






-
全球气候变暖已成为国际社会关注的焦点,而大量二氧化碳等温室气体排放形成的温室效应则是气候变暖的根源。森林是陆地上最大的生态系统,也是陆地生态系统的最大碳库,森林生态系统维持的碳库占全球总碳库的46.30%。森林碳汇功能具有比其他减排方式更经济和高效的的优点,《京都议定书》中明确森林碳汇是二氧化碳减排的主要替代方式[1-3]。碳汇造林不仅可以增加陆地碳汇,而且能减缓大气中二氧化碳的积累,具有重要的政治意义和科学实践意义[4]。竹林是地球陆地上重要的森林植被类型,也是特殊的生态系统单元。中国地处世界竹子分布的中心区,是世界第一大产竹国,有竹子39个属500余种,竹林面积占森林面积的3.06%[5]。竹林为异龄林,与其他林分相比,具有生长速度快、高产和用途广泛等优点,竹笋出土后在较短的时间内(40 d左右)完成高生长,1年生毛竹Phyllostachys edulis的碳储量在6个月时间内已占全年碳储量的88.80%以上,毛竹具有很强的固碳能力,是速生阶段杉木Cunninghamia lanceolata林的1.33倍、热带山地雨林的1.33倍、27年生杉木林的2.16倍[3]。新造毛竹林林分状态分为成林前和成林后2个阶段。成林后的毛竹林林分结构稳定,包括立竹密度、胸径、竹高和生物量等。许多学者利用地面调查和遥感监测等方式对成林后毛竹林分生物量、固碳能力进行了深入的研究[6-9],关于毛竹林林分结构与地形环境、经营方式的关系研究也有较多报道。有研究认为:人工栽植毛竹林成林稳定后在异质性环境中的分布更偏向泊松分布[10],而在近自然状态下毛竹林空间分布格局随年份发生动态变化[11]。还有研究发现:通过改善经营措施对于优化毛竹林径级结构、年龄结构和空间结构从而提高单位面积生产力有重要作用[12-14]。与成林后毛竹林研究相比,学者对成林前毛竹林的相关研究较少,同时一些关于新造毛竹林成林前的研究也主要聚焦在非空间结构上,陈婷等[16]研究表明:毛竹林在成林前与成林后非空间结构间差异较大。陈双林等[15]研究发现:新造毛竹林林分结构状况受到立地类型、母竹质量与初植密度和造林模式等多重影响。毛竹林不同阶段空间分布格局的研究有助于完善对该物种种群水平结构的定量描述[17],因此,对毛竹林成林前空间结构的研究变得更有意义。为此,本研究以2块不同栽植方式的毛竹碳汇林为对象,对新竹进行了连续监测,探究不同栽植方式对毛竹林自然成林过程中的空间分布格局特征的影响,为毛竹碳汇林营造与管理提供参考。
-
试验区位于浙江省临安市藻溪镇严家村严家山(30°10′29.7″N,119°29′55.6″E),属中亚热带季风气候,平均海拔为550 m,年平均气温为15.8 ℃,年平均降水量为1 500.0 mm,年平均无霜期为236.0 d。地形地貌为低山丘陵,土壤为红壤,pH 4.4。森林植被中,乔木层主要有萌蘖次生杉木以及少量的青冈Cyclobalanopsis glauca等。
-
2008年11月,在试验区通过母竹移栽方式新造2块毛竹碳汇林,其中一块采用匀栽方式栽植,栽植面积为15.47 hm2;另一块采用丛栽方式栽植,栽植面积为20.99 hm2。母竹选用标准:年龄为1~2年生,胸径3~6 cm,竹高3.0~3.5 m,鞭色鲜黄,鞭芽饱满,鞭根健全,竹节正常,无病虫害。匀栽方式毛竹林地栽植穴规格为1.2 m(长)×0.6 m(宽)×0.5 m(深),植竹600株·hm-2,株行距为4 m × 4 m;丛栽方式栽植穴为1.2 m(长)×0.5 m(宽)×0.5 m(深),采用品字形设计,栽植穴3个·栽植点-1,植竹675株·hm-2。在毛竹林幼林阶段,每年春季和初夏各中耕除草1次,造林后3 a内采用穴施法施用尿素300~450 kg·hm-2·a-1,除挖去退笋、败笋外,全部留笋长竹,不进行伐竹作业。在匀栽与丛栽2种不同栽植方式毛竹碳汇林中各随机布设20块20 m × 20 m固定标准地。
2009-2013年,每年对试验固定标准地内的新竹进行定位监测。用NTS-352全站仪(测距精度:3 mm+2×10-6 mm;测角精度2 s)测定每株新竹的基部三维坐标(X, Y, H)和竹高h,其中X和Y是平面直角坐标,H是海拔高度,h是竹子基部到竹梢顶部的垂直高度,并利用围尺测量新竹的胸径。
-
聚集指数是最近邻单株距离的平均值与随机分布下的期望平均距离之比,经Donnelly优化[18],公式为:
$$R=\frac{\frac{1}{N}\sum\limits_{i=1}^{N}{{{r}_{i}}}}{\frac{1}{2}\sqrt{\frac{F}{N}}+\frac{0.0514F}{N}+\frac{0.041F}{{{N}^{\frac{3}{2}}}}}。$$ (1) 式(1)中:R为聚集指数;ri为第i株毛竹到最近邻竹的距离(m);N为固定标准地内毛竹株数(株);F为固定标准地面积(m2);P为固定标准地周长(m)。
-
考虑边缘影响校正后的Ripley’s K(d)函数公式为:
$$\hat{K}\left( d \right)=A\sum\limits_{i=1}^{N}{\sum\limits_{j=1}^{N}{\frac{{{w}_{ij}}\left( d \right)}{{{N}^{2}}}\left( i\ne j \right)。}}$$ (2) 式(2)中:N为固定标准地毛竹株数(株);d为距离尺度;wij为以毛竹i为圆心;d为半径的圆在样地中的周长部分与圆周长之比的倒数,dij为毛竹i与j之间的距离(m),且dij≤d;A为固定标准地面积(m2)[19]。
BESAG等[20]提出用$\hat{L}\left( d \right)$取代$\hat{K}\left( d \right)$,并对$\hat{K}\left( d \right)$作开平方的线性变换,以保持方差稳定。在随机分布的假设下,期望值接近于0[21]。$\hat{L}\left( d \right)$公式为:
$$\hat{L}\left( d \right)=\sqrt{\frac{R\left( d \right)}{\pi }}-d。$$ (3) 本研究中通过比较$\hat{L}\left( d \right)$与d的关系来分析毛竹林空间分布格局类型。$\hat{L}\left( d \right)<0$,表示呈均匀分布$\hat{L}\left( d \right)=0$,表示呈随机分布;$\hat{L}\left( d \right)>0$,表示呈聚集分布。
然后对固定标准地内毛竹实际观测分布$\hat{L}\left( d \right)$曲线用蒙特-卡洛检验法进行显著性检验。在某个距离尺度d范围内,比较固定标准地毛竹实际观测分布$\hat{L}\left( d \right)$曲线与蒙特-卡洛检验法得出的置信区间所绘制的上下2条包迹线来判别毛竹林立竹空间分布格局。实际观测分布$\hat{L}\left( d \right)$曲线位于包迹线上测,竹林呈聚集分布;实际观测分布$\hat{L}\left( d \right)$曲线位于包迹线下测,竹林呈均匀分布;实际观测分布$\hat{L}\left( d \right)$曲线位于2条包迹线中间,竹林呈随机分布[19]。本研究蒙特-卡洛检验法模拟次数为200次,最大距离尺度为固定标准地最短边长的一半,即10 m[21]。
-
以匀栽毛竹林2013年份新竹为例,所选择标准地个数为16个(表 1),而16个标准地的新竹$\hat{L}\left( d \right)$曲线图均有所差异,故需建立标准选取该年份16张新竹$\hat{L}\left( d \right)$曲线图中最具代表性的一张作为结果图,用以说明2013年新竹在各距离尺度下的空间分布格局。选取标准如下:① 利用Ripley’s K(d)函数计算所得图统计该年份新竹在6个距离尺度(1,2,4,6,8,10 m)上呈聚集分布的图的张数,并计算所得张数占曲线图总张数(所选标准地个数)的比例,得到比值。② 根据统计学经验,若比值大于0.5,说明新竹在该尺度下呈聚集分布的标准地个数超过所选标准地个数(16个)的一半,表示新竹在该尺度上呈聚集分布;若比值小于0.5,表示新竹呈随机分布或均匀分布。由于本研究对象在所有尺度上均未显示有均匀分布趋势,故当比值小于0.5时,新竹在该尺度呈随机分布。③ 在所有$\hat{L}\left( d \right)$曲线图中选择符合各距离尺度下分布格局的图作为2013年新竹$\hat{L}\left( d \right)$曲线图。
使用ForStat 2.1软件、Matlab软件和SPSS 20.0进行试验数据计算。
-
由于母竹的栽植方式会对研究带来一定的干扰,所以匀栽与丛栽2种栽植方式标准地毛竹株数均不包含母竹株数。从表 1可以看出:不同栽植方式的毛竹林中相同年份的新竹、活立竹株数和聚集指数都有所差异;相同栽植方式的毛竹林新竹、活立竹的聚集指数随着时间的推移发生动态的变化。
表 1 不同栽植方式的毛竹林不同年份新竹和活立竹聚集指数与株数
Table 1. Different planting patterns and different years of new bamboo's and live bamboo's aggregation index and tree number in moso
年份 栽植方式 标准地个数/个 新竹株数/株 新竹聚集指数 活立竹株数/株 活立竹聚集指数 2009 匀栽 8 5±3aAB 0.18 ± 0.16 aA 5 ± 3 aA 0.18 ± 0.16 aA 丛栽 6 4±3aA 0.49 ± 0.07 bAB 4±3aA 0.49 ± 0.07 bAB 2010 匀栽 16 10 ± 5 aAC 0.35 ± 0.12 aA 13 ± 7 aB 0.35 ± 0.16 aB 丛栽 17 25 ± 15 bBC 0.46 ± 0.15 bAB 28 ± 16 bB 0.47 ± 0.13 bA 2011 匀栽 16 13 ± 7 aCD 0.50 ± 0.22 aB 24 ± 13 aC 0.40 ± 0.14 aB 丛栽 18 13 ± 11 aD 0.38 ± 0.19 aA 30 ± 25 bBC 0.45 ± 0.14 aA 2012 匀栽 18 10 ± 6 aAC 0.65 ± 0.18 aC 33 ± 18 aC 0.42 ± 0.13 aBC 丛栽 20 20 ± 14 bBD 0.52 ± 0.15 bB 59 ± 32 bC 0.49 ± 0.18 aA 2013 匀栽 16 17 ± 11 aD 0.63 ± 0.21 aBC 50 ± 28 aD 0.52 ± 0.11 aC 丛栽 20 31 ±14 bC 0.76 ± 0.15 bC 89 ± 45 bD 0.60 ± 0.19 aB 说明:同列不同小写字母仅表示相同年份不同栽植方式间差异显著,同列不同大写字母表示相同栽植方式不同年份间差异显著 (P < 0.05)。新竹株数、新竹聚集指数、活立竹株数、活立竹聚集指数为平均值±标准差。 -
从表 1可知:匀栽栽植方式毛竹林2009年至2013年的新竹聚集指数总体呈上升趋势,聚集指数均小于1,呈聚集分布。2009年新竹聚集指数最低为0.18,2012年聚集指数最高为0.65;新竹数也总体呈上升趋势,2012年略有下降,2013年新竹数达到最大值为17株。2009年与2010年,新竹聚集指数与株数差异不显著(P > 0.05),说明造林后前2 a新竹分布格局差异不大,新竹发育速度较为缓慢;而2010-2012年,聚集指数随着造林年限增加而增加,年份间聚集指数差异显著(P < 0.05),表明造林后第2年开始地下竹鞭逐渐扩展,新竹出笋位置离母竹渐远。2009-2012年,从新竹株数来看,年份间差异并不明显,直到2013年,新竹数才有显著(P < 0.05)提高。研究发现:当新竹株数增加时,聚集指数也相应增大,新竹聚集指数与新竹株数符合幂函数关系式(图 1),新竹株数(x)为自变量,林分新竹聚集指数为因变量(y),函数关系式为y=0.041 6x0.999 2, R2=0.761 6。

图 1 匀栽与丛栽毛竹林新竹聚集指数和新竹株数的关系
Figure 1. Relation of aggregation index and number of new bamboo in uniform planting and group-planting moso bamboo forest
丛栽栽植方式毛竹林中,2009-2011年,聚集指数逐年下降,2011-2013年逐年上升,2011年新竹聚集指数最低为0.38,2013年新竹聚集指数最高为0.76。这5 a中的新竹聚集指数均小于1,呈聚集分布。2009年与2010年,丛栽新竹聚集指数年份间差异性与匀栽竹林相同,造林后前2 a,聚集指数差异不显著。2011-2013年聚集指数具有显著性差异(P < 0.05),表明该阶段毛竹林正处于快速发育阶段。2009-2013年,年份间新竹株数差异显著(P < 0.05),2009年新竹株最少为4株,2010年新竹突然呈爆发式增长,达到25株,2011年新竹突然减少至13株,然后逐步增加,2013年新竹数最多为31株,表明丛栽毛竹林造林后5 a为毛竹林快速发育阶段。同时,丛栽新竹聚集指数与新竹株数符合二次函数关系式(图 1),新竹株数为自变量(x),新竹聚集指数为因变量(y),函数关系式为y=0.001 1-0.028 4x+0.588 1, R2=0.828 8。
由表 1可见:2009年2种栽植方式新竹株数差异不显著,而2010年与2009年新竹数差异明显,表明通过母竹移栽方式进行毛竹造林后第1年,栽植方式不会影响新竹出土数量,可能的原因是新造竹林第1年母竹主要以扩展地下竹鞭为主,竹鞭尚未有出笋能力。造林后第2年开始(2010-2013年),相同年份匀栽与丛栽新竹聚集指数差异显著(P < 0.05)(除2012年),丛栽新竹株数显著(P < 0.05)高于匀栽(除2012年)。表明栽植方式对新竹的发育与分布格局的影响逐渐增大。
-
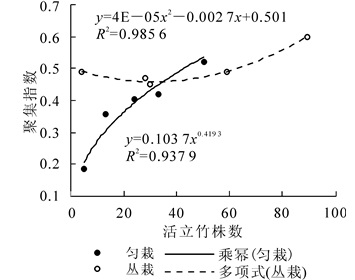
匀栽新造毛竹碳汇林造林第2年后,各年份间活立竹聚集指数差异不显著,聚集指数随着造林年限的增加呈缓慢上升趋势。2009-2013年,各年份间活立竹株数差异性显著(P < 0.05)。截至2013年,虽然每年活立竹株数都在增加,但是毛竹林空间分布格局仍然呈聚集分布,表明活立竹分布位置主要以靠近母竹为主,毛竹林还未成林,仍然处于发育阶段。活立竹聚集指数变动范围为0.18~0.52,林分活立竹株数的波动范围为5~50株。匀栽毛竹林聚集指数与活立竹株数关系见图 2。图 2中,活立竹株数为自变量(x),林分活竹聚集指数为因变量(y),拟合方程为y=0.103 7x0.419 3, R2=0.937 9。

图 2 匀栽与丛栽毛竹林聚集指数与活立竹株数的关系
Figure 2. Relation of aggregation index and number of stand live bamboo in uniform planting and groupplanting moso bamboo forest
丛栽栽植方式毛竹林中,2009-2011年活立竹聚集指数逐年下降,2011年聚集指数最低为0.45,随后聚集指数逐年递增,2013年聚集指数最大为0.60,该毛竹林分布格局仍呈聚集分布。丛栽毛竹林活立竹株数与聚集指数年份间差异均不显著,与匀栽竹林相同。可见,毛竹造林后5 a内,丛栽竹林也处于成林初期阶段。活立竹聚集指数变动范围为0.45~0.60,活立竹株数变动范围为4~89株。由图 2可见:活立竹聚集指数(y)和株数(x)的函数关系式满足y=0.000 04x2-0.002 7x+0.501,R2=0.985 6。
2009-2013年5 a造林年限中,对比2种栽植方式新造毛竹林可以发现,造林后前2 a,丛栽活立竹聚集指数显著(P < 0.05)高于匀栽,第3年(2011年)开始无显著差异。而在活立竹株数方面,除2009年外,丛栽竹林均显著(P < 0.05)高于匀栽。从林业生产角度出发,相同年份下丛栽聚集指数与活立竹株数均高于匀栽,因此,丛栽栽植方式要优于匀栽栽植方式,并且能够缩短成林所需时间。到目前为止,对比成熟毛竹林分[22],2种栽植方式下的新造毛竹碳汇林仍然处于成林初期阶段。
-
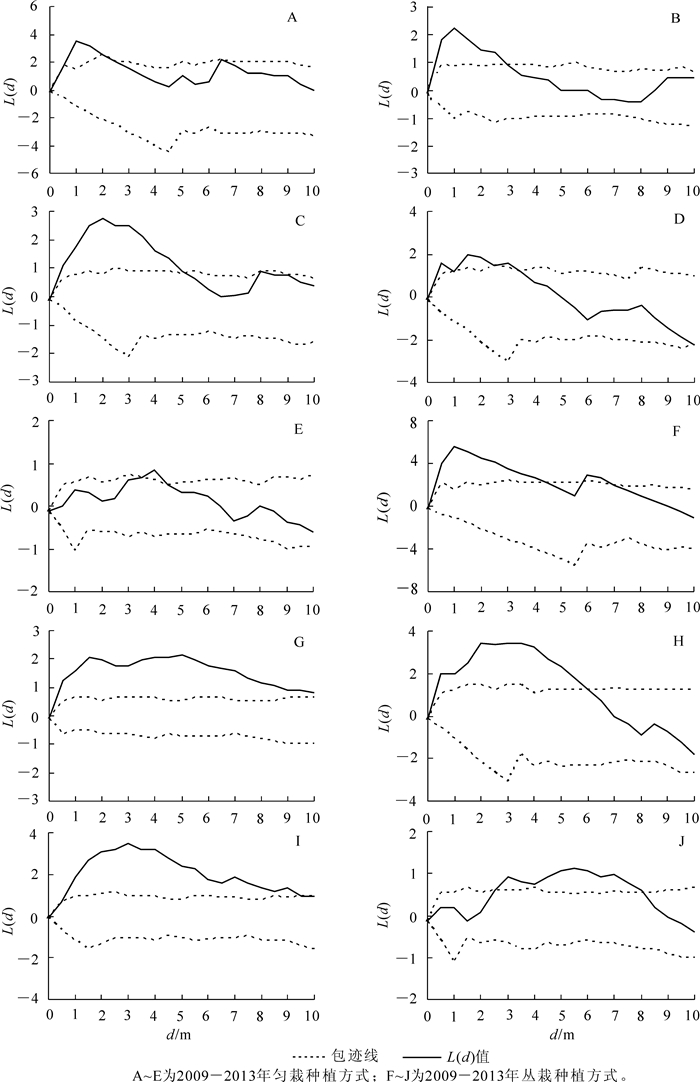
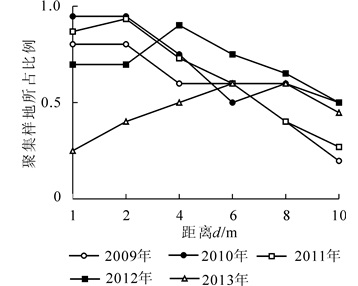
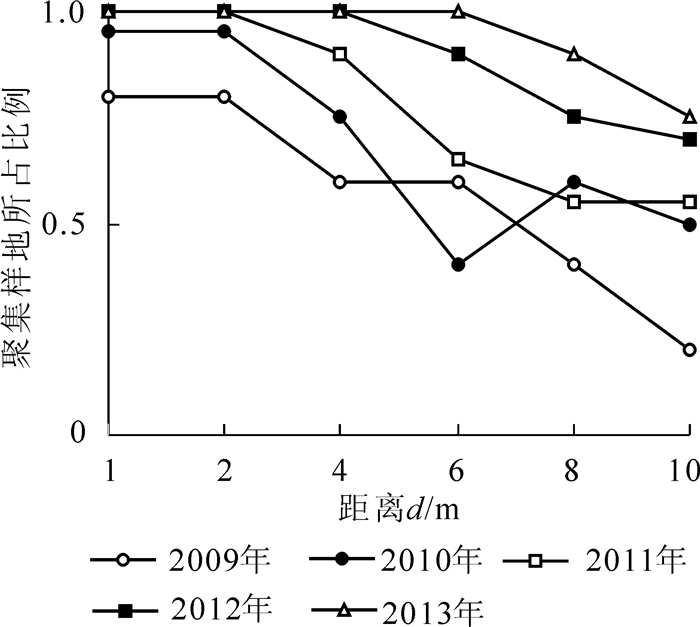
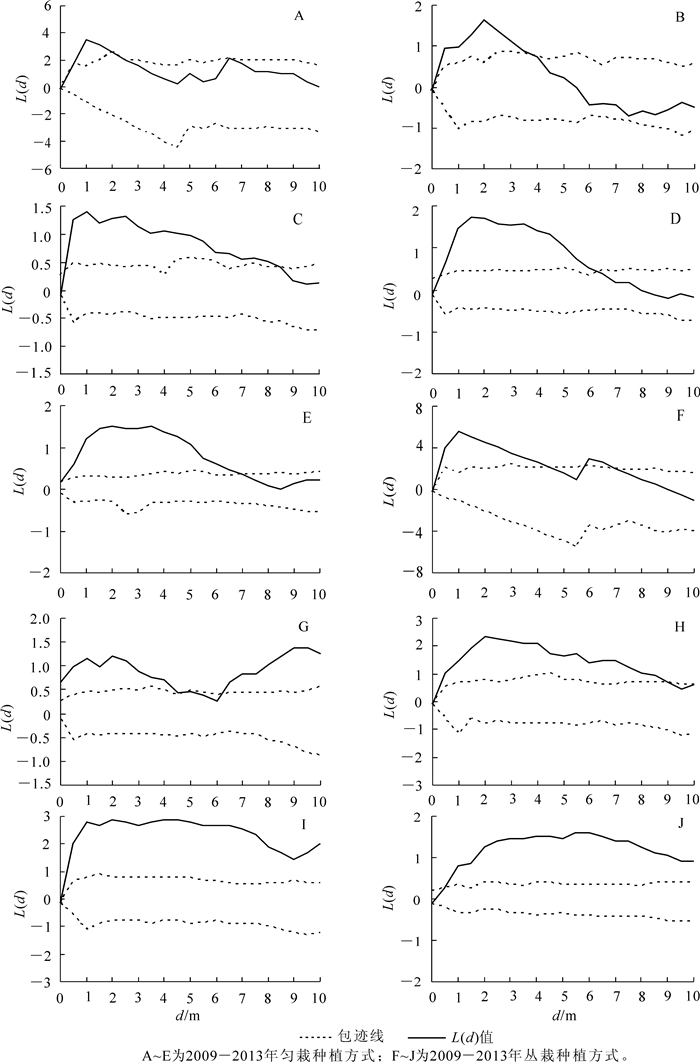
根据1.3.3小节介绍,由图 3中可见:2013年匀栽毛竹林新竹在4 m尺度下呈聚集分布,在1,2,6,8,10 m尺度下呈随机分布,图 4E为按照1.3.3小节标准选择的2013年匀栽毛竹林新竹$\hat{L}\left( d \right)$曲线图,符合新竹在4 m尺度下呈聚集分布,在1,2,6,8,10 m尺度下呈随机分布。以此类推,通过图 3,图 5,图 6和图 7所示新竹与活立竹在匀栽与丛栽毛竹林各距离尺度下的分布格局,得到匀栽与丛栽毛竹林各年份新竹、活立竹空间分布格局结果图(图 4,图 8)。

图 3 匀栽毛竹林新竹各距离尺度聚集标准地所占比例
Figure 3. Proportion of aggregated sample plots of new bamboo in all distance dimension in uniform planting moso bamboo forest

图 4 匀栽与丛栽毛竹林各年份新竹空间分布格局
Figure 4. Different years of stand new bamboo's spatial pattern in uniform planting moso bamboo forest and group-planting moso bamboo forest

图 5 匀栽毛竹林活立竹各距离尺度聚集标准地所占比例
Figure 5. Proportion of aggregated sample plots of live bamboo in all distance dimension in uniform planting moso bamboo forest

图 6 丛栽毛竹林新竹各距离尺度聚集标准地所占比例
Figure 6. Proportion of aggregated sample plots of new bamboo in all distance dimension in group-planting moso bamboo forest

图 7 丛栽毛竹林活立竹各距离尺度聚集标准地所占比例
Figure 7. Proportion of aggregated sample plots of live bamboo in all distance dimension in group-planting moso bamboo forest

图 8 匀栽与丛栽毛竹林各年份活立竹空间分布格局
Figure 8. Different years of stand live bamboo's spatial pattern in uniform planting moso bamboo forest and group-planting moso bamboo forest
-
图 4 A~E分别是匀栽栽植方式2009-2013年新竹空间分布格局图。匀栽的新造毛竹碳汇林2009年的新竹在0~2.2 m的尺度下,呈聚集分布,在2.2 m尺度以上呈随机分布;2010年新竹在0~3.3 m的尺度下呈聚集分布,在3.3 m尺度以上呈随机分布;2011年新竹在5.4 m尺度以上呈随机分布。新竹的繁殖离不开母竹竹鞭的扩展,新竹出笋位置离母竹较近,因此新竹小尺度呈聚集分布,大尺度呈随机分布表明造林前几年新竹分布格局受到了母竹栽植方式的影响。随着造林年限的增加,新竹呈随机分布的距离尺度逐渐变远,表明母竹竹鞭的长度逐渐增加,新竹出土位置离母竹渐远。2012-2013年,新竹逐渐在各距离尺度上呈随机分布表明毛竹林地下鞭根系统逐渐完善。图 4A~E中的波谷尖峰所在距离表明新竹在该尺度下存在团状分布,波谷增多说明新竹的团状分布个数增多,表明竹林地下鞭根出现交错现象,致使部分不同种源新竹在一定尺度下呈现团状聚集分布。图 4F~J分别是丛栽栽植方式2009-2013年新竹空间分布格局图,丛栽栽植方式2009年新竹在距离尺度大于4.4 m以上呈随机分布也受到了母竹栽植方式的影响。但2010与2012年新竹整体分布格局主要呈聚集分布,在0~3.0 m距离尺度上新竹聚集趋势明显,但在距离尺度大于5.0 m以上,聚集趋势开始减弱。2013年新竹聚集趋势进一步降低,以随机分布为主要分布格局。
-
匀栽毛竹林2009年活立竹在0~2.2 m的尺度下呈聚集分布,在2.2 m尺度以上呈随机分布(图 8A)。2010年林分活立竹从距离尺度4.0 m以下的聚集分布过渡到距离尺度4.0 m以上的随机分布,在距离尺度为6.0 m处有一个波谷尖峰(图 8B)。2011年在距离尺度为1.0, 3.5, 6.0 m处有波谷尖峰,表明活立竹在该尺度下存在团状分布(图 8C)。2012年活立竹在距离尺度4.0 m以上聚集趋势减弱,在距离尺度为9.0 m处有1个波谷尖峰(图 8D),至2013年该波谷尖峰变得明显(图 8E)。丛栽毛竹林2010年活立竹在尺度距离6.0 m处有明显波谷尖峰(图 8G),2011年活立竹在6.0, 9.4 m距离尺度下有明显尖峰(图 8H),2012年活立竹在9.0 m距离尺度下团状分布现象更加明显(图 8I),至2013年活立竹在固定尺度上的尖峰逐渐消失,$\hat{L}\left( d \right)$估计值曲线波动变小趋于稳定,说明新造毛竹碳汇林地下鞭根系统逐渐完善(图 8J)。图 8C~E发现匀栽方式的毛竹林在9.0 m距离尺度下的尖峰没有丛栽方式那么明显,出现该现象的时间比丛栽方式迟1 a。随着造林年限的增加,不管是匀栽、丛栽栽植方式下的毛竹林,活立竹在固定尺度下的团状分布会逐渐消失,在0~2.0 m尺度下聚集趋势明显,在2.0~10.0 m尺度下聚集趋势开始减弱。
-
本研究在新造毛竹碳汇林设置固定标准地,并对标准地中新竹坐标等连续监测,利用聚集指数和Ripley’s K(d)函数2种方法对2009-2013年不同栽植方式下新造毛竹碳汇林成林初期空间分布格局进行研究,得出以下结论:① 5 a造林年限间,匀栽与丛栽毛竹林均呈聚集分布。② 新造毛竹碳汇林成林初期,新竹分布格局受母竹栽植方式影响明显。③ 不同栽植方式下的毛竹碳汇林,活立竹株数与聚集指数之间存在显著相关性。匀栽毛竹林活立竹株数(x)与聚集指数(y)拟合方程为y=0.103 7x0.419 3, R2=0.937 9。丛栽毛竹林活立竹株数(x)与聚集指数(y)符合二次方程:y=0.000 04x2-0.002 7x+0.501, R2=0.985 6。④ 新造毛竹碳汇林在成林过程中存在不同距离尺度下的团状聚集分布。⑤ 栽植方式对新竹与活立竹株数和分布格局影响明显。
本研究发现2009-2010年间,匀栽栽植方式下新竹存在小尺度聚集(0~3 m),大尺度(6~10 m)随机分布。范辉华[23]研究认为:新造竹林造林当年竹鞭扩展范围为1.934 m。因此,新竹在6~10 m距离尺度上呈随机分布是受到了母竹栽植方式的影响。丛栽栽植方式下2009-2010年新竹株数增加,而聚集指数2009-2010年下降,表明新竹的分布格局也同样受到了母竹栽植方式的影响。
造林后4 a(2009-2012年),匀栽与丛栽竹林内新竹与活立竹随着造林年限增加相继在距离尺度为1.5, 6.1, 9.0 m处出现团状聚集分布。这种现象与毛竹该物种通过地下鞭繁殖有着密切的关系,新造毛竹林造林3 a地下竹鞭最大生长长度可达6.18 m[24],正因地下鞭根系统的扩展,部分新竹出笋位置离母竹越来越远,致使毛竹团状分布的距离尺度随着造林年限的增加而增大。造林年限的增加(2012-2013年)小尺度团状分布(d=1.5 m)因为地下竹鞭的完善而消失,竹林分布格局趋于稳定。同时还发现,毛竹林各距离尺度下的团状聚集分布在一定程度上可以反映地下竹鞭的生长蔓延情况,为研究毛竹林地下竹鞭的生长蔓延状况提供了一种新的思路。
本研究利用聚集指数对试验数据计算发现,栽植方式对新竹、活立竹株数与聚集指数影响显著。毛竹造林后第1年,匀栽与丛栽竹林在新竹株数上无显著差异,可能的原因是母竹移栽时只保留了30~50 cm的地下鞭,造林第1年母竹主要以竹鞭发育为主,造成第1年新竹出土数量较少。从第2年开始,丛栽竹林新竹与活立竹株数便显著高于匀栽竹林,新竹的聚集指数也显著高于匀栽竹林。不仅如此,利用Ripley’s K(d)函数对试验数据的研究发现,匀栽竹林在距离尺度9.0 m出现明显波谷尖峰较丛栽方式竹林晚1 a,截至2013年,丛栽竹林新竹与活立竹的$\hat{L}\left( d \right)$曲线较之匀栽竹林更为平滑,表明丛栽竹林内各距离尺度下团状分布逐渐减少,林分分布格局相对于匀栽竹林更为稳定。从林业生产角度出发,丛栽栽植方式相对于匀栽方式更利于新竹数量上的扩展,对新造竹林的发育起到了积极的作用,加快了毛竹林成林进程,缩短了成林时间。造成这些现象的原因可能是:匀栽栽植方式母竹行距要小于丛栽栽植方式。丛栽方式母竹3株呈品字形为1个栽植点,虽然3株母竹会产生竞争,但是毛竹为单轴散生型竹种,合理改变栽植时母竹竹鞭和鞭芽的方向,使得3株母竹竹鞭在3个不同方向上生长,而且栽植点行距较远,在造林初期该方法能使母竹更好地利用周围的空间和土壤养分。
在种群发育过程中,分布格局是随着时间的推移而表现出动态变化的过程[25]。本研究采用时间尺度与空间尺度相结合的方法,设置不同栽植方式下的毛竹林地,能更好地反映竹林在成林初期空间分布格局的动态变化。竹林碳汇是森林碳汇中一个重要的组成部分,因此,毛竹的造林方式将对碳汇造林起到重要影响,为毛竹林以及碳汇林的营造和经营提供了科学的依据。同时,对毛竹林成林初期以及成林后格局的分析,对森林碳汇计量与监测以及增汇技术的提高提供科学的帮助。由于新造竹林到达成熟稳定阶段一般需要6~7 a或者更长的时间[15],笔者还将继续对现阶段以及成林稳定后的2种不同栽植方式下的毛竹林进行监测,完善更长时间尺度的在自然状态下竹林从营造到成林特征变化的研究。
Planting pattern's influence for characteristics of variation of spatial distribution pattern in the early stages of moso bamboo carbon sink stands
-
摘要: 毛竹Phyllostachys edulis林空间结构是毛竹生长过程的重要因子,是提高竹林质量,优化经营水平的重要参考依据。利用聚集指数和Ripley's K(d)函数,分析从2009-2013年毛竹碳汇林成林初期空间分布格局年际变化特征及匀栽、丛栽栽植方式对分布格局的影响。结果表明:① 在5 a造林年限间,2种栽植方式下的新竹与活立竹的空间分布格局均呈聚集分布;② 丛栽栽植毛竹林新竹聚集指数变动范围为0.38~0.76,活立竹聚集指数变动范围为0.45~0.60;匀栽栽植毛竹林新竹聚集指数变动范围为0.18~0.65,活立竹聚集指数变动范围为0.18~0.52;③ 聚集指数与活立竹株数间存在较强的相关性,匀栽毛竹林活立竹株数(x)与聚集指数(y)拟合方程为y=0.1037x0.4193,R2=0.9379;丛栽毛竹林活立竹株数(x)与聚集指数(y)符合二次方程:y=0.00004x2-0.0027x+0.501,R2=0.9856;④ 新造毛竹碳汇林在成林过程中存在不同尺度距离下的团状聚集分布;⑤ 栽植方式对立竹株数和分布格局影响明显。Abstract: Spatial structure in a moso bamboo (Phyllostachys edulis) stand is an important factor in its growing process. To analyze the influence of interannual variation characteristics of spatial patterns having different planting patterns in the early stage of moso bamboo stands used as carbon sinks, an aggregation index and Ripley's K(d) function were applied from 2009 to 2013, and results were fitted with regression equations. Results showed 1) spatial patterns of new moso bamboo and established moso bamboo with an aggregated distribution pattern after five years of afforestation. 2) The aggregation index in group-planted moso bamboo stands for new moso bamboo was from 0.38 to 0.76 and for established moso bamboo was from 0.45 to 0.60; for uniformly planted moso bamboo stands, the aggregation index for new moso bamboo was from 0.18 to 0.65, and the established moso bamboo stand was from 0.18 to 0.52. 3) A strong relationship was found between the aggregation index of an established stand of moso bamboo (y) and culm number (x). For a uniformly planted moso bamboo stand, the fitted regression equation was y=0.103 7x0.419 3, R2=0.937 9; and for group-planted moso bamboo stands it was y=0.000 04x2-0.002 7x + 0.501, R2=0.985 6. 4) In the process of forest establishment, new moso bamboo carbon sink stands existed as an agglomerate distribution for different distance scales. 5) Thus, the cultivation mode influenced the moso bamboo stand's culm number and spatial distribution pattern.
-
图 1 匀栽与丛栽毛竹林新竹聚集指数和新竹株数的关系
Figure 1 Relation of aggregation index and number of new bamboo in uniform planting and group-planting moso bamboo forest
图 2 匀栽与丛栽毛竹林聚集指数与活立竹株数的关系
Figure 2 Relation of aggregation index and number of stand live bamboo in uniform planting and groupplanting moso bamboo forest
图 3 匀栽毛竹林新竹各距离尺度聚集标准地所占比例
Figure 3 Proportion of aggregated sample plots of new bamboo in all distance dimension in uniform planting moso bamboo forest
图 4 匀栽与丛栽毛竹林各年份新竹空间分布格局
Figure 4 Different years of stand new bamboo's spatial pattern in uniform planting moso bamboo forest and group-planting moso bamboo forest
图 5 匀栽毛竹林活立竹各距离尺度聚集标准地所占比例
Figure 5 Proportion of aggregated sample plots of live bamboo in all distance dimension in uniform planting moso bamboo forest
图 6 丛栽毛竹林新竹各距离尺度聚集标准地所占比例
Figure 6 Proportion of aggregated sample plots of new bamboo in all distance dimension in group-planting moso bamboo forest
图 7 丛栽毛竹林活立竹各距离尺度聚集标准地所占比例
Figure 7 Proportion of aggregated sample plots of live bamboo in all distance dimension in group-planting moso bamboo forest
图 8 匀栽与丛栽毛竹林各年份活立竹空间分布格局
Figure 8 Different years of stand live bamboo's spatial pattern in uniform planting moso bamboo forest and group-planting moso bamboo forest
表 1 不同栽植方式的毛竹林不同年份新竹和活立竹聚集指数与株数
Table 1. Different planting patterns and different years of new bamboo's and live bamboo's aggregation index and tree number in moso
年份 栽植方式 标准地个数/个 新竹株数/株 新竹聚集指数 活立竹株数/株 活立竹聚集指数 2009 匀栽 8 5±3aAB 0.18 ± 0.16 aA 5 ± 3 aA 0.18 ± 0.16 aA 丛栽 6 4±3aA 0.49 ± 0.07 bAB 4±3aA 0.49 ± 0.07 bAB 2010 匀栽 16 10 ± 5 aAC 0.35 ± 0.12 aA 13 ± 7 aB 0.35 ± 0.16 aB 丛栽 17 25 ± 15 bBC 0.46 ± 0.15 bAB 28 ± 16 bB 0.47 ± 0.13 bA 2011 匀栽 16 13 ± 7 aCD 0.50 ± 0.22 aB 24 ± 13 aC 0.40 ± 0.14 aB 丛栽 18 13 ± 11 aD 0.38 ± 0.19 aA 30 ± 25 bBC 0.45 ± 0.14 aA 2012 匀栽 18 10 ± 6 aAC 0.65 ± 0.18 aC 33 ± 18 aC 0.42 ± 0.13 aBC 丛栽 20 20 ± 14 bBD 0.52 ± 0.15 bB 59 ± 32 bC 0.49 ± 0.18 aA 2013 匀栽 16 17 ± 11 aD 0.63 ± 0.21 aBC 50 ± 28 aD 0.52 ± 0.11 aC 丛栽 20 31 ±14 bC 0.76 ± 0.15 bC 89 ± 45 bD 0.60 ± 0.19 aB 说明:同列不同小写字母仅表示相同年份不同栽植方式间差异显著,同列不同大写字母表示相同栽植方式不同年份间差异显著 (P < 0.05)。新竹株数、新竹聚集指数、活立竹株数、活立竹聚集指数为平均值±标准差。  下载: 导出CSV
下载: 导出CSV
-
[1] 何英, 张小全, 刘云仙.中国森林碳汇交易市场现状与潜力[J].林业科学, 2007, 43(7):106-111. HE Ying, ZHANG Xiaoquan, LIU Yunxian. Present status and potentiality of forest carbon trade market in China[J]. Sci Silv Sin, 2007, 43(7):106-111. [2] 李海奎, 雷渊才, 曾伟生.基于森林清查资料的中国森林植被碳储量[J].林业科学, 2011, 47(7):7-12. LI Haikui, LEI Yuancai, ZENG Weisheng. Forest carbon storage in China estimated using forestry inventory data[J]. Sci Silv Sin, 2011, 47(7):7-12. [3] 陈茂铨, 金晓春, 吴林森, 等.竹林碳汇功能及其影响因子研究进展[J].竹子研究汇刊, 2010, 29(3):5-9. CHEN Maoquan, JIN Xiaochun, WU Linsen, et al. Research progress of carbon sink and its influencing factors of bamboo forest[J]. J Bamboo Res, 2010, 29(3):5-9. [4] 史军, 刘纪远, 高志强, 等.造林对陆地碳汇影响的研究进展[J].地理科学进展, 2004, 23(2):58-67. SHI Jun, LIU Jiyuan, GAO Zhiqiang, et al. Research advances of the influence of afforestation on terrestrial carbon sink[J]. Prog Geogr, 2004, 23(2):58-67. [5] 陈先刚, 张一平, 张小全, 等.过去50年中国竹林碳储量变化[J].生态学报, 2008, 28(11):5218-5227. CHEN Xiangang, ZHANG Yiping, ZHANG Xiaoquan, et al. Carbon stock changes in bamboo stands in China over the last 50 years[J]. Acta Ecol Sin, 2008, 28(11):5218-5227. [6] DU Huaqiang, CUI Ruirui, ZHOU Guomo, et al. The responses of moso bamboo (Phyllostachys heterocycla var. pubescens) forest aboveground biomass to Landsat TM spectral reflectance and NDVI[J]. Acta Ecol Sin, 2010, 30(5):257-263. [7] ZHOU Guomo, MENG Cifu, JIANG Peikun, et al. Review of carbon fixation in bamboo forests in China[J]. Bot Rev, 2011, 77(3):262-270. [8] YEN T M, JI Y J, LEE J S. Estimating biomass production and carbon storage for a fast-growing makino bamboo (Phyllostachys makinoi) plant based on the diameter distribution model[J]. For Ecol Manage, 2010, 260(3):339-344. [9] YEN T M, LEE J S. Comparing aboveground carbon sequestration between moso bamboo (Phyllostachys heterocycla) and China fir (Cunninghamia lanceolata) forests based on the allometric model[J]. For Ecol Manage, 2011, 261(6):995-1002. [10] 时培建, 郭世权, 杨清培, 等.毛竹的异质性空间点格局分析[J].生态学报, 2010, 30(16):4401-4407. SHI Peijian, GUO Shiquan, YANG Qingpei, et al. Inhomogeneous spatial point pattern analysis of moso bamboo (Phyllostachys edulis)[J]. Acta Ecol Sin, 2010, 30(16):4401-4407. [11] 仇建习, 汤孟平, 沈利芬, 等.近自然毛竹林空间结构动态变化[J].生态学报, 2014, 34(6):1444-1450. QIU Jianxi, TANG Mengping, SHEN Lifen, et al. Dynamic analysis of spatial structure in a close-to-nature Phyllostachys edulis stands[J]. Acta Ecol Sin, 2014, 34(6):1444-1450. [12] 汤孟平, 徐文兵, 陈永刚, 等.毛竹林空间结构优化调控模型[J].林业科学, 2013, 49(1):120-125. TANG Mengping, XU Wenbing, CHEN Yonggang, et al. Spatial structure optimizing adjustment and control model of Phyllostachys edulis stand[J]. Sci Silv Sin, 2013, 49(1):120-125. [13] 刘恩斌, 施拥军, 李永夫, 等.浙江毛竹林分非空间结构特征及其动态变化[J].林业科学, 2013, 49(9):1-7. LIU Enbing, SHI Yongjun, LI Yongfu, et al. Non spatial structural characteristic of moso bamboo forest and its dynamics in Zhejiang Province[J]. Sci Silv Sin, 2013, 49(9):1-7. [14] 汤孟平, 徐文兵, 陈永刚, 等.天目山近自然毛竹林空间结构与生物量的关系[J].林业科学, 2011, 47(8):1-6. TANG Mengping, XU Wenbing, CHEN Yonggang, et al. Relationship between spatial structure and biomass of a close-to-nature Phyllostachys edulis stand in Tianmu Mountain[J]. Sci Silv Sin, 2011, 47(8):1-6. [15] 陈双林, 吴柏林, 吴明, 等.新造毛竹林林分结构年际演替规律及影响因子[J].浙江林学院学报, 2004, 21(4):393-397. CHEN Shuanglin, WU Bolin, WU Ming, et al. A study of the interannual succession rule and influential factors of young stands structures of Phyllostachys pubescens[J]. J Zhejiang For Coll, 2004, 21(4):393-397. [16] 陈婷, 施拥军, 周国模, 等.毛竹碳汇林营造初期林分非空间结构年际变化特征[J].浙江农林大学学报, 2015, 32(2):181-187. CHEN Ting, SHI Yongjun, ZHOU Guomo, et al. Interannual variation characteristics for stand structures in the early stages of new moso bamboo carbon sink stands[J]. J Zhejiang A & F Univ, 2015, 32(2):181-187. [17] 惠刚盈, 胡艳波, 徐海.森林空间结构的量化分析方法[J].东北林业大学学报, 2005, 33(增刊):45-48. HUI Gangying, HU Yanbo, XU Hai. Quantitative analysis of forest spatial structure[J]. J Northeast For Univ, 2005, 33(supp):44-48, 60. [18] DONNELLY K P, HOLDNER I. Simulations to Determine the Variance and Edge Effect of Total Nearest-neighour Distance[M]. London:Cambridge University Press:1978:91-95. [19] 汤孟平.森林空间结构分析[M].北京:科学出版社, 2013.. [20] BESAG J L, DIGGLE P J. Simple Monte Carlo tests for spatial pattern[J]. Appl Stat, 1977, 26(3):327-333. [21] 汤孟平, 周国模, 施拥军, 等.天目山常绿阔叶林优势种群及其空间分布格局[J].植物生态学报, 2006, 30(5):743-752. TANG Mengping, ZHOU Guomo, SHI Yongjun, et al. Study of dominant plant populations and their spatial patterns in evergreen broadleaved forest in Tianmu mountain[J]. J Plant Ecol, 2006, 30(5):743-752. [22] 邓英英, 汤孟平, 徐文兵, 等.天目山近自然毛竹纯林空间分布格局研究[J].竹子研究汇刊, 2011, 30(1):11-15. DENG Yingying, TANG Mengping, XU Wenbing, et al. A study on spatial patterns of nature-approaching moso bamboo pure forest in Tianmu Mountain[J]. J Bamboo Res, 2011, 30(1):11-15. [23] 范辉华.新造毛竹林竹鞭生长规律的研究[J].福建林学院学报, 1999, 19(1):30-32. FAN Huihua. Study on rhizome growth regularity of new-planted Phyllostachys edulis[J]. J Fujian Coll For, 1999, 19(1):30-32. [24] 邱尔发, 陈存及, 范辉华, 等.毛竹种源竹鞭生长进程研究[J].江西农业大学学报, 2002, 24(2):245-250. QIU Erfa, CHEN Cunji, FAN Huihua, et al. A study on the rhizome growth process of provenances of Phyllostachys pubescens[J]. Acta Agric Univ Jiangxi, 2002, 24(2):245-250. [25] 操国兴, 钟章成, 刘芸, 等.缙云山川鄂连蕊茶种群空间分布格局研究[J].生物学杂志, 2003, 20(1):10-12. CAO Guoxing, ZHONG Zhangcheng, LIU Yun, et al. The study of distribution pattern of Camellia rosthorniana population in Jinyun mountain[J]. J Biol, 2003, 20(1):10-12. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2017.03.003
 点击查看大图
点击查看大图

计量
- 文章访问数: 3004
- HTML全文浏览量: 700
- PDF下载量: 506
- 被引次数: 0
