




-
森林植被分布信息对于森林资源清查、科学管护、林火监测预警、林业航空植保等有重要作用[1-2]。传统野外调研手段获取森林植被分布信息,需投入大量的人力、物力和财力,费工费时,遥感技术快速提取森林植被空间分布信息可为其提供支撑,具有很大的实际应用价值。现今卫星遥感影像的空间分辨率从0.3 m(WorldView-3等)到数十千米(FY-3等),数据丰富、类型多样,很多高中低空间分辨率影像已被应用于森林植被信息提取[3-4]。国外高分辨率卫星影像价格昂贵,随着高分辨率卫星技术的发展,涌现出GF-1,GF-2和资源三号等一大批国产高分辨率数据,大大降低了数据获取成本,其中GF-1卫星的16.0 m影像可免费使用。目前,森林植被遥感监测时,大多凭以往研究生产经验来判断和筛选影像,然而森林植被信息提取涉及遥感影像量化分析。如何从众多遥感影像中通过科学判断来确定适合森林植被提取的最佳分辨率影像是森林植被信息提取中面临的一个问题。尽管目前已有一些学者研究林地遥感监测尺度[5-11],但大多利用国外卫星(IKONOS,Landsat等)数据,且主要侧重于最佳尺度选择方法理论研究,较少考虑林业实际应用。随着新近高分辨率卫星影像数据业务化获取及民用的推广,森林植被监测已成为主要应用领域之一,但是林业生产者或林业遥感用户从森林植被遥感监测精度、成本和效率等因素考虑,如何选择和确定适合森林植被监测的国产高分辨率影像最佳分辨率的知识缺乏,有待针对林业生产实际开展深入研究。在理想情况下,选取合适空间分辨率的标准是选取包含所需信息且数据量最小的空间分辨率[12]。目前,确定最佳尺度的方法主要有3类:①基于局部方差的方法[5, 13],②基于离散度的方法[14],③基于变异函数的方法[15-16]。局部方差法存在边界效应影响精度,离散度法未考虑到影像的空间分布特征。柏延臣等[3]提出了基于一个步长的变异函数法。该方法的优点是没有局部方差方法中的边界效应问题,也不需要通过复杂的计算过程去获取任意分辨率影像的半方差,计算量小,可方便快速地确定最优影像分辨率[3, 17]。本研究针对森林植被遥感监测实际应用需要,采用基于1个步长的变异函数法,计算GF-2全色影像变异函数,初步确定适合森林植被监测的影像最低空间分辨率;并对比不同尺度影像的森林植被提取精度,综合考虑成本和效率,最终确定适合森林植被监测影像的最优空间分辨率。
-
研究区位于湖南省衡阳市常宁市洋泉镇东侧林区(26°15′49″~ 26°17′07″N,112°15′05″~112°16′32″E),海拔为120~300 m。洋泉镇位于常宁市西南边陲,属中亚热带季风湿润气候,四季分明,地形较复杂。地势南高北低,西南区域为山地,东部有洋泉水库,北部区域主要为城镇及村庄。研究区地物类型丰富,主要为森林植被、草地、建筑、道路、水体等。森林植被分布广泛且类型多样,大面积原始山体林、中等面积人工林及小面积行道树、城镇内绿化林等交错分布。
-
以研究区2015年9月17日GF-2卫星影像为基础数据。其中1个全色波段,影像空间分辨率为0.8 m;多光谱影像有4个波段,分别为红波段、绿波段、蓝波段和近红外波段,影像空间分辨率为3.2 m。野外调研所得的森林植被分布结果用于辅助分析。
-
利用遥感专业软件ENVI对GF-2遥感影像进行辐射校正、大气校正、正射校正、影像融合、影像裁剪等一系列预处理,得到本研究所需的全色和多光谱融合影像数据,空间分辨率均为0.8 m,影像大小为3 000 × 3 000(像元)。参考目前常用全色或多光谱影像(GF-1,GF-2,Landsat)的空间分辨率情况,利用最近邻算法对融合后的0.8 m影像进行重采样,降低空间分辨率,得到2.0,3.2,8.0,16.0,30.0 m多尺度影像集。从森林植被的冠幅大小来看,常见的空间分布类型有3种:大冠幅、小冠幅和大小冠幅混合(图 1)。由于森林植被的冠幅大小直接影响着适合其遥感提取的影像最佳分辨率,因此,结合研究区实际情况,选择3种典型的森林植被类型(大冠幅森林植被、小冠幅森林植被和混合冠幅森林植被),利用全色影像进行变异函数尺度实验。对全色影像数据进行裁剪,得到3种典型森林植被影像,影像大小为200 × 200(像元)。

图 1 研究区位置及3种典型森林植被分布图
Figure 1. Location of the study area and distribution map of three typical forest vegetation
-
利用变异函数法初步确定适合森林植被监测的影像最低空间分辨率,并对比不同尺度影像中森林植被提取精度进行评价,综合考虑成本和效率等确定适合森林植被监测的影像最优空间分辨率。
-
遥感影像的像素是与空间位置紧密相关的函数,属于区域化变量。近些年,随着地统计学(以区域化变量理论为基础,研究自然现象的空间变异和空间结构)的发展,其理论和方法被应用于遥感影像的尺度效应问题研究。变异函数可衡量遥感影像的空间相关性(空间变异或空间依赖性),通过研究分析变异函数的结构可确定变量因子空间变异的尺度和程度,从而确定遥感影像的最佳空间分辨率[14]。实验变异函数表示如下[3]:
$$ \hat \gamma \left( h \right) = \frac{1}{{2N\left( h \right)}}\sum\limits_{i = 1}^{N\left( h \right)} {{{[Z({x_i}) - Z({x_i} + h)]}^2}} 。 $$ (1) 式(1)中: $\hat \gamma \left(h \right)$ 为空间相距h的全部点对的平均半方差,N(h)是间隔距离为h的点对的数目,Z(xi)和Z(xi+h)分别是间隔为h的样本点在各自位置上的像素。在变异程度内,变量具有空间自相关的特性,变异函数值随着h值增大逐渐增大;超出变异程度后,变量空间自相关性消失,变异函数值保持稳定不再变化[18]。此稳定值叫做基台值(c),表示该区域化变量的变异大小或程度;第1次达到基台值时的间隔距离叫做变程(a),表示区域化变量空间自相关范围或变异尺度[19-20]。h为0时的半方差叫做块金值(c0),主要由测量误差引起[21]。为了对实验变异函数进行数学描述,通常选用数学模型进行拟合表示,经典拟合模型有以下几种:指数模型、高斯模型、球状模型、线性模型等[22]。对遥感影像(地物类型主要或全部为森林植被)进行变异函数分析时,间隔距离h指的是像元空间分辨率大小与间隔像素数的乘积;由于本研究采用的是基于1个步长的变异函数,因此间隔像素数为1,间隔距离对应的就是影像空间分辨率的大小。本研究空间变异分析的对象为森林植被,故变程大小实际指的就是森林植被空间自相关的大小,变程大小与步长(影像空间分辨率)的乘积便是森林植被具有空间自相关的范围。对森林植被进行遥感提取时,影像单个像素内必须保证森林植被具有空间自相关性,因此,通过基于1个步长的变异函数对影像进行分析,可初步确定适合森林植被提取的影像最低空间分辨率。本研究利用经典的地统计学软件GS+,以1个像元(0.8 m)为步长,对研究区3种典型森林植被的0.8 m全色GF-2影像进行变异函数实验,采用拟合效果较好的指数模型和高斯模型,分析和确定研究区森林植被的空间变异尺度。
-
常用的分类算法有神经网络、随机森林、决策树等[23],基于计算量、人工参与度、分类效果等因素,本研究选择支持向量机监督分类方法,对多尺度的多光谱遥感影像集进行森林植被信息提取。森林植被提取精度包含两部分内容:面积精度和位置精度。为了对森林植被提取面积进行评估和评价,采用面积相差度进行衡量。
$$ 面积相差度 = \frac{{森林植被提取面积-森林植被实际面积}}{{森林植被实际面积}} \times 100\% 。 $$ 为了对森林植被提取位置的准确与否进行评估和评价,本研究将森林植被样点与多尺度影像森林植被提取结果进行对比分析,采用位置精度进行衡量。
$$ 位置精度 = \frac{{森林植被准确提取点数}}{{森林植被样点总数}} \times 100\% 。 $$ -
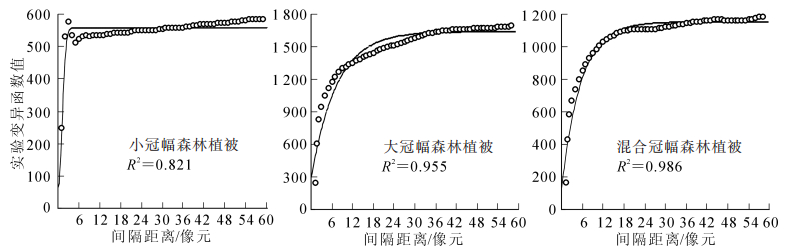
从图 2可看出:3种典型森林植被的空间变异情况不一致。这说明森林植被的空间分布情况不同,空间变异也不同,为探究适合森林植被提取的遥感影像最佳分辨率,必须对研究区内不同分布形态的森林植被进行单独的空间变异分析,然后才能综合判定最佳尺度。通过进行变异函数计算拟合,发现不同森林植被适合的变异函数模型也不一样,小冠幅森林植被最佳拟合模型为高斯模型,大冠幅森林植被和混合冠幅森林植被最佳拟合模型为指数模型,且3个模型的R2均大于0.8,说明模型的拟合效果都较好。表 1对应的分辨率是通过变程(a)乘以步长(0.8 m)而得。

图 2 3种典型森林植被以1个像元为步长的变异函数
Figure 2. Variation functions based on a step length of three typical forest vegetation
表 1 3种典型森林植被的变异函数模型拟合参数
Table 1. Variation function model fitting parameters of three typical forest vegetation
森林植被类型 模型类型 R2 基台值(c) 块金值(c0) 变程(a) 对应分辨率/m 小冠幅森林植被 高斯模型 0.821 557.3 60 2.77 2.2 大冠幅森林植被 指数模型 0.955 1 636.0 257 23.40 18.7 混合冠幅森林植被 指数模型 0.986 1 148.0 77 17.70 14.2 GF-2影像分辨率为0.8 m,为高空间分辨率遥感影像,可对各类林木进行精细刻画。从表 1可看出:对于小冠幅森林植被,基台较小为557.3,说明影像内地物均一,细节信息丰富,空间变异程度较低。变程较小为2.77,说明影像内小冠幅森林植被空间变异范围较小,空间分辨率≥2.2 m的影像可用于该区小冠幅森林植被信息提取。
对于大冠幅森林植被样地影像,块金值为257,基台值为1 636.0,都最大。影像内大部分是大冠幅林木,树冠大多相连,故空间变异反映的是林木的空间相关性。变程为23.40,是3种类型中变程最大的,说明影像内森林植被空间变异范围较大,空间分辨率≥18.7 m的影像可用于该区大冠幅森林植被信息提取。
对于混合冠幅森林植被,区域内均为森林植被且树冠基本相连,故空间变异反映的是林木的空间相关性。基台值较大为1 148.0,说明森林植被空间变异程度较高。变程为17.70,说明森林植被空间变异范围较大,空间分辨率≥14.2 m的影像均可用于该区混合冠幅森林植被信息提取。
通过分析3种典型森林植被样地影像的空间变异,可初步确定该地区森林植被提取的影像最低分辨率大致为16.0 m。但是要确定森林植被提取时所需影像的最佳分辨率,还需进行实际的分类效果比对和精度验证。
-
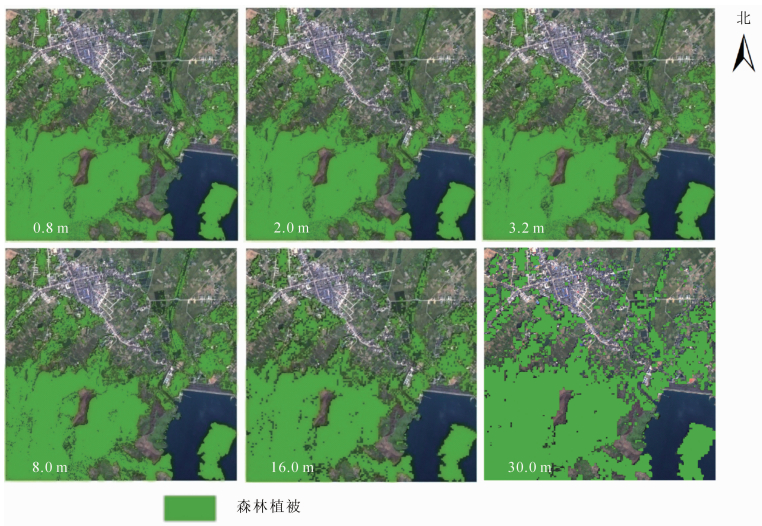
研究区不同尺度多光谱遥感影像通过支持向量机提取森林植被,结果与原始影像叠加显示如图 3。将图 3与森林植被调研结果进行对比可以发现,对于3种常见森林植被类型:①大冠幅森林植被(如大面积山体林区)。不同分辨率影像,提取效果相差不大,均较好。②混合冠幅森林植被(如城区散落分布林区)。分辨率高于8.0 m的影像提取效果均较好。③小冠幅森林植被(如行道树)。分辨率高于3.2 m的影像提取效果较好。

图 3 多尺度影像森林植被提取结果及GF-2影像叠加图
Figure 3. Overlay chart of forest vegetation extraction result by multi-scale images and GF-2 images
-
按照随机抽样、均匀分布的原则,根据野外实际调研的森林植被分布图,并结合Googel Earth高清遥感影像,选取360个森林植被样点,用于对多尺度影像森林植被提取结果进行位置精度评价。通过森林植被提取面积与实际调研面积的对比计算,以及森林植被提取结果与森林植被样点进行对比计算,具体精度评价情况见表 2。从表 2可看出:空间分辨率最高的影像(0.8 m),森林植被提取精度并非最高,面积相差度为0.92%,较好,位置精度却仅有82.2%,较差。这是由于空间分辨率较高时,影像对森林植被内部信息刻画很细致,不同树种、不同树龄的树冠均有差异,同物异谱情况较为严重,出现了“只见树木,不见森林”的现象,因此,用该影像进行森林植被提取时精度反而降低了。空间分辨率最低的影像(30.0 m),森林植被提取精度最低,面积相差度为5.43%,位置精度却仅有73.6%,均最差。这是因为研究区存在小冠幅林木,此类林地面积较小,30.0 m影像太粗略,不能对森林植被信息进行良好表达,因此,提取效果不好。空间分辨率为8.0 m的影像,森林植被提取精度最高,面积相差度为0.32%,位置精度有91.1%,均最好。空间分辨率为3.2和16.0 m的影像,森林植被提取效果较好,面积相差度均在3.00 %以下,位置精度均在85.0%以上。也就是说,基于遥感影像进行研究区森林植被提取时,影像空间分辨率最好在16.0 m以上。该判断与基于变异函数分析得到的该地区森林植被提取影像最低分辨率大致为16.0 m相一致,验证了基于1个步长的变异函数进行森林植被提取遥感影像分辨率选择的可靠性。
表 2 森林植被提取结果精度评价
Table 2. Precision evaluation of forest vegetation extraction result
影像分辨率/m 提取面积/hm2 面积相差度/% 准确提取样点数/个 位置精度/% 0.8 212.193 7 0.92 296 82.2 2.0 212.294 0 0.97 301 83.6 3.2 212.772 6 1.20 308 85.6 8.0 210.918 6 0.32 328 91.1 16.0 215.705 2 2.60 306 85.0 30.0 221.664 0 5.43 265 73.6 -
根据研究区多尺度影像森林植被提取精度评定的结果,0.8,2.0和30.0 m影像的森林植被提取结果均较差,位置精度均在85.0%以下,难以满足森林植被遥感高精度提取的要求,因此,本研究选取提取精度高于85.0%的适合森林植被提取的3.2,8.0和16.0 m影像进行林地提取效果综合评价(表 3)。表 3数据成本来源于中科遥感旗下遥感云服务平台——遥感集市。
表 3 森林植被提取结果综合评价
Table 3. Comprehensive evaluation of forest vegetation extraction result
影像分辨率/m 提取精度 数据成本/(元·km2) 处理时间 3.2 较好 30.0 较长 8.0 最好 1.5 较短 16.0 -般 免费 最短 针对3种常见的森林植被分布类型,综合考虑研究区森林植被提取精度、成本和效率,可以发现:①小冠幅森林植被适合提取最佳影像分辨率为3.2 m。②大冠幅森林植被适合提取最佳影像分辨率为16.0 m。③混合冠幅森林植被适合提取最佳影像分辨率为8.0 m。
本研究仅基于GF-2全色影像进行空间变异分析,未考虑多光谱影像的空间变异情况,未来将通过对GF-2多光谱影像或者WorldView,QuickBird等多种影像进行变异分析,以期进一步挖掘和完善森林植被提取最佳分辨率确定方法和结果。另外,为保证高中低分辨率影像分类方法的一致性,以便于统一对比分析,分类方法统一选择的是支持向量机监督分类法,未考虑面向对象的分类方法,未来将考虑用面向对象方法进一步开展深入研究。
-
基于1个步长的变异函数方法确定森林植被提取影像最低分辨率的方法比较可靠。通过多尺度影像森林植被提取结果得到森林植被提取影像最低分辨率为16.0 m,与基于1个步长的变异函数分析结果相一致,验证了该方法确定森林植被提取影像最低分辨率的可靠性,也可为其他地区、其他地物的遥感监测影像最佳空间分辨率选择提供参考。
确定三大典型空间分布的森林植被遥感监测影像的最佳空间分辨率为:小冠幅森林植被3.2 m;大冠幅森林植被16.0 m;混合冠幅森林植被8.0 m。该研究成果可为森林植被高精度、低成本、高效率遥感监测提供科学依据,丰富了森林植被遥感监测尺度研究,尤其为国产高分辨率影像数据(GF-1,GF-2)的行业应用提供了科学支撑。
Optimal resolution selection for monitoring forest vegetation using remote sensing images
-
摘要: 遥感影像分辨率的高低直接影响着森林植被监测的精度、成本和效率,故选择适合森林植被监测的影像最佳分辨率具有重要的应用价值。针对森林植被监测影像最佳分辨率选择方法及结果缺乏的问题,从林业实际应用出发,提出了基于1个步长的变异函数分析空间变异并综合考虑监测精度、成本和效率来确定森林植被监测影像最佳分辨率方法。基于最新的国产高分二号(GF-2)全色影像,利用1个步长的变异函数对湖南常宁洋泉镇林区3种典型分布类型森林植被进行拟合分析,初步确定适合森林植被监测的影像最低分辨率。然后对重采样形成的不同尺度多光谱影像分别进行监督分类,并对结果进行定量定性分析,结合影像成本和数据处理时间,找到适合不同类型森林植被监测的影像最佳分辨率。研究表明:不同分布类型的森林植被,适合遥感监测的影像最佳分辨率不同:①小冠幅森林植被3.2 m;②大冠幅森林植被16.0 m;③混合冠幅森林植被8.0 m。该森林植被遥感监测影像最佳分辨率确定方法和结果可为其他区域森林植被遥感监测影像最佳分辨率确定提供借鉴。Abstract: Image resolution directly affects precision, cost, and efficiency of forest vegetation extraction. To determine the best spatial resolution of remote sensing images for forest vegetation extraction, a method was proposed for optimal spatial resolution of remote sensing images when monitoring forest vegetation. Based on a panchromatic image of GF-2, a step length variation function was used to analyze forest vegetation of three distribution types considering monitoring precision, cost, and efficiency in Yangquan Town, Changning City, Hunan Province. A preliminary minimum resolution of the image suitable for forest vegetation extraction was determined. Then, through resampling of GF-2 multi-spectral images after fusion, middle and low resolution images of different scales were made to extract forest vegetation using the supervised classification method. Quantitative and qualitative analyses of results were carried out for accuracy of forest vegetation extraction, image cost, and data processing time. Results showed that the optimal spatial resolution for remote sensing monitoring images was different for different types of forest vegetation, with forest vegetation of small canopies being 3.2 m, for big canopies being 16.0 m, and for mixed canopies being 8.0 m. The optimal spatial resolution of remote sensing images to monitor forest vegetation could be used as a reference in other areas.
-
图 1 研究区位置及3种典型森林植被分布图
Figure 1 Location of the study area and distribution map of three typical forest vegetation
图 2 3种典型森林植被以1个像元为步长的变异函数
Figure 2 Variation functions based on a step length of three typical forest vegetation
图 3 多尺度影像森林植被提取结果及GF-2影像叠加图
Figure 3 Overlay chart of forest vegetation extraction result by multi-scale images and GF-2 images
表 1 3种典型森林植被的变异函数模型拟合参数
Table 1. Variation function model fitting parameters of three typical forest vegetation
森林植被类型 模型类型 R2 基台值(c) 块金值(c0) 变程(a) 对应分辨率/m 小冠幅森林植被 高斯模型 0.821 557.3 60 2.77 2.2 大冠幅森林植被 指数模型 0.955 1 636.0 257 23.40 18.7 混合冠幅森林植被 指数模型 0.986 1 148.0 77 17.70 14.2  下载: 导出CSV
下载: 导出CSV
表 2 森林植被提取结果精度评价
Table 2. Precision evaluation of forest vegetation extraction result
影像分辨率/m 提取面积/hm2 面积相差度/% 准确提取样点数/个 位置精度/% 0.8 212.193 7 0.92 296 82.2 2.0 212.294 0 0.97 301 83.6 3.2 212.772 6 1.20 308 85.6 8.0 210.918 6 0.32 328 91.1 16.0 215.705 2 2.60 306 85.0 30.0 221.664 0 5.43 265 73.6
下载: 导出CSV
表 3 森林植被提取结果综合评价
Table 3. Comprehensive evaluation of forest vegetation extraction result
影像分辨率/m 提取精度 数据成本/(元·km2) 处理时间 3.2 较好 30.0 较长 8.0 最好 1.5 较短 16.0 -般 免费 最短
下载: 导出CSV
-
[1] TORRESAN C, BERTON A, CAROTENUTO F, et al. Forestry applications of UAVs in Europe:a review[J]. Int J Remote Sens, 2017, 38(8/10):2427-2447. [2] 张东彦, 兰玉彬, 陈立平, 等.中国农业航空施药技术研究进展与展望[J].农业机械学报, 2014, 45(10):53-59. ZHANG Dongyan, LAN Yubin, CHEN Liping, et al. Current status and future trends of agricultural aerial spraying technology in China[J]. Trans Chin Soc Agric Mach, 2014, 45(10):53-59. [3] 柏延臣, 王劲峰.遥感信息的不确定性研究:分类与尺度效应模型[M].北京:地质出版社, 2003. [4] 梁文海, 刘吉凯, 张伟, 等.基于面向对象方法的GF-2影像桉树林信息提取[J].浙江农林大学学报, 2017, 34(4):721-729. LIANG Wenhai, LIU Jikai, ZHANG Wei, et al. Extracting Eucalyptus information using GF-2 images based on an object-oriented method[J]. J Zhejiang A & F Univ, 2017, 34(4):721-729. [5] 明冬萍, 王群, 杨建宇.遥感影像空间尺度特性与最佳空间分辨率选择[J].遥感学报, 2008, 12(4):529-537. MING Dongping, WANG Qun, YANG Jianyu. Spatial scale of remote sensing image and selection of optimal spatial resolution[J]. J Remote Sens, 2008, 12(4):529-537. [6] BAO Nisha, WU Lixin, LIU Shanjun, et al. Scale parameter optimization through high-resolution imagery to support mine rehabilitated vegetation classification[J]. Ecol Eng, 2016, 97(2):130-137. [7] TAO Shengli, GUO Qinghua, WU Fangfang, et al. Spatial scale and pattern dependences of aboveground biomass estimation from satellite images:a case study of the Sierra National Forest, California[J]. Landscape Ecol, 2016, 31(8):1711-1723. [8] PALENICHKA R, DOYON F, LAKHSSASSI A, et al. Multi-scale segmentation of forest areas and tree detection in LiDAR Images by the attentive vision method[J]. IEEE J Sel Topics Appl Earth Observ Remote Sens, 2013, 6(3):1313-1323. [9] HUANG Chunbo, ZHOU Zhixiang, WANG Di, et al. Monitoring forest dynamics with multi-scale and time series imagery[J]. Environ Monit Assess, 2016, 188(5):273. doi:10.1007/s10661-016-5271-x. [10] 张忠华, 胡刚, 祝介东, 等.喀斯特常绿落叶阔叶混交林物种多度与丰富度空间分布的尺度效应[J].生态学报, 2012, 32(18):5663-5672. ZHANG Zhonghua, HU Gang, ZHU Jiedong, et al. Scale-dependent spatial variation of species abundance and richness in two mixed evergreen-deciduous broad-leaved karst forests Southwest China[J]. Acta Ecol Sin, 2012, 32(18):5663-5672. [11] 徐俊锋. IKONOS信息提取的尺度效应研究[D]. 杭州: 浙江大学, 2006. XU Junfeng. Scale Issues in Information Extraction from IKONOS Data[D]. Hangzhou: Zhejiang University, 2006. [12] ATKINSON P M, CURRAN P J. Choosing an appropriate spatial resolution for remote sensing investigations[J]. Photogr Eng Remote Sens, 1997, 63(12):1345-1351. [13] 陈文凯, 何少林, 裴惠娟, 等.地震前后遥感影像空间尺度特性与最优空间尺度选择[J].地震, 2013, 33(2):19-28. CHEN Wenkai, HE Shaolin, PEI Huijuan, et al. Analysis and optimal selection of spatial scale for remote sensing images before and after earthquakes[J]. Earthquakes, 2013, 33(2):19-28. [14] 柏延臣, 王劲峰.基于特征统计可分性的遥感数据专题分类尺度效应分析[J].遥感技术与应用, 2004, 19(6):443-449. BAI Yanchen, WANG Jinfeng. Exploring the scale effect in thematic classification of remotely sensed data:the statistical separability-based method[J]. Remote Sens Technol Appl, 2004, 19(6):443-449. [15] 温兆飞, 张树清, 白静, 等.农田景观空间异质性分析及遥感监测最优尺度选择:以三江平原为例[J].地理学报, 2012, 67(3):346-356. WEN Zhaofei, ZHANG Shuqing, BAI Jing, et al. Agricultural landscape spatial heterogeneity analysis and optimal scale selection:an example applied to Sanjiang plain[J]. Acta Geogr Sin, 2012, 67(3):346-356. [16] 卫春阳, 徐丹丹, 董凯凯, 等.遥感影像空间格局变异函数分析研究进展[J].地球信息科学学报, 2017, 19(4):540-548. WEI Chunyang, XU Dandan, DONG Kaikai, et al. Advances in analysis of remote sensing image pattern based on semi-variogram[J]. J Geo-Inf Sci, 2017, 19(4):540-548. [17] 韩鹏, 龚健雅, 李志林.基于信息熵的遥感分类最优空间尺度选择方法[J].武汉大学学报(信息科学版), 2008, 33(7):676-679. HAN Peng, GONG Jianya, LI Zhilin. A new approach for choice of optimal spatial scale in image classification based on entropy[J]. Geom Inf Sci Wuhan Univ, 2008, 33(7):676-679. [18] CHILÈS J P, DELFINER P. Geostatistics: Modeling Spatial Uncertainty[M]. New York: John Wiley and Sons, 1999. [19] 王政权.地统计学及其在生态学中的应用[M].北京:科学出版社, 1999. [20] PANNATIER Y. Variowin: Software for Spatial Data Analysis in 2D[M]. New York: Springer Verlag, 1996. [21] ATKINSON P M, CURRAN P J. Defining an optimal size of support for remote sensing investigations[J]. IEEE Trans Geosc Remote Sens, 1995, 33(3):768-776. [22] DEUTSCH C V, JOURNEL A G. GSLIB:Geostatistical Software Library and User's Guide[M]. 2nd. New York:Oxford University Press, 1998. [23] FU Bolin, WANG Yeqiao, CAMPBELL A, et al. Comparison of object-based and pixel-based Random Forest algorithm for wetland vegetation mapping using high spatial resolution GF-1 and SAR data[J]. Ecoll Indic, 2017, 73:105-117. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2018.04.017
 点击查看大图
点击查看大图

计量
- 文章访问数: 4012
- HTML全文浏览量: 801
- PDF下载量: 557
- 被引次数: 0
