术已经十分成熟,眼下已经能够大规模批量化生产,并且广泛应用于建筑结构材、家具制造以及装饰装修领域。中国的竹资源十分丰富,而重组竹的竹材资源利用率高达90.0%,因此,重组竹的研发利用在保护木材资源,缓解中国木材资源供需紧张的局面上有着不可磨灭的贡献[1-3]。重组竹新产品的开发利用进展较差,影响了重组竹产业的发展。目前,已经有部分专家学者开始探索重组竹刨切单板的开发,在前期刨切竹单板的研究的基础上[4-6]提出开发纹理细腻、质量较好的重组竹刨切单板的产品,带动重组竹加工产业的发展。由于重组竹加工工艺的特殊性,重组竹密度达到了1.1~1.2 g·cm-3,含水率低,硬度大,而且结构十分致密[7-9],因而难以直接进行刨切。因此,如何提高重组竹方材含水率以达到软化刨切就成为重组竹刨切单板加工的关键难题。本研究以重组竹竹方为研究对象,采用加压浸注和水煮软化的生产工艺,增大重组竹方材的含水率与软化程度,并研究竹方材的导热情况,解决重组竹单板刨切过程的关键难题,为企业生产提供技术支持。
-
重组竹方材来自于浙江绍兴竹木制品有限公司,采用6~8年生毛竹Phyllostachys edulis制成去青去黄后的竹片,进行疏解制成精细竹束,经炭化、干燥、浸胶、干燥、冷压热固化等工序制备而成,其规格为2 550 mm × 140 mm × 140 mm,竹方密度为1.1 g·cm-3左右,初始含水率为6.0%~7.0%。
加压浸注试验采用DN800-2500型加压浸渍罐进行;高温水煮采用自制蒸汽加热水槽。所有试件都采用磅秤进行质量的称量。
-
重组竹方材放入加压罐后,抽真空处理,压力为0.8 MPa,时间为30 min。处理完后在不同的压力和时间下进行加压浸注,分别采用加压压力为0.3,0.6,0.9,1.2,1.5 MPa以及保压时间为2.0,4.0,6.0,8.0,10.0 h。以重组竹方材的增重率为评价指标。重组竹方材的增重率按下式计算:RW=(m1-m0)/m0×100%,其中:RW(%)为试件的增量率,m1(g)为试件处理后的质量,m0(g)为试件处理前的质量。
-
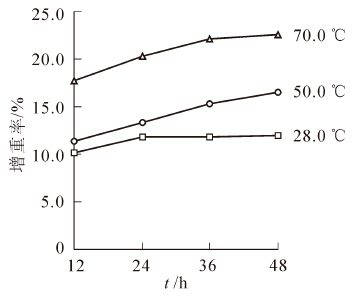
水煮软化试验在不同的温度和水煮时间下进行,分别采用温度为常温28.0,50.0, 70.0 ℃以及水煮时间为12.0,24.0,36.0,48.0 h的水煮工艺,将重组竹方材放入水煮池中后让水淹没方料表面10 cm以上,然后进行升温,升温速率为1.0~1.5 ℃·h-1,升温至(70.0 ± 5.0) ℃进行水煮,以水煮后重组竹方材的增重率为评价指标。
-
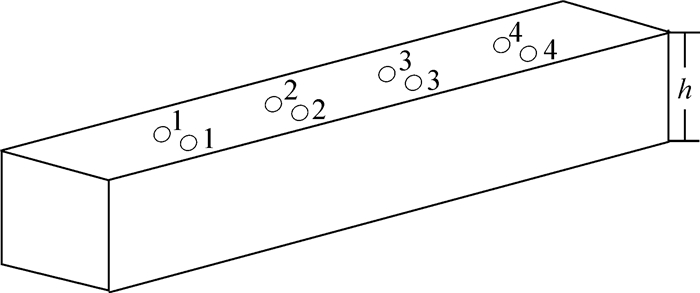
将含水率分别为10.0%和20.0%的重组竹方材按图 1所示位置进行打孔处理。将试样分别放到水温为70.0 ℃的恒温水槽中,让方料表面露出水面5 mm左右,然后对孔内温度用温度测定仪隔0.5 h测定1次,以最终各孔达到稳定不再变化温度值的为指标。

图 1 重组竹导热测试的打孔位置
Figure 1. Punch position of thermal conductivity test of recombinant bamboo
-
竹材硬度试件的制作参照国家标准GB/T 1941-2009《木材硬度试验方法》。试件尺寸为(长×宽×厚)70 mm × 50 mm × 50 mm,其长轴应与竹材纹理相平行。具体方法如下:采用硬度计测试实验设备,将试样放于硬度试验设备支座上,并使试验设备的钢半球直径为(5.64 ± 0.01) mm端部正对竹片试验面中心位置,以3~6 mm·min-1的速度将钢压头压入试样的试验面,直至压入2 mm深。将载荷读数计下,准确至10 N。
-
试件经软化处理前后,肉眼观察记录试件表面的裂纹数、裂纹长度,显微镜观察裂纹宽度,得到平均裂纹长度和平均裂纹宽度[16]。
-
图 2所示为重组竹方材在不同压力和时间下重组竹方材的增重率随时间的变化。由图 2所示,随着加压浸注压力的增大试件的增重率随着时间的延长呈逐渐增大的趋势,这是因为压力的增加使得试件的内外压力差变大,使得水分更加容易浸注到试件内部[10],但如图 2所示:在压力为1.2 MPa和1.5 MPa时试件的最终增重率趋于一致,压力大于1.2 MPa后增大趋势减缓。随着加压浸注时间的延长,增重率呈逐渐增大的趋势,但在8.0 h以后增大趋势逐渐减缓。在实际生产中,并不是压力和浸注时间越大越好,一方面要考虑设备的受压能力,另一方面也要考虑成本,所以在综合考虑的情况下,应选择1.2 MPa,8.0 h为最佳的浸注条件。

图 2 压力对重组竹方材增重率的影响
Figure 2. Influence of pressrue on weight gain rate of recombinant bamboo
图 3 所示的是重组竹方材在不同温度和时间水煮后试件增重率的变化情况。从图 3上可知:常温28.0 ℃状态下,重组竹的增重率变化不大,在50.0 ℃温度水煮下,增重率随着水煮时间的延长逐步升高,在36.0 h以后这种升高趋势也逐渐变小。同样在70.0 ℃条件下进行水煮时,36.0 h前试件增重率增加显著,36.0 h以后随着时间的延长,增重率逐步上升,但是上升趋势逐步变缓,所以36.0 h为最佳水煮软化时间。可以看出重组竹方材由于其制造工艺的特殊性,其结构十分紧凑,导致水分的渗透十分困难,因此,在进行重组竹方料的刨切等工艺时需要采用本研究的加压浸注和水煮软化工艺进行增湿软化处理,提高重组竹方材的增重率及温度,达到一定程度上的软化。

图 3 蒸煮温度对重组竹方材增重率的影响
Figure 3. Influence of temperature on weight gain rate of recombinant bamboo
-
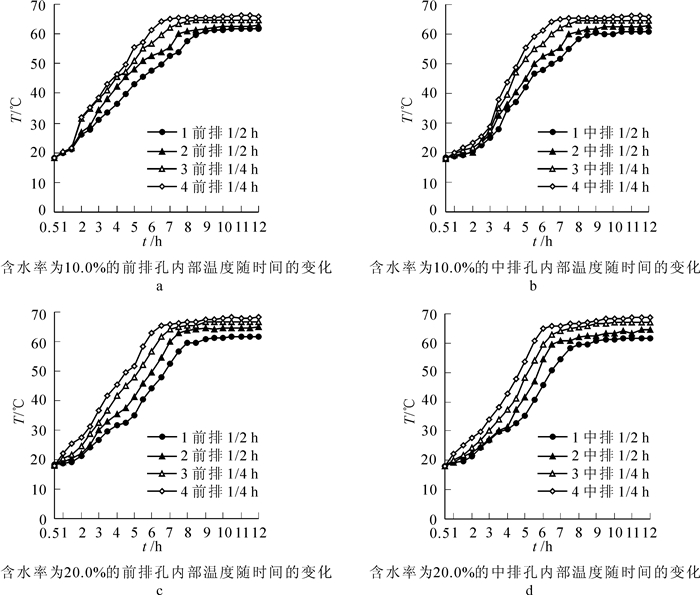
实验过程中分别采用含水率为10.0%和20.0%的重组竹来研究其内部的导热性能。如图 1所示:重组竹导热测试的打孔位置其中前排的1,2,3,4位于整个面的1/4处,中排的1,2,3,4位于整个面的1/2处。上述所有的孔1和孔2为深孔占总厚度h的1/2,孔3和孔4为浅孔占总厚度h的1/4,将温度测试仪的探头插入孔中隔0.5 h定点的测孔内温度的变化,目的就是为了通过测试重组竹内部的温度来找到最佳的浸渍时间点,即重组竹内部温度趋于平衡的点。因为竹材和木材的成分和结构很相似,所以竹材软化与木材软化具有相似的性质,因为水热处理(水煮法)软化重组竹,主要是利用水对纤维素的非结晶区、半纤维素和木素进行润胀,为分子剧烈运动提供自由体积空间,靠由外到里逐渐对重组竹进行传导加热。重组竹方材经水热处理后,一部分半纤维素易分解溶解成液态,便于在外力作用下产生相对滑移,从而使重组竹方材软化,加热可以使非结晶区中的木素、纤维素和半纤维素分子能量加大。在水、热的作用下,纤维素非结晶区湿胀,木素呈黏流态,半纤维素失去其联结作用,重组竹塑性加大即重组竹的软化程度提高,这样会有利于后期的刨切[11-15]。
图 4所示为重组竹在70.0 ℃的水浴中水煮,各孔内部温度随时间的变化,图 4中显示内部各孔温度变化的趋势都是大致相同的,都是经历了缓慢升温、快速升温和温度相对平衡的过程,且随着时间的延长,最终8.0 h各孔的温度都趋于稳定。图 4中所示:孔的升温速率和最终的温度大小依次为孔1>孔4>孔2>孔3,说明了孔内温度的传递方式主要为端部沿长度方向传导,而沿厚度方向为次或速率不明显,原因主要与竹材的纤维结构有关。通过比较图 4A与图 4C,图 4B与图 4D可知:重组竹的含水率越高,则其内部导热速率越快,到达相对平衡温度的时间越短。其中图 4A图 4B图所示的前排和中排孔最快在6.0 h左右达到稳定;图 4C图 4D图所示在5.0 h左右达到稳定,所以在实际操作中为了后期的刨切质量,应增大竹方的含水率,这样可以在水分和热的作用下,更好地软化重组竹。结合水煮软化试验和导热分析试验可知:当重组竹内部温度全部达到稳定时约在8.0 h左右,而水煮软化最佳的浸注时间为36.0 h,所以在重组竹水煮软化过程中的最佳浸注时间即为试验的最佳时间。

图 4 孔内部温度随时间的变化
Figure 4. Internal temperature change of the hole with time
-
从表 1中可以看出:在70.0 ℃软化36.0 h后,重组竹材的硬度下降明显。和未软化处理重组竹对比,经70.0 ℃软化36.0 h后,硬度下降了36.2%,P值小于0.01。说明处理前后样品的硬度差异还是非常显著的,采用水煮软化处理工艺能显著提高重组竹的硬度。
表 1 70.0 ℃软化36.0 h和对照组重组竹材的硬度
Table 1. Hardness of recombinant bamboo after no treatment and softening treatment at 70.0 ℃ for 36.0 h
试验号 未处理材(含水率 10.0%~12.0%) 70℃处理 36.0h(含水率 25.0%~27.0%) 1 2 670 1 820 2 2 580 1 590 3 2 730 1 570 4 2 100 1 680 5 2 620 1 430 6 2 380 1 320 7 2 290 1 710 8 2 610 1 630 9 2 750 1 540 10 2 270 1 650 平均 2 500 1 594 P值 2.602E-09 -
软化处理前,肉眼观察发现,重组竹表面裂纹都比较细小。原因为重组竹在压制的过程中,碾压竹丝和铺装时,都会使竹材维管束的方向发生改变,竹丝只是相对的平行,很多维管束都会发生倾斜和弯曲,这些倾斜和弯曲会在一定程度上抑制开裂的蔓延,所以重组竹表面的裂纹较细短[16];在本研究的条件下软化处理后试件表面肉眼观察到裂纹的变化情况如表 2。
表 2 重组竹表面裂纹情况
Table 2. Cracks in the surface of recombinant bamboo
测试指标 未处理材
(含水率10.0%~12.0%)70℃处理36.0h
(含水率25.0%~27.0%)裂纹数/条 12 12 裂纹长度/mm(平均值) 15.20 18.40 裂纹宽度/mm(平均值) 0.18 0.21 从表 2中可以看出:在70.0 ℃软化36.0 h后,重组竹材的表面裂纹数没有变化而裂纹宽度、裂纹长度有所增加。和未软化处理重组竹对比,经70.0 ℃软化36.0 h后,表面裂纹长度平均增加了21.1%,裂纹宽度平均增加了16.7%,说明水煮软化处理会在一定程度上增加裂纹的长度和宽度。
-
在本研究重组竹方材的加压浸注过程中,随着压力的增长和加压时间的延长,重组竹方材的增重率呈均匀升高的趋势,当压力大于1.2 MPa后,增重率增加趋势逐渐减缓;加压时间大于8.0 h后增重率增大的趋势逐渐减缓。
在本研究的水煮软化工艺中,重组竹方材的增重率随着温度的升高变化率逐渐增大,并且随着水煮时间的延长,重组竹方材的增重率逐渐增大,但是在36.0 h以后增大的趋势趋于缓慢。
传热试验表明:重组竹和木材的导热方向都是端部沿长度方向为主,厚度方向为次。
硬度测试结果表明:经70.0 ℃软化36.0 h后,重组竹材的硬度下降明显,和未处理材相比下降了36.2%。
裂纹测试结果表明:经70.0 ℃软化36.0 h后,表面裂纹长度增加了21.1%,裂纹宽度增加了16.7%,而裂纹数没有变化。
Humidifying and softening processes of recombinant bamboo
-
摘要: 针对重组竹方材含水率低、密度大且难刨切的现状,采用不同压力的加压浸注和不同温度的水煮软化工艺对重组竹方材进行增湿软化处理,研究增湿软化后重组竹方材的增重率、传热及硬度的变化情况。研究结果表明:随着压力的增大和时间的延续,重组竹方材的增重率逐步增大,压力1.2 MPa以后增重率增加不明显;随着水煮软化温度的升高和时间的延续,重组竹方材的增重率变化率呈增大的趋势,36.0 h后增大趋势变缓;重组竹内部温度在8.0 h时已趋于稳定。硬度测试结果表明,经过加压浸渍和水煮软化后,重组竹材的硬度下降明显,和未处理材相比下降了36.2%,而重组竹表面的裂纹长度和宽度有所增加,和未处理材相比裂纹平均长度增加了21.1%,平均宽度增加了16.7%。说明采用加压浸注和水煮软化的生产工艺能对重组竹方材起到较好的软化效果,但也在一定程度上影响了其表面的裂纹长度和宽度。图4表2参16Abstract: Recombinant bamboo has many good properties, such as nice outlook, fine phisical properties. However, recombinant bamboo has a low moisture content and high density. To determine changes in the weight gain rate and to provide temperature and hardness guidance for humidifying and softening recombinant bamboo, the technique of pressure impregnating and water softening with different pressures and temperatures were conducted. Results showed that with an increase in pressure and time, the weight gain rate increased, but above 1.2 MPa increases in the weight gain rate were not obvious. For the water softening process, with increases of temperature and time, the weight gain rate increased, but slowed after 36.0 h; the internal temperature was stable after 8.0 h. Compared to the control, pressure impregnating and water softening decreased hardness 36.2%. Also, compared to the control group, the crack test showed that crack length increased 21.1% and crack width increased 16.7%. Thus, the manufacturing process had a great positive softening effect on recombinant bamboo, and it had a small nagetive effect on crack length and width with recombinant bamboo surfaces.[Ch,4 fig. 2 tab. 16 ref.]
-
图 1 重组竹导热测试的打孔位置
Figure 1 Punch position of thermal conductivity test of recombinant bamboo
图 2 压力对重组竹方材增重率的影响
Figure 2 Influence of pressrue on weight gain rate of recombinant bamboo
图 3 蒸煮温度对重组竹方材增重率的影响
Figure 3 Influence of temperature on weight gain rate of recombinant bamboo
表 1 70.0 ℃软化36.0 h和对照组重组竹材的硬度
Table 1. Hardness of recombinant bamboo after no treatment and softening treatment at 70.0 ℃ for 36.0 h
试验号 未处理材(含水率 10.0%~12.0%) 70℃处理 36.0h(含水率 25.0%~27.0%) 1 2 670 1 820 2 2 580 1 590 3 2 730 1 570 4 2 100 1 680 5 2 620 1 430 6 2 380 1 320 7 2 290 1 710 8 2 610 1 630 9 2 750 1 540 10 2 270 1 650 平均 2 500 1 594 P值 2.602E-09  下载: 导出CSV
下载: 导出CSV
表 2 重组竹表面裂纹情况
Table 2. Cracks in the surface of recombinant bamboo
测试指标 未处理材
(含水率10.0%~12.0%)70℃处理36.0h
(含水率25.0%~27.0%)裂纹数/条 12 12 裂纹长度/mm(平均值) 15.20 18.40 裂纹宽度/mm(平均值) 0.18 0.21
下载: 导出CSV
-
[1] 于文吉. 我国重组竹产业发展现状与趋势分析[J]. 木材工业, 2012, 26(1): 11-14. YU Wenji. Current status and future development of bamboo scrimber industry in China [J]. China Wood Ind, 2012, 26(1): 11-14. [2] 束必清, 肖忠平, 赵志高, 等. 重组竹框架结构的设计[J]. 林业科技开发, 2014, 28(5): 82-86. SHU Biqing, XIAO Zhongping, ZHAO Zhigao, et al. Design on reconstituted bamboo frame structure [J]. China For Technol, 2014, 28(5): 82-86. [3] 黄圣游, 吴智慧. 重组竹新中式家具的发展前景[J]. 竹子研究汇刊, 2010, 29(3): 1-4, 14. HUANG Shengyou, WU Zhihui. Prospects of new Chinese-style furniture made of recombinant bamboo [J]. J Bamboo Res, 2010, 29(3): 1-4,14. [4] 李延军, 杜春贵, 鲍滨福, 等. 大幅面刨切薄竹的生产工艺[J]. 木材工业, 2006, 20(4): 38-40. LI Yanjun, DU Chungui, BAO Binfu, et al. Techniques for making large size sliced bamboo veneer [J]. China Wood Ind, 2006, 20(4): 38-40. [5] 刘志坤, 李延军, 杜春贵, 等. 刨切薄竹生产工艺研究[J]. 浙江林学院学报, 2003, 20(3):227-231. LIU Zhikun, LI Yanjun, DU Chungui, et al. A study on production technology of sliced bamboo veneer [J]. J Zhejiang For Coll, 2003, 20(3): 227-231. [6] 章卫钢, 李延军, 方明俊. 竹集成材增湿处理技术研究[J]. 林业科技, 2010, 35(5):42-45. ZHANG Weigang, LI Yanjun, FANG Mingjun. The research of moistening treatment for glued laminated bamboo [J]. For Sci Technol, 2010, 35(5):42-45. [7] 程大莉, 蒋身学. 热处理对慈竹重组材性能的影响[J]. 林业科技开发, 2012, 26(4):73-75. CHENG Dali, JIANG Shenxue. Effect on properties of high temperature heat-treated Neosinocalamus affinis reconstituted bamboo lumber [J]. China For Technol, 2012, 26(4): 73-75. [8] 秦莉. 热处理对重组竹材物理力学及耐久性能影响的研究[D]. 北京:中国林业科学研究院, 2010. QIN Li. Effect of Thermo-treatment on Physical, Mechanical Properties and Durability of Reconstituted Bamboo Lumber [D]. Beijing:Chinese Academy of Forestry, 2010. [9] 于文吉, 余养伦, 周月, 等. 小径竹重组结构材性能影响因子的研究[J]. 林产工业, 2006, 33(6):24-28. YU Wenji, YU Yanglun, ZHOU Yue, et al. Studies on factors influencing properties of reconstituted engineering timber made from small-sized bamboo [J]. China For Pro Ind, 2006, 33(6): 24-28. [10] 戴仔财. 速生杉木横向复合型浸渍的研究[D]. 南京:南京林业大学, 2012. DAI Zaicai. The Research About Fast-growing Fir of Its Horizontal Compound Type Impregnation [D]. Nanjing:Nanjing Forestry University, 2012. [11] 李兆红. 论木材软化处理工艺[J]. 农村实用科技信息, 2013(6):57. LI Zhaohong. The softening process theory of timber [J]. Rural Prac Sci Technol Inf, 2013(6):57. [12] 陈淑梅. 木材软化处理技术及加工工艺[J]. 投资与合作, 2014 (2):131. CHEN Shumei. Softening technology and manufacturing process of wood [J]. Top Capital, 2014(2): 131. [13] RATTANADECHO P. The simulation of microwave heating of wood using a rectangular wave guide: influence of frequency and sample size [J]. Chem Eng Sci, 2006, 61(14): 4798-4811. [14] ÖRS Y, KESKIN H, ATAR M, COLAKOGLU M. Effects of impregnation with Imersol-aqua on the modulus of elasticity in bending of laminated wood materials [J]. Construct Build Mat, 2007, 21(8): 1647-1655. [15] TJEERDSMA B F, BOONSTRA M, PIZZI A, et al. Characterisation of thermally modified wood: molecular reasons for wood performance improvement [J]. Holz Roh Werkst, 1998, 56(3): 149-153. [16] 邹怡佳. 改性三聚氰胺树脂制备及其对竹材表面开裂影响[D]. 北京:中国林业科学研究院, 2013. ZOU Yijia. Preparation of Modified Melamine Formaldehyde Resin and Its Effects on Surface Cracks of Bamboo-based Board [D]. Beijing:Chinese Academy of Forestry, 2013. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2016.03.015
 点击查看大图
点击查看大图
计量
- 文章访问数: 2917
- HTML全文浏览量: 698
- PDF下载量: 477
- 被引次数: 0
