








-
超声波是一种重要的木材无损检测技术,对超声波信号的传播参数进行分析可以测定木材的杨氏模量、密度、含水率等物理参数,还可以检测木材内部的腐朽、孔洞、节子等缺陷[1]。然而传统的超声波检测技术属于接触式检测范畴,需要在待测材料表面涂抹液态耦合剂或者在液体中进行浸入式检测,以便更好地将超声波导入待测材料。木材具有多孔性,并不适合采用这种方式进行检测[2]。空气耦合超声波是指超声波换能器与待测木材之间不接触、直接利用空气作为耦合介质的超声波检测方法[3-4]。这种检测方法克服了传统超声波检测需要液态耦合剂的固有缺陷,不会弄脏或者损坏待测木材。目前,空气耦合超声波已经被广泛应用于木材物理参数检测[5-6]、缺陷检测[2, 7]、人造板检测[8-9]、木板画检测[10]等。大量研究结果表明,空气耦合超声波的传播参数与木材的物理特性、内部缺陷等有密切的对应关系。利用空气耦合超声波技术,可以实现木材特性参数的精准、快速检测。目前,商业化的空气耦合超声波检测设备主要有美国Ultran公司的SencondWave M510,QMI公司的AirScan,日本探头株式会社的NAUT等。这些设备价格非常昂贵,而且是针对碳复合材料等其他材料研制的[11],限制了空气耦合超声波在中国木材加工检测领域中的应用。笔者研制了基于空气耦合超声波技术的木材扫描成像系统,并对该成像系统进行了实验测试,分析了测试结果。
-
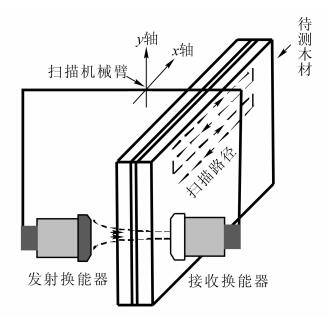
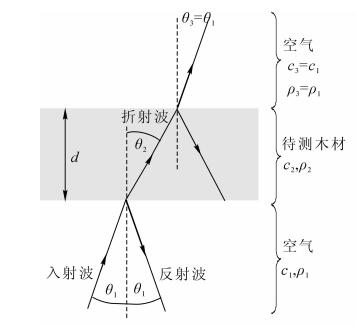
图 1为木材无损检测中常用的透射式空气耦合超声波检测系统。发射换能器激发出的超声波经过空气后直接进入待测木材,在穿过待测木材后再次经过空气,到达接收换能器。如图 1所示,在木材与空气的分界面上,超声波会出现折射与反射现象。设传播介质的密度为ρn,超声波在介质中的传播速度为Cn,其中n=1, 2, 3,分别对应于图 1中所示的空气层、待测木材层和空气层。介质的声阻抗为:

图 1 透射式空气耦合超声波检测技术原理
Figure 1. Schematic of transmitted-through air-coupled ultrasonic technique
$$ {Z_n} = {\rho _n}{C_n}。 $$ (1) 超声波在2个“木材/空气”分界面上的透射率分别为[12]:
$$ {T_{12}} = \frac{{2 \cdot {Z_1} \cdot \cos {\theta _2}}}{{{Z_1} \cdot \cos {\theta _2} + {Z_2} \cdot \cos {\theta _1}}};{T_{23}} = \frac{{2 \cdot {Z_2} \cdot \cos {\theta _3}}}{{{Z_2} \cdot \cos {\theta _3} + {Z_3} \cdot \cos {\theta _2}}}。 $$ (2) 式(2)中:θ1为超声波的入射角,θ2和θ3均为超声波的折射角。若超声波垂直入射,即入射角θ1=0°,则超声波经过2个分界面后总的透射率为:
$$ {T_{123}} = \frac{{4{Z_1}{Z_2}}}{{{{\left( {{Z_1} + {Z_2}} \right)}^2}}}。 $$ (3) 在空气声阻抗Z1固定不变的情况下,T123的大小与待测木材的声阻抗Z2密切相关。Z2越接近空气的声阻抗Z1,则T123越大。根据式(1)可知,Z2的大小与木材密度ρ2成正比,因此接收端的超声波信号幅度直接反映了待测木材的密度。超声波在木材内部传播时也会产生导致传播衰减,如吸收衰减和散射衰减。空气的声阻抗为415 Rayl,木材的典型声阻抗为1.57×106 Rayl,声阻抗失配非常严重[12-13],超声波在“空气/木材”分界面上损失的能量远大于吸收衰减以及材料不均匀性引起的散射衰减[14]。
-
图 2所示为空气耦合超声波扫描成像系统的结构图,将匹配的换能器对固定在龙门式扫描机械臂上,在木材的待测区域内沿图 2中虚线所示的路线进行逐点检测,利用超声波声速或传播衰减构建图像。

图 2 空气耦合超声波扫描成像系统原理图
Figure 2. Schematic of air-coupled ultrasonic imaging system
-
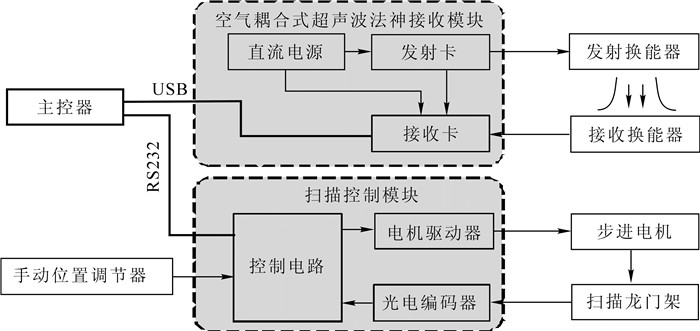
图 3显示了空气耦合超声波木材扫描成像系统的结构图,主要包含3个部分:主控器、空气耦合超声波发射接收模块、扫描控制模块。主控器是系统的核心,由工控机实现,主要功能为控制各模块协同工作,并根据采集到的超声波信号构建待测材料的扫描图像;空气耦合超声波发射接收模块与主控器之间通过USB总线连接,其主要功能是产生特定频率、幅度的脉冲信号,激励发射换能器产生超声波,空气耦合超声波发射接收模块还与接收换能器一起实现超声波信号的采集功能;扫描控制模块与主控器之间通过RS232接口进行连接,主要任务是根据设定的扫描路线在待检测区域内移动超声波换能器,实现逐点检测。

图 3 空气耦合超声波木材扫描成像系统框图
Figure 3. Structure of the air-coupled ultrasonic imaging system
-
由于高频超声波在木材中传播时会迅速衰减[1],所以在木材无损检测中,通常选用低频超声波换能器。选用Airstar公司的空气耦合超声波换能器,中心频率为125 kHz。还选用了Airstar公司的AS2100型空气耦合超声波发射接收器,实现超声信号的激励与采集。该发射接收器由发射卡与接收卡组成,采用双通道12 V/1.7 A的直流电源供电。发射卡与接收卡之间用6芯电缆连接。接收卡上有USB接口,完成主控器与超声波发射接收器之间的数据通讯。超声波换能器及发射接收器的各项参数如表 1所示。
表 1 超声波换能器及发射接收器的性能参数
Table 1. Parameters of the ultrasonic transducers, pulser and receiver
类别 参数名称 取值范围 换能器 中心频率 125 kHz 波束直径 5 mm 发射卡 脉冲波形 方波脉冲串 脉冲电压 0~200 V,步长10 V 脉冲串频率 10?50串.s-1,步长1串.s-1 串内脉冲个数 1~10个,步长1个 串内载波频率 30~500 kHz,步长1 kHz 接收卡 增益 0~255 dB,步长1 dB 采样频率 100,120,150,200,300, 400,500,800,1 000,2 000 kHz 模/数转换位数 双极性13位 -
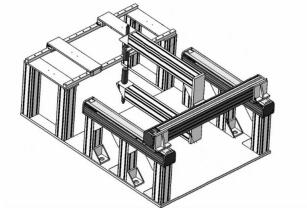
扫描机械臂采用龙门式结构,如图 4所示,由x,y 2个方向的滑台构成。x,y方向均采用步进电机驱动,采用同步带传动。步进电机主轴的运动通过锥齿轮箱减速、换向后,从箱的两端出轴带动两组同步带轮转动,直线导轨上的滑块与同步带固定相联,因此同步带可带动滑块自由滑动。x方向的扫描行程为500 mm,y方向的扫描行程为300 mm,单轴精度均为0.01 mm。扫描机械臂的控制系统采用STC12C5A60S2单片机作为核心控制器,主要完成3项任务:①根据手动位置调节器的指令控制x方向和y方向的步进电机进行控制,使空气耦合超声波换能器移动到指定的位置;②根据上位机的指令控制x方向和y方向的步进电机,使空气耦合超声波换能器沿设定的扫描路线进行自动扫描;③显示当前电机的状态。

图 4 龙门式扫描机械臂的结构图
Figure 4. Structure of the gantry scanner
-
上位机软件采用Borland C++ Builder编写,主要包含人机接口、扫描过程控制、数据通讯等模块,软件系统结构如图 5所示。人机接口模块的主要作用是提供接口,使用户可以根据测试需求设置超声波频率、电压、采样频率、扫描间隔等参数,以及观察扫描结果等。扫描过程控制的主要功能是根据用户设置的扫描参数向超声波发射接收器、龙门式扫描机械臂的驱动程序发出命令,实现整个系统协调工作。数据通讯驱动模块则完成命令发送、数据接收等功能。扫描结束后,软件保存扫描图像,并以文本文件形式保存每次扫描的结果,为今后利用第3方软件分析数据提供便利。

图 5 软件系统结构图
Figure 5. Structure of the software modules
-
研制的空气耦合超声波木材扫描成像系统以及软件界面如图 6所示。为了测试空气耦合超声波木材成像仪的准确性,采用多块木板分别进行测试。

图 6 研制的木材扫描成像系统实物图
Figure 6. The photo of the developed wood imaging system
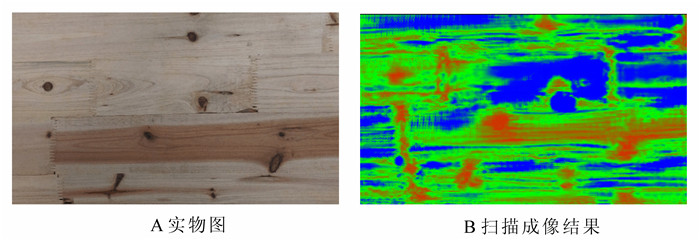
图 7所示为含节子缺陷的水杉Metasequoia glyptostroboides木板照片,厚度为13 mm,在节子中间(实线方框内)有一肉眼可以识别的裂缝。采用研制的空气耦合超声波木材扫描成像系统对其进行扫描,使用的参数为:脉冲电压为200 V,串内脉冲个数为5,脉冲串频率为50 Hz,串内载波频率110 kHz,接收信号放大增益为100 dB,采样频率为500 kHz,扫描间隔为0.5 mm,扫描区域为370 mm×110 mm。

图 7 含节子缺陷的水杉木板实物图
Figure 7. The photo of the Metasequoia glyptostroboides sample with knot
首先通过手动位置调节器将超声波换能器分别移至图 7中A,B,C点位置上,采集穿透木材后的超声波信号,并进行归一化处理,各位置上采集到的信号波形如图 8所示。如前所述,不同位置上的超声波信号幅度受材质密度的影响较大。B点位于节子区域,木材的密度较大,所以传播衰减较大,接收到的超声波信号幅度较小。C点位于节子中间的裂缝区域,超声波传播过程中,不需要穿透木材,因此接收到的超声波信号幅度最大。A点所在区域为早材区域,密度较低,肉眼观察也可以判断出A点附近的材质较为疏松,与B点上采集到的信号相比,信号幅度较大;但与C点上采集到的信号相比,其信号幅度快速衰减。

图 8 不同位置上采集到的扫描信号
Figure 8. Signals received at different positions
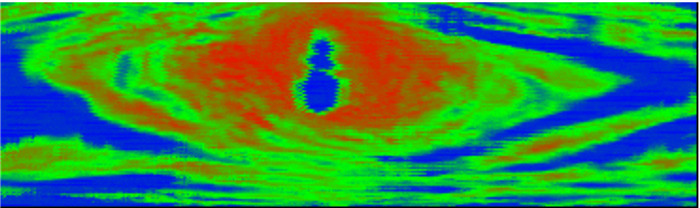
采用研制的空气耦合超声波木材成像仪对图 7所示的木材进行扫描成像,结果如图 9所示。图中红色表示木材的材质密度最大,接收到的超声波信号幅度最低;蓝色表示木材的密度最小,或者所在区域为裂缝、空洞,接收到的超声波信号幅度最大。从图 9中可以清晰地观察到节子以及节子中间的裂缝,扫描图像基本符合样本木材的实际情况。对比图 7,图 9显示的节子以及裂缝明显偏大,是因为超声波换能器发出的是5 mm粗的波束,致使扫描图像出现畸变。

图 9 含节子木材样本成像结果
Figure 9. The imaging result of the sample with knot
图 10A所示为15 mm厚的杉木Cunninghamia lanceolata指接板实物图,采用研制的空气耦合超声波扫描成像系统对其进行检测。所用参数为:脉冲电压为100 V,串内载波频率为120 kHz,串内脉冲个数为10个,脉冲串频率为50 Hz,接收信号放大增益为为80 dB,采样频率为500 kHz,扫描间隔为0.5 mm,扫描区域为450 mm×260 mm。扫描图像如图 10B所示。从图 10中可以看出,节子处对应的扫描图像均为红色;深色心材(密度较大)对应的扫描图像为红色;纹理较稀疏的区域(密度较低)对应的扫描图像为蓝色。指接处含有与木质材料不同的胶合剂,会增大超声波的传播衰减,对应的超声波扫描图像也为红色。扫描图像较好地反应了杉木指接板的平面密度分布情况。

图 10 指接板实物图及扫描成像结果
Figure 10. The photo and the imaging result of the finger joint panel
-
根据空气耦合超声波的原理,研究并实现了用于木材无损检测的空气耦合超声波扫描成像系统。设计了扫描成像系统的超声波发射接收系统、龙门式扫描机械臂及其控制系统,编写了上位机软件。将笔者研制的空气耦合扫描成像系统应用于水杉、杉木指接板等木材的扫描成像中。实验结果表明:利用该扫描成像系统建立的扫描图像能够准确地检测出节子、裂缝等木材缺陷,也能检测出木材在平面内的密度变化情况,具有较大的应用潜力。下一步将研究扫描图像的校正工作,减少超声波波束直径引起的图像误差,为提高木材无损检测准确度提供技术保障。
An air-coupled ultrasonic imaging system for non-destructive wood testing
-
摘要: 空气耦合超声波不需要液态耦合剂,可以实现真正的非接触式无损检测,已成为一种可靠的木材无损检测技术,但国内还没有相应的检测设备。介绍了空气耦合超声波及成像技术的原理,研制了一种用于木材无损检测的空气耦合超声波扫描成像系统。详细介绍了超声波发射接收系统、龙门式扫描机械臂及其控制系统、上位机软件等模块的设计方法。设计的系统可以实现单轴精度为0.01 mm的二维扫描,最大扫描区域为500 mm×300 mm。还可以针对样本木材的特性,方便地设定空气耦合超声波的频率、幅度、增益等参数。采用水杉Metasequoia glyptostroboides实木板、杉木Cunninghamia lanceolata指接板进行了实验,空气耦合超声波信号的幅度对木材裂缝、节子以及密度变化等较为敏感。该系统生成的扫描图像可以清晰地反映样本木材中裂缝、节子以及指接处的位置、形状和大小等信息。该系统可以应用于木材裂缝、节子等缺陷的无损检测,也可以用于木材在平面内的密度分布检测。Abstract: Air-coupled ultrasonic techniques, allowing contactless nondestructive testing using air as the coupling media instead of a liquid, have proven to be reliable and indispensable for nondestructive wood testing; however, instrumentation is still not widely available domestically. To help familiarize prospective users, the principle of the air-coupled ultrasonic was described first. Subsequently, a scanning imaging system was developed and its structure comprising a pulser and receiver, a gantry scanner, and computer software, was described in detail. Two-dimensional scanning imaging was conducted with a single-spindle accuracy of 0.01 mm and a maximum scanning area of 500 mm×300 mm. Scanning parameters, such as frequency, amplitude, and gain, were set conveniently according to the sample under testing. Metasequoia glyptostroboides board and Cunninghamia lanceolata finger jointed board were utilized to test the designed scanning imaging system. Results showed that amplitude of the received signal was sensitive to cracks, knots, and density. The constructed images provided information on knots, cracks, and finger joints, including location, shape, and size. Based on preliminary experimental results, the developed scanning imaging system could be used for non-destructive detection of defects including knots and cracks and could be utilized to scan a wooden material for in-plane distribution of density.
-
Key words:
- wood science and technology /
- air-coupled ultrasonic /
- nondestructive testing of wood /
- scanning imaging /
- density /
- knots /
- cracks
-
图 1 透射式空气耦合超声波检测技术原理
Figure 1 Schematic of transmitted-through air-coupled ultrasonic technique
图 7 含节子缺陷的水杉木板实物图
Figure 7 The photo of the Metasequoia glyptostroboides sample with knot
图 10 指接板实物图及扫描成像结果
Figure 10 The photo and the imaging result of the finger joint panel
表 1 超声波换能器及发射接收器的性能参数
Table 1. Parameters of the ultrasonic transducers, pulser and receiver
类别 参数名称 取值范围 换能器 中心频率 125 kHz 波束直径 5 mm 发射卡 脉冲波形 方波脉冲串 脉冲电压 0~200 V,步长10 V 脉冲串频率 10?50串.s-1,步长1串.s-1 串内脉冲个数 1~10个,步长1个 串内载波频率 30~500 kHz,步长1 kHz 接收卡 增益 0~255 dB,步长1 dB 采样频率 100,120,150,200,300, 400,500,800,1 000,2 000 kHz 模/数转换位数 双极性13位  下载: 导出CSV
下载: 导出CSV
-
[1] SENALIK A C, SCHUENEMAN G, ROSS R J. Ultrasonic-based nondestructive evaluation methods for wood:a primer and historical review[G]//ROSS R J. Nondestructive Evaluation of Wood. 2nd ed. Madison:United States Department of Agriculture, 2015:21-51. [2] HSU D K, UTRATA D, KUO M. NDE of lumber and natural fiber based products with air coupled ultrasound[C]//Proceedings of the Review of Progress in Quantitative Nondestructive Evaluation. Kingston, RI:AIP, 2010:1533-1540. [3] 周正干, 魏东.空气耦合式超声波无损检测技术的发展[J].机械工程学报, 2007, 44(3):10-14. ZHOU Zhenggan, WEI Dong. Progress of air-coupled ultrasonic non-destructive testing technology[J]. Chin J Mech Eng, 2007, 44(3):10-14. [4] CHIMENTI D E. Review of air-coupled ultrasonic materials characterization[J]. Ultrasonics, 2014, 54(7):1804-1816. [5] HASEGAWA M, MORI M, MATSUMURA J. Nondestructive Prediction of Bending Strength Performance in Wood with Artificial Hole by Air-coupled Ultrasonic Wave[EB/OL].[2016-03-10] http://210.101.116.28/W_files/kiss6/06804543_pv.pdf. [6] FANG Yiming, LU Z X, LIN Lujun, et al. Application of air-coupled ultrasonic imaging technique for nondestructive testing of solid wood board[J]. ICIC Expr Lett Part B Appl, 2015, 6(10):2773-2778. [7] GAN T H, HUTCHINS D A, GREEN R J, et al. Noncontact, high-resolution ultrasonic imaging of wood samples using coded chirp waveforms[J]. IEEE Trans Ultrason Ferroelectr Freq Control, 2005, 52(2):280-288. [8] SANABRIA S J, HILBERS U, NEUENSCHWANDER J, et al. Modeling and prediction of density distribution and microstructure in particleboards from acoustic properties by correlation of non-contact high-resolution pulsed air-coupled ultrasound and X-ray images[J]. Ultrasonics, 2013, 53(1):157-170. [9] HILBERS U, NEUENSCHWANDER J, HASENER J, et al. Observation of interference effects in air-coupled ultrasonic inspection of wood-based panels[J]. Wood Sci Technol, 2012, 46(5):979-990. [10] SIDDIOLO A M, D'ACQUISTO L, MAEVA A R, et al. Wooden panel paintings investigation:an air-coupled ultrasonic imaging approach[J]. IEEE Trans Ultrason Ferroelectr Freq Control, 2007, 54(4):836-846. [11] 常俊杰, 卢超, 川嶋紘一郎.非接触空气耦合超声波的材料无损评价与检测[J].浙江理工大学学报 (自然科学版), 2015, 33(4):532-536, 542. CHANG Junjie, LU Chao, KAWASHIMA K. Nondestructive material evaluation and testing based on non-contact air-coupled ultrasonics[J]. J Zhejiang Inst Sci Technol, 2015, 33(4):532-536, 542. [12] FLEMING M R, BHARDWAJ M C, JANOWIAK J J, et al. Noncontact ultrasound detection of exotic insects in wood packing materials[J]. For Prod J, 2005, 55(6):33-37. [13] POUDEL A, STRYCEK J, CHU T P. Air-coupled ultrasonic testing of carbon/carbon composite aircraft brake disks[J]. Mater Eval, 2013, 71(8):987-994. [14] HILBERS U, THOEMEN H, HASENER J, et al. Effects of panel density and particle type on the ultrasonic transmission through wood-based panels[J]. Wood Sci Technol, 2012, 46(4):685-698. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2017.02.021
 点击查看大图
点击查看大图
计量
- 文章访问数: 4056
- HTML全文浏览量: 701
- PDF下载量: 749
- 被引次数: 0
