




-
欧洲鹅耳枥Carpinus betulus为桦木科Betulaceae鹅耳枥属Carpinus落叶乔木,原产于土耳其、乌克兰[1],树形清秀,枝叶浓密,叶形秀丽,秋叶金黄,果穗奇特[2],颇为美观,园艺品种众多,在欧美等地应用广泛,是观赏价值极高的园林绿化树种。对欧洲鹅耳枥引种驯化并进行生态适应性评价可为其推广应用提供理论基础及实践支撑,为城市绿化彩叶树种的匮乏提供新的资源。观赏价值和适应能力是决定观赏植物推广应用的2个关键因素[3]。欧洲鹅耳枥及其园艺品种观赏价值极高,因此其适应能力强弱是决定其能否大面积推广应用的决定性因素。目前,国内对植物稳定性和适应性评价的方法有多种,如Franics和Kannenberg模型、秩次分析法、高稳系数法、Shukla模型及AMMI模型等。其中,AMMI模型将方差分析和主成分分析有效地结合在一起,在分析基因型与环境互作效应方面有明显的优势,可定量地描述各品种稳定性差异及各地点对品种鉴别力的大小[4],已广泛地用于荞麦Fagopyrum esculentum,桤木Alnus cremastogyne,烤烟Nicotiana tabacum,小麦Triticum aestivum,水稻Oryza sativa,甘蔗Saccharum officinarum,棉花Gossypium hirsutum等[5-8]作物品种和植物种质资源的稳定性和适应性研究,但在观赏树木方面应用较少。本研究采用AMMI模型分析方法对欧洲鹅耳枥及其园艺品种在不同地区的适应性和稳定性进行评价,旨在找出各地适应性最好的种(品种)及适应性最广的种(品种),为欧洲鹅耳枥及其园艺品种在国内的推广和应用寻找更适宜的地点及范围。
-
试验地设在南京林业大学下蜀实习林场、江苏紫藤园艺绿化工程有限公司苗圃(靖江)和北京人地科技发展有限责任公司苗圃。3地自然概况如表 1。
表 1 试验地概况
Table 1. Description of experiment field
试验地点 气候 经纬度 年平均降水量/mm 年平均气温/℃ 年平均日照时间/h 无霜期/d 土壤类型 北京 暖温带大陆性季风气候 39°44’N,116°20’E 626 8.0-12.0 2 780.0 180-200 砂性土 南京 北亚热带季风气候 32°12’N, 119°26’E 1 105 15.1 2 018.0 229 壤土 靖江 亚热带季风气候 32°02’N, 120°16’E 1 062 15.3 2 114.6 230 黏性土 -
本试验所用的材料欧洲鹅耳枥及其2个园艺品种Carpinus betulus ‘Frans Fontaine’和C. betulus ‘Lucas’,均为法国进口的统一规格的3年生裸根苗,苗高40~60 cm。欧洲鹅耳枥为实生苗,2个园艺品种为嫁接苗,砧木为欧洲鹅耳枥,砧木及接穗规格基本一致。
-
2013年3月,将欧洲鹅耳枥及其2个园艺品种同时在南京、靖江和北京等3地室外地栽,各个种(品种)300株列植,栽植株行距为30 cm × 50 cm,100株设置1个单元,各个种(品种)设3个单元,设置标准株3株·单元-1,挂上标签。生长期间只进行简单的水分和杂草管理。地栽后,连续2 a观察3地欧洲鹅耳枥及其园艺品种生长期的物候变化,记录时间;卷尺测定苗高,精确到0.1 cm,游标卡尺测定地径,精确到0.002 cm。地栽当年3月测其标准株苗高和地径,翌年3月再次测量,苗高和地径净增量为其1 a净生长量。待苗木生长稳定后,于当年8月采集各个地方不同种(品种)枝条中上部成熟叶,用冰盒储藏带回试验室后迅速用叶面积仪测量叶长、叶宽、叶面积大小。翌年5月,计算苗木成活率,其公式为引种成活率(%)=(剩余存活苗木数量/初始栽植苗木数量)×100%。
-
采用Excel 2003进行数据处理分析和制图;采用SPSS 20.0进行方差分析、多重比较和相关分析;采用DPS v9.05进行AMMI模型(加性主效应乘积交互作用模型)计算。
AMMI模型公式[9]为yge=μ+αg+βe+Σλnγgnσen+θge+εger。其中:yge是在环境e中基因型g的产量,μ代表总体平均值,αg是基因型平均偏差,βe是环境的平均偏差,λn是第n个主成分分析的特征值,γgn是第n个主成分的基因型主成分得分,σen是第n个主成分的环境主成分得分,n是模型主成分分析中主成分因子轴的总个数,σen为残差,εger为误差,误差等于yge平均值与r个重复的单个观察值之间的偏差,并具有可加性。
-
表 2为欧洲鹅耳枥及其园艺品种的物候期。不同种(品种)欧洲鹅耳枥在同一地区物候差异不大。同一种(品种)欧洲鹅耳枥在3地物候差异较大,靖江和南京地区展叶时间较北京地区早20 d左右,二次抽梢时间较北京早30 d左右,叶变黄时间较北京地区晚约60 d,落叶时间较北京地区晚30 d左右(欧洲鹅耳枥除外)。3地病虫害情况亦有较大差别,总体而言,虫害严重程度为靖江>南京>北京。
表 2 3地欧洲鹅耳枥及其品种物候期
Table 2. Phenology of Carpinus betulus and its cultivars in three regions
物候期 地点 ‘Frans Fontaine’ ‘Lucas’ 欧州鹅耳枥 展叶时间 北京 4月下旬 4月下旬 4月下旬 南京 4月上旬 4月上旬 4月上旬 靖江 4月上旬 4月上旬 4月上旬 二次抽梢时间 北京 7月中旬 7月中旬 7月中旬 南京 6月下旬 6月上旬 6月下旬 靖江 6月中旬 6月中旬 6月中旬 叶变黄时间 北京 9月中旬 9月下旬 9月下旬 南京 11月中旬 11月下旬 11月下旬 靖江 11月下旬 11月下旬 11月下旬 落叶时间 北京 11月上旬 11月上旬 11月下旬 南京 12月上旬 12月上旬 叶枯而不落 靖江 12月上旬 12月上旬 叶枯而不落 病虫害 北京 刺蛾(较轻,夏季) 刺蛾(较轻,夏季) 刺蛾(较轻,夏季) 南京 蚜虫(轻,夏季) 蚜虫(轻,夏季) 蚜虫(轻,夏季) 靖江 刺蛾(重,夏季) 刺蛾(重,夏季) 刺蛾(重,夏季) -
苗高净增量是反映植物生长状况的重要指标。方差分析表明:不同种(品种)、不同地区对欧洲鹅耳枥苗高净增量的影响分别达到显著(P=0.026),极显著(P=0.000)水平,种(品种)和地区之间的交互效应极显著(P=0.002)。如表 3所示,针对不同地区同一种(品种)而言,‘Frans Fontaine’的苗高净增量南京地区最大,约为靖江的2倍,北京的4倍。‘Lucas’苗高净增量靖江和南京地区最大,差异不显著,净增量最小的是北京地区。欧洲鹅耳枥的苗高净增量南京地区最大,约为北京和靖江地区的3倍,北京和靖江地区差异不显著。针对同一地区不同种(品种)而言,北京和靖江地区3个种(品种)间苗高净增量差异不显著,南京地区‘Lucas’与其他2个种(品种)差异显著。南京地区3种欧洲鹅耳枥的苗高净增量均最大,说明它更适宜3种欧洲鹅耳枥的营养生长。北京地区3种欧洲鹅耳枥的苗高净增量均较小,说明它生长速度明显慢于南京和靖江地区。这可能是因为它地处北方,相对于南京和北京地区来说,展叶期较晚,落叶期又较早,光合周期短,营养积累少所致。
表 3 欧洲鹅耳枥3地形态指标
Table 3. Morphological parameters of Carpinu betulus in three regions
指标 地区 种(品种) ‘Frans Fontaine’ ‘Lucas’ 欧洲鹅耳枥 苗高净增量/cm 北京 ll.80±5.78aB 13.92±5.74aB 19.72±7.08aB 南京 50.98±l6.50aA 22.12±8.17bAB 54.72±6.61aA 靖江 21.86±5.47aB 24.48±7.42aA 18.84±7.22aB 地径净增量/cm 北京 0.36±0.10aA 0.49±0.21aA 0.45±0.14aB 南京 0.59±0.23aA 0.53±0.11aA 0.75±0.17 aA 靖江 0.40±0.05aA 0.43±0.08aA 0.54±0.11aB 叶长/cm 北京 7.40±0.57 aA 6.75±0.34bA 6.06±0.26cB 南京 7.63±0.73 aA 6.90±0.71aA 7.81±0.55 aA 靖江 6.12±0.33abB 5.74±0.51bB 6.28±0.17aB 叶宽/cm 北京 4.49±0.27 aA 4.33±0.22aA 3.64±0.20bB 南京 4.32±0.31abA 3.85±0.32bB 4.55±0.41aA 靖江 3.67±0.25abB 3.40±0.25bC 3.95±0.18aB 叶面积/cm2 北京 23.65±2.39aA 21.13±1.82bA 16.53±0.73cB 南京 23.52±3.27aA 18.45±2.50bA 26.21±3.73aA 靖江 16.31±1.73abB 14.12±2.14bB 18.43±0.95aB 说明:同行中不同的小写字母表示同一地区不同品种间差异显著(P<0.05);同列中不同的大写字母表示同一品种不同地区间差异显著(P<0.05)。 地径净增量是反映植物生长状况的又一重要指标。方差分析表明:欧洲鹅耳枥及其园艺品种的地径净增量种(品种)间差异不显著(P=0.073),不同地区间差异极显著(P=0.003),种(品种)和地区之间的交互效应不显著(P=0.405)。如表 3所示:针对不同地区同一种(品种)而言,‘Frans Fontaine’‘Lucas’地区间差异均不显著,欧洲鹅耳枥在南京地区的地径净增量最大,与其他2个地区差异显著。针对同一地区不同种(品种)而言,3地各个种(品种)间差异均不显著,说明种(品种)在地径净增量方面影响较小。这可能与欧洲鹅耳枥各种(品种)生长缓慢有关。
-
植物叶片的大小与其光合作用、蒸腾速率等密切相关。方差分析表明:欧洲鹅耳枥不同种(品种)、不同地区间叶长差异均极显著(P=0.010,P=0.000),叶宽不同种(品种)间差异显著(P=0.020),不同地区间差异极显著(P=0.000),不同种(品种)、不同地区间叶面积差异均达极显著差异(P=0.001,P=0.000),叶长、叶宽和叶面积3个指标的种(品种)和地区交互效应均极显著(P=0.003,P=0.000,P=0.000)。如表 3所示:对不同地区同一种(品种)而言,‘Frans Fontaine’的叶长、叶宽、叶面积与‘Lucas’的叶长、叶面积均为靖江地区最小,均与其他2个地区差异显著,南京和北京地区差异不显著,‘Lucas’的叶宽3地间差异均显著,分别为北京>南京>靖江。C. betulus的叶长、叶宽、叶面积均为南京地区最大,与其他2个地区差异显著。对同一地区不同种(品种)而言,北京地区叶长、叶面积3个种(品种)之间差异均显著,分别为‘Frans Fontaine’>‘Lucas’>欧洲鹅耳枥,叶宽为欧洲鹅耳枥最小,南京地区叶长3个种(品种)之间差异不显著,叶宽欧洲鹅耳枥与‘Lucas’间差异显著,且欧洲鹅耳枥>‘Lucas’,叶面积‘Lucas’最小,其他2个种(品种)差异不显著,靖江地区叶长、叶宽、叶面积均为欧洲鹅耳枥> ‘Lucas’,两者之间差异显著,均与‘Frans Fontaine’差异不显著。说明环境与基因型产生了交互作用。南京与北京地区自然环境与土壤环境差异较大,但两地‘Frans Fontaine’叶长、叶宽、叶面积与‘Lucas’的叶长、叶面积均无显著差异,而两地欧洲鹅耳枥的叶指标差异显著,说明‘Frans Fontaine’具有较广的适应性,其次是‘Lucas’,而欧洲鹅耳枥具有较强的特异性。南京与靖江地区自然环境相似,但两地每个种(品种)的叶长、叶宽与叶面积均差异显著,可能与土壤环境差异较大有关。
-
引种成活率是区域化试验及引种是否成功的基本指标。引种1 a后,欧洲鹅耳枥及其品种成活率如表 4所示,南京地区欧洲鹅耳枥及其各个品种的成活率总体较高,其次是靖江地区,稍差的是北京地区。南京和靖江地区‘Lucas’成活率最高,达98%,其次是‘Frans Fontaine’,成活率最低的为欧洲鹅耳枥。北京地区‘Frans Fontaine’成活率最高,其次是‘Lucas’,成活率最低的为欧洲鹅耳枥。经观察发现,北京地区欧洲鹅耳枥及其品种死亡集中于翌年3-4月,可能是春旱作用引起的,靖江地区欧洲鹅耳枥及其品种死亡集中于当年夏季8-9月,可能是热害引起的。相对来说,南京地区环境条件适中,总体更适合各欧洲鹅耳枥的生长。
表 4 欧洲鹅耳枥及其品种引种成活率
Table 4. Survival rate of Carpinus betulus and its cultivars
地点 成活率/% ‘Frans Fontaine’ ‘Lucas’ 欧洲鹅耳枥 北京 90.53 88.37 74.24 南京 97.33 100.00 96.33 靖江 96.15 98.00 89.13 -
各欧洲鹅耳枥的稳定性和适应性主要取决于基因型和环境互作效应的大小。鉴于欧洲鹅耳枥的生长指标—苗高净增量和形态指标—叶面积的基因和环境互作效益显著,以这2个指标作为代表,进行各欧洲鹅耳枥的稳定性和适应性分析。
-
由AMMI模型分析结果(表 5)可知:苗高净增量的基因、环境和基因×环境的变异平方和分别占总变异平方和的6.93%,44.46%和17.84%。说明试验中环境对苗高净增量的总变异起的作用最大,其次是基因×环境,基因对苗高净增量总变异所起作用最小。AMMI模型分析结果表明:基因型与环境的交互作用达到极显著水平(P=0.002 0),IPCA1的平方和占交互作用平方和的94.38%,残差的平方和占交互作用平方和的5.62%,说明交互作用大部分的变异集中在IPCA1轴上。叶面积的基因、环境和基因×环境的变异平方和分别占总变异平方和的10.30%,37.75%和28.88%。说明试验中环境对叶面积总变异所起的作用最大,其次是基因×环境的交互作用,基因型对叶面积总变异所起的作用最小。AMMI模型分析结果表明:基因型与环境的交互作用达到极显著水平(P=0.000 1),IPCA1的平方和占交互作用平方和的96.60%,残差的平方和占交互作用平方和的3.40%,说明交互作用大部分的变异集中在IPCA1轴上。
表 5 叶面积基因与环境的AMMI模型分析结果
Table 5. Analysis of variance of leaf area for AMMI mode
变异来源 自由度 苗高净增量 叶面积 离差平方和 均方 F值 P值 离差平方和 均方 F值 P值 总的 44 13 864.744 4 315.107 8 847.565 7 19.262 9 处理 8 9 599.524 4 1 199.940 6 10.127 9 0.000 1 651.990 4 81.498 8 15.001 7 0.000 1 基因 2 961.256 4 480.628 2 4.056 7 0.025 8 87.325 6 43.662 8 8.037 1 0.001 3 环境 2 6 164.343 1 3 082.171 6 26.014 6 0.000 1 319.925 3 159.962 6 29.444 7 0.000 1 交互作用 4 2 473.924 9 618.481 2 5.220 2 0.002 0 244.739 6 61.184 9 11.262 4 0.000 1 IPCA1 3 2 334.897 3 778.299 1 5.598 2 0.003 0 236.414 2 78.804 7 9.465 6 0.000 1 残差 1 139.027 6 139.027 6 8.325 4 8.325 4 误差 36 4 265.220 0 118.478 3 195.575 3 5.432 7 -
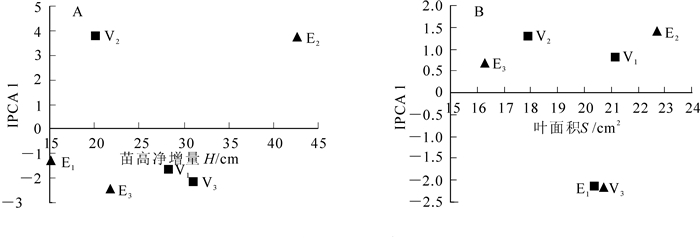
将各欧洲鹅耳枥的苗高净增量、叶面积作为横轴,将其各基因型在IPCA1上的得分作为纵轴,分别绘制苗高净增量与IPCA1的AMMI双标图(图 1A)和叶面积与IPCA1的AMMI双标图(图 1B)。由图 1可知:横轴方向表明试验地比种(品种)更分散,说明种(品种)的变异小于环境的变异,即同一种(品种)在不同地区表现的苗高净增量、叶面积差异较大,同一地点的各种(品种)苗高净增量、叶面积差异相对较小。纵轴方向表明基因与环境交互作用的差异,较为稳定的种(品种)靠近IPCA1值水平线。从苗高净增量的角度而言,各欧洲鹅耳枥稳定性为‘Frans Fontaine’(V1)>欧洲鹅耳枥(V3)>‘Lucas’(V2),苗高净增量为欧洲鹅耳枥(V3)>‘Frans Fontaine’(V1)>‘Lucas’(V2),各试验地苗高净增量为南京(E2)>靖江(E3)>北京(E1)。从叶面积角度而言,各欧洲鹅耳枥稳定性为‘Frans Fontaine’(V1)>‘Lucas’(V2)>欧洲鹅耳枥(V3),叶面积为‘Frans Fontaine’(V1)>欧洲鹅耳枥(V3)>‘Lucas’(V2),各试验地叶面积为南京(E2)>北京(E1)>靖江(E3)。此外,位于横轴同侧的种(品种)和地点之间存在正向交互效应,位于横轴2侧的种(品种)和地点之间存在负向的交互效应。从苗高净增量的角度而言,‘Frans Fontaine’(V1)和欧洲鹅耳枥(V3)与试验地北京(E1)和南京(E2)有正向交互作用,这2个种(品种)在该试点有较好的适应性,‘Lucas’(V2)在试验地南京(E2)有较好的适应性。从叶面积角度而言,‘Frans Fontaine’(V1)和‘Lucas’(V2)与试验地南京(E2)和靖江(E3)有正向交互作用,这2个种(品种)在该试点有较好的适应性,欧洲鹅耳枥(V3)在试验地北京(E1)有较好的适应性。

图 1 苗高净增量、叶面积与IPCA1的双标图
Figure 1. Biplot of net height, leaf area and IPCA1
-
区域化试验起初主要应用于农作物及经济作物等,如水稻、甘蔗等[10-13],近年来在园林植物方面也有应用,试验点一般分布较为均匀、具有代表性和一定的数量。在此基础上,对植物的适应性评价主要分为3个目标,即产量、抗逆性和景观效果[3, 14-15]。本研究的试验材料——欧洲鹅耳枥及其2个园艺品种景观效果相似,其生长速度影响着景观建成的速度,因此,采用生长指标及形态指标作为代表进行适应性评价,但在试验地的选择上存在一定的缺陷,即靖江和南京地区相距较近,自然环境相似,选址不够典型。因其土壤条件相差较大,因此,在这2个试验地进行区域化试验是可行的,也具有一定的意义。本试验中欧洲鹅耳枥及其园艺品种在各地生长时基因型与环境互作效应显著,环境对其生长影响最大,其次是环境与基因型的互作效应,基因型对其生长的影响最小。这与国内外很多作物对产量的研究结果一致[5],但亦存在某些作物基因型对生长的影响大于环境[13],可能是因为其区域试验点相距均较近,环境差异较小所致。
从物候的角度而言,北京地区欧洲鹅耳枥及其品种展叶晚,落叶早,其植株营养积累时间较南京和靖江地区少,因此生长速度偏慢。南京和靖江地区纬度较北京地区低,夏季炎热,高温和虫害是影响其生长的主要因素。综合物候、生长状况及成活率来看,南京地区是3种欧洲鹅耳枥的最适生长地,北京地区成活率稍差,靖江地区病虫害较严重导致景观效果较差;生长速度最快且最易形成景观效果的品种是Carpinus betulus ‘Frans Fontaine’,其次是欧洲鹅耳枥,‘Lucas’生长速度最慢,形成景观效果所需时间也较长。综合生长指标与形态指标的AMMI模型分析,辅以各欧洲鹅耳枥在3地的田间表现得出:各欧洲鹅耳枥稳定性最大且生长速度较快的是‘Frans Fontaine’,其次是欧洲鹅耳枥,‘Lucas’。最适宜欧洲鹅耳枥及其2个品种生长的试验地为南京地区,其次为北京和靖江。‘Frans Fontaine’在南京和北京地区适应性较好,‘Lucas’在南京和靖江地区适应性较好,欧洲鹅耳枥在南京地区适应性较好。
Regional adaptability trials for Carpinus betulus in China
-
摘要: 为研究欧洲鹅耳枥Carpinus betulus及其园艺品种在国内的引种适应性,以欧洲鹅耳枥及其2个园艺品种C. betulus‘Frans Fontaine’,C. betulus‘Lucas’为试验材料,观测它们在北京、南京、靖江等3地物候、生长和形态差异,并采用AMMI模型对它们进行适应性评价。研究表明:欧洲鹅耳枥及其2个品种在各地生长时基因型与环境交互作用显著(P<0.05)。C. betulus‘Frans Fontaine’稳定性最好且生长速度较快,其次是欧洲鹅耳枥,C. betulus‘Lucas’。3地相比较而言,南京为最适宜欧洲鹅耳枥及其2个品种生长的试验地,其次为北京和靖江。C. betulus‘Frans Fontaine’在南京和北京地区适应性较好,C. betulus‘Lucas’在南京和靖江地区适应性较好,欧洲鹅耳枥在南京地区适应性较好。Abstract: To research adaptability of Carpinus betulus in China, C. betulus and its two cultivars (C. betulus ‘Frans Fontaine’ and C. betulus ‘Lucas’) were used as experimental materials with the phenological period, growth condition, and morphological index being observed in Beijing, Nanjing, and Jingjiang. The Additive Main Effects and Multiplicative Interaction (AMMI) Model was adopted to evaluate adaptability through index of net hight and leaf area among C. betulus and its two cultivars in three regions. Results showed that the interaction between genotype and environment was strong to net hight(P=0.002 0) and leaf area(P=0.000 1). Take phenology, morphological parameters, survival rate and AMMI model analysis togethter, C. betulus ‘Frans Fontaine’ was the most stable and had the fastest growth followed by C. betulus and C. betulus ‘Lucas’; Nanjing was the most suitable region for growth of C. betulus and its two cultivars followed by Beijing and Jingjiang; for C. betulus ‘Frans Fontaine’ adaptability was better in Nanjing and Beijing, for C. betulus ‘Lucas' adaptability was better in Nanjing and Jingjiang, and for C. betulus adaptability was better in Nanjing. Therefor, C. betulus and its two cultivars can be introduced and cultured in Nanjing, C. betulus ‘Frans Fontaine’ can be introduced and cultured in Beijing, and C. betulus ‘Lucas’ can be introduced and cultured in Jingjiang.
-
Key words:
- forest tree breeding /
- Carpinus betulus /
- regional trials /
- adaptability evaluation
-
图 1 苗高净增量、叶面积与IPCA1的双标图
E为试验地点;V为种(品种)。E1. 北京;E2. 南京;E3. 靖江;V1. ‘Frans Fontaine’;V2. ‘Lucas’;V3. 欧洲鹅耳枥。
Figure 1 Biplot of net height, leaf area and IPCA1
表 1 试验地概况
Table 1. Description of experiment field
试验地点 气候 经纬度 年平均降水量/mm 年平均气温/℃ 年平均日照时间/h 无霜期/d 土壤类型 北京 暖温带大陆性季风气候 39°44’N,116°20’E 626 8.0-12.0 2 780.0 180-200 砂性土 南京 北亚热带季风气候 32°12’N, 119°26’E 1 105 15.1 2 018.0 229 壤土 靖江 亚热带季风气候 32°02’N, 120°16’E 1 062 15.3 2 114.6 230 黏性土  下载: 导出CSV
下载: 导出CSV
表 2 3地欧洲鹅耳枥及其品种物候期
Table 2. Phenology of Carpinus betulus and its cultivars in three regions
物候期 地点 ‘Frans Fontaine’ ‘Lucas’ 欧州鹅耳枥 展叶时间 北京 4月下旬 4月下旬 4月下旬 南京 4月上旬 4月上旬 4月上旬 靖江 4月上旬 4月上旬 4月上旬 二次抽梢时间 北京 7月中旬 7月中旬 7月中旬 南京 6月下旬 6月上旬 6月下旬 靖江 6月中旬 6月中旬 6月中旬 叶变黄时间 北京 9月中旬 9月下旬 9月下旬 南京 11月中旬 11月下旬 11月下旬 靖江 11月下旬 11月下旬 11月下旬 落叶时间 北京 11月上旬 11月上旬 11月下旬 南京 12月上旬 12月上旬 叶枯而不落 靖江 12月上旬 12月上旬 叶枯而不落 病虫害 北京 刺蛾(较轻,夏季) 刺蛾(较轻,夏季) 刺蛾(较轻,夏季) 南京 蚜虫(轻,夏季) 蚜虫(轻,夏季) 蚜虫(轻,夏季) 靖江 刺蛾(重,夏季) 刺蛾(重,夏季) 刺蛾(重,夏季)
下载: 导出CSV
表 3 欧洲鹅耳枥3地形态指标
Table 3. Morphological parameters of Carpinu betulus in three regions
指标 地区 种(品种) ‘Frans Fontaine’ ‘Lucas’ 欧洲鹅耳枥 苗高净增量/cm 北京 ll.80±5.78aB 13.92±5.74aB 19.72±7.08aB 南京 50.98±l6.50aA 22.12±8.17bAB 54.72±6.61aA 靖江 21.86±5.47aB 24.48±7.42aA 18.84±7.22aB 地径净增量/cm 北京 0.36±0.10aA 0.49±0.21aA 0.45±0.14aB 南京 0.59±0.23aA 0.53±0.11aA 0.75±0.17 aA 靖江 0.40±0.05aA 0.43±0.08aA 0.54±0.11aB 叶长/cm 北京 7.40±0.57 aA 6.75±0.34bA 6.06±0.26cB 南京 7.63±0.73 aA 6.90±0.71aA 7.81±0.55 aA 靖江 6.12±0.33abB 5.74±0.51bB 6.28±0.17aB 叶宽/cm 北京 4.49±0.27 aA 4.33±0.22aA 3.64±0.20bB 南京 4.32±0.31abA 3.85±0.32bB 4.55±0.41aA 靖江 3.67±0.25abB 3.40±0.25bC 3.95±0.18aB 叶面积/cm2 北京 23.65±2.39aA 21.13±1.82bA 16.53±0.73cB 南京 23.52±3.27aA 18.45±2.50bA 26.21±3.73aA 靖江 16.31±1.73abB 14.12±2.14bB 18.43±0.95aB 说明:同行中不同的小写字母表示同一地区不同品种间差异显著(P<0.05);同列中不同的大写字母表示同一品种不同地区间差异显著(P<0.05)。
下载: 导出CSV
表 4 欧洲鹅耳枥及其品种引种成活率
Table 4. Survival rate of Carpinus betulus and its cultivars
地点 成活率/% ‘Frans Fontaine’ ‘Lucas’ 欧洲鹅耳枥 北京 90.53 88.37 74.24 南京 97.33 100.00 96.33 靖江 96.15 98.00 89.13
下载: 导出CSV
表 5 叶面积基因与环境的AMMI模型分析结果
Table 5. Analysis of variance of leaf area for AMMI mode
变异来源 自由度 苗高净增量 叶面积 离差平方和 均方 F值 P值 离差平方和 均方 F值 P值 总的 44 13 864.744 4 315.107 8 847.565 7 19.262 9 处理 8 9 599.524 4 1 199.940 6 10.127 9 0.000 1 651.990 4 81.498 8 15.001 7 0.000 1 基因 2 961.256 4 480.628 2 4.056 7 0.025 8 87.325 6 43.662 8 8.037 1 0.001 3 环境 2 6 164.343 1 3 082.171 6 26.014 6 0.000 1 319.925 3 159.962 6 29.444 7 0.000 1 交互作用 4 2 473.924 9 618.481 2 5.220 2 0.002 0 244.739 6 61.184 9 11.262 4 0.000 1 IPCA1 3 2 334.897 3 778.299 1 5.598 2 0.003 0 236.414 2 78.804 7 9.465 6 0.000 1 残差 1 139.027 6 139.027 6 8.325 4 8.325 4 误差 36 4 265.220 0 118.478 3 195.575 3 5.432 7
下载: 导出CSV
-
[1] 施曼, 程龙霞, 祝遵凌. 不同地区欧洲鹅耳枥光合特性研究[J]. 西南农业学报, 2015, 28(1): 105-109. SHI Man, CHENG Longxia, ZHU Zunling. Studies on photosynthesis characteristic of Carpinus betulus in different areas [J]. Southwest China J Agric Sci, 2015, 28(1): 105-109. [2] GILMAN E F, WATSON D G. Carpinus betulus [R]. Fact Sheet ST-118, 1993. [2015-07-11] http://hort.ufl.edu/trees/CARBETA.pdf [3] 赵天荣, 徐志豪, 施永泰, 等. 大花萱草在宁波地区观赏性和适应性的综合评价[J]. 浙江农业学报, 2015, 27(4): 560-566. ZHAO Tianrong, XU Zhihao, SHI Yongtai, et al. Evaluation of ornamental value and adaptability for Hemerocallis hybridus in Ningbo [J]. Acta Agric Zhejiang, 2015, 27(4): 560-566. [4] 李本贵, 阎俊, 何中虎, 等. 用AMMI模型分析作物区域试验中的地点鉴别力[J]. 作物学报, 2004, 30(6): 593-596. LI Bengui, YAN Jun, HE Zhonghu, et al. Analyzing site discrimination in crop regional yield trials by AMMI model [J]. Acta Agron Sin, 2004, 30(6): 593-596. [5] 常磊, 柴守玺. AMMI模型在旱地春小麦稳定性分析中的应用[J]. 生态学报, 2006, 26(11): 3677-3684. CHANG Lei, CHAI Shouxi. Application of AMMI model in the stability analysis of spring wheat in rainfed areas [J]. Acta Ecol Sin, 2006, 26(11): 3677-3684. [6] 孙计平, 李雪君, 吴照辉, 等. 应用AMMI模型分析烤烟区试品种稳定性[J]. 中国农学通报, 2011, 27(19): 263-267. SUN Jiping, LI Xuejun, WU Zhaohui, et al. Analysis on the stability of flue-cured tobacco varieties in regional trail by AMMI model [J]. Chin Agric Sci Bull, 2011, 27(19): 263-267. [7] 王军辉, 顾万春, 李斌, 等. 桤木优良种源/家系的选择研究: 生长的适应性和遗传稳定性分析[J]. 林业科学, 2000, 36(3): 59-66. WANG Junhui, GU Wanchun, LI Bin, et al. Study on selection of Alnus cremastogyne provenance/family: analysis of growth adaptation and genetic stability [J]. Sci Silv Sin, 2000, 36(3): 59-66. [8] 高金锋. 荞麦品种稳定性与适应性分析及评价研究[D]. 杨凌: 西北农林科技大学, 2008. GAO Jinfeng. Analysis and Assessment of Stability and Adaptility of Buck Wheat Variety [D]. Yangling: Northwest A & F University, 2008. [9] 范士杰, 王蒂, 张俊莲, 等. 应用AMMI模型分析分析评判贵州省马铃薯区试品种的稳定性和适应性[J]. 种子, 2012, 31(7): 106-109. FAN Shijie, WANG Di, ZHANG Junlian, et al. Application of AMMI model to analysis stability and adaptability of potato in variety regional trial of Guizhou Province [J]. Seed, 2012, 31(7): 106-109. [10] SHA X Y, LINSCOMBE S D, GROTH D E, et al. Registration of ‘Della-2’ aromatic long-grain rice [J]. J Plant Registr, 2013, 7(3): 26-30. [11] STAMM M, BERRADA A, BUCK J, et al. Registration of ‘Riley’ winter canola [J]. J Plant Registr, 2012, 6(3): 243-245. [12] BURTON J W, MIRANDA L M, CARTER T E, et al. Registration of ‘NC-Miller’ soybean with high yield and high seed-oil content [J]. J Plant Registr, 2012, 6(3): 294-297. [13] 陈学宽, 刘家勇, 赵俊, 等. 甘蔗品种区域化试验基因与环境互作研究[J]. 中国糖料, 2013(3): 1-7. CHEN Xuekuan, LIU Jiayong, ZHAO Jun, et al. Research on gene-environment interactions in sugarcane regional trial in Yunnan [J]. Sugar Crops China, 2013(3): 1-7. [14] 刘娜娜. 长三角平原水网地区耐湿景观树种引种适应性评价与选择[J]. 中南林业科技大学学报, 2010, 30(8): 47-52. LIU Nana. Adaptability evaluation and selection of humidity tolerance and landscape tree species introduced from USA to Yangtze Delta Region [J]. J Cent South Univ For Technol, 2010, 30(8): 47-52. [15] 骆仁祥, 张春霞, 刘国华, 等. 南京地区园林绿化竹种生态适应性评价[J]. 林业科技开发, 2012, 26(3): 42-46. LUO Renxiang, ZHANG Chunxia, LIU Guohua, et al. Evaluation on ecological adaptation of landscaped bamboo species in Nanjing [J]. China For Sci Technol, 2012, 26(3): 42-46. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2016.05.015
 点击查看大图
点击查看大图
计量
- 文章访问数: 3817
- HTML全文浏览量: 716
- PDF下载量: 452
- 被引次数: 0
