







-
发声现象在昆虫纲中普遍存在,其中有16个目的昆虫能发声,有11个目能以摩擦的方式发声[1-2]。鞘翅目Coleoptera中有30个科包含14种类型的摩擦器官,分别位于头盖部、胸部、腹部、足和鞘翅等部位[3-9]。AIKEN[6]总结了水龟虫科Hydrophilidae 16个属的昆虫具有摩擦发声器,这16个属分别为Tropisternus,毛腿牙甲属Anacaena,Berosus,Cymbiodyta,Derallus,Enochrus,Helophorus,Hemiosus,毛跗牙甲属Hydrobius,刺腹牙甲属Hydrochara,条脊牙甲属Hydrochus,牙甲属Hydrophilus,Laccobius,Limnoxenus,隆胸牙甲属Paracymus和Spercheus,且多数雌雄虫都有摩擦发声器。WESSEL[9]也认为绝大多数属的水龟虫两性个体都有摩擦发音器。昆虫摩擦发音的机制不尽相同,AIKEN[6]认为水龟虫是通过鞘翅-腹部的机制进行摩擦发声,虽然条脊牙甲属水龟虫摩擦器的结构和位置与其他种类不同,但是也通过鞘翅-腹部的机制进行摩擦发声。鞘翅目昆虫后翅翅脉分为6种类型:长扁甲型、肉食甲型、萤甲型、隐翅甲型、藻食甲型和缨翅甲型[10-12]。通过对翅的脉序、折叠方式和翅基骨片形态变化和功能的研究,可以揭示具翅昆虫的起源、目级分化和系统发育关系[10]。部分鞘翅目和双翅目类群均有翅退化的现象,因此鉴别现存的翅脉通常成为一个难题[10]。BALFOUR-BROWNE[13]以水生甲虫为对象研究了鞘翅目肉食亚目Adephaga的翅脉及与多食亚目Polyphaga的进化关系,并以Hydrophilus piceus为例研究了水龟虫科昆虫的翅脉。国内关于水龟虫的后翅脉序、摩擦器结构、发声机制等研究尚无报道。钝突刺腹牙甲Hydrochara affinis体型为长卵形,成虫体长为12.5~18.0 mm,是一种广泛分布于古北区的擅长游泳的中型水龟甲。本研究利用扫描电镜观察了钝突刺腹牙甲雌、雄两性的摩擦发音器,研究其发音机制,以期为水龟虫科昆虫的发音器官和听觉通信的研究提供参考。
-
钝突刺腹牙甲成虫于2016年7月灯诱采集于东北林业大学帽儿山实验林场(45°23′N,127°32′E;海拔为320 m)。将采集的甲虫样本,放于8 cm×20 cm×30 cm的水盆中饲养。
-
分别成对提起钝突刺腹牙甲雌雄成虫的前、中和后足,用镊子触碰其身体;再按压其腹部和鞘翅,观察研究其发声行为。雌雄虫各用30只。将死亡钝突刺腹牙甲雌雄成虫的鞘翅和后翅取下,用手动模拟的方式摩擦2部分。雌雄虫各用30只。
-
首先用Motic SMZ 168解剖镜和OLYMPUS SZX 12解剖镜观察钝突刺腹牙甲摩擦器的位置和形态,再用JSM 7500F扫描电镜(SEM)进一步观察分析摩擦发音器的结构。用常规标准方法干燥标本、固定和喷金[14-15]。
-
采集设备用SONY PCM-D100线性录音棒。
采集方法和条件参考文献[16-18]。为减少环境噪音的影响,19:00-20:00在室内(有日光灯照明)录音,室温为23 ℃;录音棒距离发音虫体5 cm。
采集声音信号的类型为自然声音信号,是指实验虫体在自由游泳活动时发出的声音信号。
-
钝突刺腹牙甲摩擦发音器的测量数据是对标本的直接测量和依据扫描电镜照片上的标尺测得,然后计算5只的平均值,没有使用软件测量。用SPSS 17.0统计软件对测量数据进行方差分析和多重比较,并用方差分析检验显著性(P<0.05)。
声音信号的分析处理参见文献[18-19]。用音频编辑处理软件Adobe Audition CS6读取钝突刺腹牙甲成虫游泳时采集的5个声音文件,进行选择性片段分割与脉冲组的确定与截取,对提取的特征脉冲组进行取样去噪,对持续时间进行测量。
-
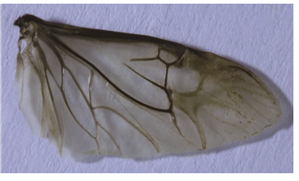
解剖镜下观察发现,钝突刺腹牙甲成虫的摩擦发音器均由发音锉和刮器2部分组成(图 1和图 2)。雌、雄成虫的发音锉位于后翅中部的径脉上(图 1),雌、雄成虫的刮器位于鞘翅外缘内侧的突起处(图 2),与发音锉的位置相对应。自然状态时发音锉完全被鞘翅遮盖,去掉鞘翅肉眼也可观察到。

图 1 钝突刺腹牙甲发音锉的位置
Figure 1. Position of the file of Hydrochara affinis

图 2 钝突刺腹牙甲刮器的位置
Figure 2. Position of the scrape of Hydrochara affinis
-
钝突刺腹牙甲后翅翅脉(图 3)属于萤甲型,这种脉序的主要特征是中脉(M)退化,但端部与肘脉(Cu)相连,臀脉(A)很发达,有径室和臀室(图 4)。钝突刺腹牙甲摩擦发音器所在的后翅中央位置正是径脉(R)位置,因钝突刺腹牙甲雌、雄成虫的发音锉均位于后翅的径脉(R),也就是说钝突刺腹牙甲雌、雄成虫的后翅径脉(R)上具有摩擦发音器。

图 3 钝突刺腹牙甲后翅图
Figure 3. Hindwing of Hydrochara affinis

图 4 萤甲型后翅模式图
Figure 4. Model hindwing of cantharid type
-
钝突刺腹牙甲后翅径脉中部的发音锉,由更微小的发音齿组成(图 5)。由表 1可见:雌虫的发音锉平均为(15.86±1.60)mm,比雄虫发音锉(14.28±1.20)mm略长。对发音齿的密度观察发现:发音锉的前、中、后部分布不同,中部的音齿比两侧的略显密集,但与两侧的密度差异不显著(P>0.05)。雌、雄两性间发音齿的密度差异也不显著(P>0.05)。据此推测:钝突刺腹牙甲成虫发出的声音不是专门用于两性间的沟通,而是个体之间的信息交流。

图 5 钝突刺腹牙甲发音锉的微细结构
Figure 5. Fine structure of file of Hydrochara affinis
表 1 钝突刺腹牙甲成虫发音锉的结构测量数据
Table 1. Fine data of file of Hydrochara affinis
性别 发音锉形状 发音锉长度/mm 发首锉宽度/μm 音齿平均密度/(个·mm-1) 前面 中间 后面 雌虫 弧形 15.86±7.60 162.10±81.70 15 18 16 雄虫 弧形 14.28±7.20 142.00±43.50 16 19 15 说明:所测数据为平均值±标准差 -
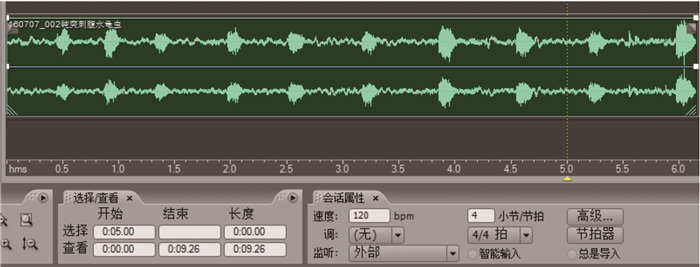
钝突刺腹牙甲成虫在自然状态和受刺激状态下均可发出响亮的摩擦声,声音在1 m以外可被清晰听见。在适当距离之内,可以听到钝突刺腹牙甲成虫游泳时发出的平和声音(图 6)。用镊子触碰试验昆虫的鞘翅,可以听到虫体发出的激烈摩擦声。如用手压鞘翅,同样有摩擦声发出。

图 6 钝突刺腹牙甲的声音波形
Figure 6. Wave of Hydrochara affinis
用镊子分别成对提起试验虫体的前、中、后足,使提起的每一对足不能有相对运动,钝突刺腹牙甲成虫均能够发出声音,这说明摩擦发声器官不在3对足上。按住试验虫体的头和胸部,使头胸部与腹部之间没有相对运动,钝突刺腹牙甲成虫也能发出声音,说明其发声器官也不在头胸部和腹部间。钝突刺腹牙甲成虫发声时,腹部鼓起,足也在运动,可见鞘翅也微微在动。多次实验和观察发现:只有腹部和鞘翅没有相对运动时,钝突刺腹牙甲成虫才不能发出声音,说明钝突刺腹牙甲成虫的发音器官在腹部和鞘翅间。
钝突刺腹牙甲成虫的刮器位于胸部第2节的鞘翅上,而发音锉位于胸部第3节。当它们运动而鼓起腹部时,导致身体的不同体节产生不一致的运动,使胸部第2节(着生刮器所在的鞘翅)和胸部第3节(着生发音锉所在的膜翅)产生相对运动,从而产生摩擦音。可见钝突刺腹牙甲成虫的发音机制是后翅径脉-鞘翅模式。
-
利用死亡的钝突刺腹牙甲雌、雄成虫的鞘翅和后翅径脉摩擦,模拟活体的发音。结果显示:雌雄两性用于模拟的这2部分都能够发出摩擦音。证明钝突刺腹牙甲的鞘翅摩擦后翅径脉能够发音,后翅径脉-鞘翅模式是鞘翅目昆虫摩擦发声的一种发音机制。
-
鞘翅目昆虫的摩擦发音器官可以出现在身体的任何部位,只要身体存在2个能够相互摩擦的坚硬表面,这也是此类昆虫特殊的进化适应能力。鞘翅目昆虫有高度几丁质化的外壳,它的表面分化出一系列纤细平行的脊,可以经受一个或者多个位于身体边缘的刮器摩擦[4]。甲虫发音器显示出一系列摩擦发声的独立性进化,虫体的任何连接处都可以因相对运动而发音,但人们对甲虫的声音通信和声音产生重要性的认知还非常有限[9]。
早期研究认为:水龟虫的摩擦发音器出现在后翅和鞘翅上,摩擦发音机制是前缘脉-鞘翅机制[20]。AIKEN[6]总结了水龟虫科16个属甲虫的发音,认为甲虫采用鞘翅-腹部机制声音,其中就有钝突刺腹牙甲所属的Hydrochara属。这与本研究结果不一致,说明AIKEN[6]的结论不能全面覆盖水龟虫科所有的摩擦发音机制,由于当时查阅文献数量少,可以说鞘翅-腹部机制产生声音是局限于当时的行为学观察而得出的结论。当然,Hydrochara属也可能还存在不同的摩擦发音机制。还有一种小概率的可能是鞘翅-腹部机制是错误的,因为AIKEN[6]的总结中,只有示意图而没有实物照片,而自然状态后翅的边缘紧邻腹侧片,在低倍镜下观察是容易混淆的。
本研究观察到钝突刺腹牙甲的发音锉在其后翅径脉上,与AIKEN[6]报道的龙虱科Dytiscidae 13个属的发音锉位置相似[6]。龙虱和水龟虫在形态上相似,它们的发音器位置也相似,这是生物学结构决定功能规律的一种体现。随着数字图像扫描电镜时代的到来,更多的昆虫发音机制的真相将被揭开和完善。鞘翅目金龟子科Scarabaeidae摩擦发音器也有在后翅相似位置的报道[21]。水龟虫和金龟子这两类甲虫,其发音器却出现在相似的位置,说明发音锉在鞘翅目内的起源不是孤立的,是有多个收敛的起源或者说有一个合适的主题方向[22]。甲虫发音器官的多样性比其他类群昆虫都高,只是对其研究的还不够深入,还不能总结其起源的共同规律。本研究表明:水龟虫科甲虫存在着后翅径脉-鞘翅模式的发音机制,但与前人研究存在差异,还需要进一步深入证实。
Fine structure and mechanisms of stridulatory organs in Hydrochara affinis
-
摘要: 为了解钝突刺腹牙甲Hydrochara affinis的摩擦发音器微细结构并探讨其发音机制,于东北林业大学帽儿山实验林场采集实验标本,利用扫描电镜(scanning electron microscope,SEM)观察钝突刺腹牙甲摩擦发音器微细结构,并采用实验促使发音方法探讨它的发音机理。结果显示:钝突刺腹牙甲的雌、雄成虫都具有摩擦发音器,雌虫的发音锉平均长为(15.86±7.60)mm,宽为(162.10±81.70)μm;雄虫发音锉平均长为(14.28±7.20)mm,宽为(142.00±43.50)μm。虫体通过摩擦后翅径脉-鞘翅产生声音。钝突刺腹牙甲雌、雄成虫发音主要用于个体间通信,发音机制属于后翅径脉-鞘翅模式。Abstract: Since no reports on the structure and sound formation of Hydrophilidae have been found in Chinese, Hydrochara affinis was used as an example to study the fine structure of Hydrophilidae stridulatory organs. Many specimens were collected from the state forest farm of Northeast Forestry University, and observations were made with a scanning electron microscope (SEM). Results showed that both males and females had stridulatory organs. The stridulatory organ size was (15.86±7.60) mm long and (162.10±81.70) μm wide for females but (14.28±7.20) mm long and (142.00±43.50) μm wide for males. The mechanisms of pronunciation for this kind of stridulatory organ were the same as in the model of the underwing radius vein-elytra. Thus, the pronunciation used may be for communication among different individuals of H. affinis.
-
Key words:
- entomology /
- file /
- fine structure /
- Hydrochara affinis
-
表 1 钝突刺腹牙甲成虫发音锉的结构测量数据
Table 1. Fine data of file of Hydrochara affinis
性别 发音锉形状 发音锉长度/mm 发首锉宽度/μm 音齿平均密度/(个·mm-1) 前面 中间 后面 雌虫 弧形 15.86±7.60 162.10±81.70 15 18 16 雄虫 弧形 14.28±7.20 142.00±43.50 16 19 15 说明:所测数据为平均值±标准差  下载: 导出CSV
下载: 导出CSV
-
[1] 彩万志.昆虫的发音及其在分类上的应用[J].昆虫知识, 1988(1):41-44. CAI Wanzhi. Insect making sound and its applicated in classification[J]. Entomol Knowl, 1988(1):41-44. [2] 曹凤勤, 程立生.昆虫鸣声的研究进展及其应用概述[J].华南热带农业大学学报, 2004, 10(1):29-32. CAO Fengqin, CHENG Lisheng. Progress in the research of insect acoustics and its application[J]. J South China Univ Trop Agric, 2004, 10(1):29-32. [3] ARROW G J. Sound-production in the lamellicorn beetles[J]. Trans Ent Soc Lond, 1904, 52(4):709-750. [4] ARROW G J. The origin of stridulation in beetles[J]. Proc R Ent Soc Lond, 1942, 17(7/9):83-86. [5] BARR B A. Sound production in Scolytidae (Coleoptera) with emphasis on the genus Ips[J]. Can Entomol, 1969, 101(6):636-672. [6] AIKEN R B. Sound production by aquatic insects[J]. Biol Rev, 1985, 60(2):163-211. [7] LYAL C H C, KING T. Elytro-tergal stridulation in weevils (Insecta:Coleoptera:Curculionoidea)[J]. J Nat Hist, 1996, 30(5):703-773. [8] HIRSCHBERGER P. Stridulation in Aphodius dung beetles:behavioral context and intraspecific variability of song patterns in Aphodius ater (Scarabaeidae)[J]. J Insect Behav, 2001, 14(1):69-88. [9] WESSEL A. Stridulation in the Coleoptera: an overview[C]//DROSOPOULOS S, CLARIDGE M F. Insect Sounds and Communication: Physiology, Behaviour, Ecology and Evolution. Boca Raton, FL: Taylor and Francis Group, 2006. [10] MAILLARD Y P. Premières observations comparatives sur I'appareil sonore élytro-abominal des Coléoptères Palpicornes[J]. Ann Stn Biol Besse-en-Chandesse, I969, 4:191-197. [11] 牛一平.基于后翅特征对全变态类昆虫系统发育关系的探讨[D].保定: 河北大学, 2011: 24. NIU Yiping. Phylogenetic Relationship Analysis of Holometabola Based on Hind Wing Character[D]. Baoding: Hebei University, 2011: 24. [12] 刘杉杉, 任国栋.中国齿甲属Uloma后翅的比较形态学研究(鞘翅目: 拟步甲科)[C]//任国栋, 张润志, 石福明.全国昆虫区系分类与多样性学术研讨会会议论文集.北京: 中国科学技术出版社, 2005: 215-218. [13] BALFOUR-BROWNE W A F. The wing venation of the Adephaga (Coleoptera) with special reference to the Hydradephaga and some homologies with the Polyphaga[J]. J Royal Microsc Soc, 1943, 63(3/4):55-64. [14] 彩万志, 庞雄飞, 花保祯, 等.普通昆虫学[M]. 2版.北京:中国农业大学出版社, 2011. [15] 李玲, 迟德富, 李燕, 等. 3种天牛科昆虫摩擦发音器的超微结构及摩擦发音机制分析[J].南京林业大学学报(自然科学版), 2013, 37(1):19-24. LI Ling, CHI Defu, LI Yan, et al. Stridulatory organs ulstrstructure of three longhorned beetles and their stridulation mechanism[J]. J Nanjing For Univ Nat Sci Ed, 2013, 37(1):19-24. [16] 王鸿斌, 赵丽稳, 罗茜, 等.红脂大小蠹声音信号记录、分析与发声器官电镜观察[J].林业科学研究, 2012, 25(1):17-22. WANG Hongbin, ZHAO Liwen, LUO Qian, et al. Acoustic signals of Dendroctonus valens and structure of its stridulatory apparatus[J]. For Res, 2012, 25(1):17-22. [17] LI Ling, SUN Fan, HU Jihua, et al. Ultrastructure of stridulating organ of Xylotrechus rusticus L. and behavioral responses to resisting sound(Coleoptera, Cerambycidae)[J]. J For Res, 2013, 24(2):547-552. [18] 娄定风, 许小芳, 李嘉, 等. 6种木材钻蛀性昆虫的声学特征与比较[J].植物检疫, 2013, 27(1):6-10. LOU Dingfeng, XU Xiaofang, LI Jia, et al. Acoustic characteristics and their comparison of six species of wood borers[J]. Plant Quar, 2013, 27(1):6-10. [19] 何忠, 陈念丽.北京地区5种常见鸣虫的鸣声结构[J].动物学报, 1985, 31(4):324-330. HE Zhong, CHEN Nianli. The sound structure of five species of common chirp insects in Beijing[J]. Acta Zool Sin, 1985, 31(4):324-330. [20] 马玉堃, 孙凡, 李玲. 2种墨天牛摩擦发音器超微结构的比较研究[J].安徽农业科学, 2014, 42(18):5895-5896, 5923. MA Yukun, SUN Fan, LI Ling. Comparative study on the ulstra-structure of stridulation apparatus in two species of Monochamus[J]. J Anhui Agric Sci, 2014, 42(18):5895-5896, 5923. [21] BROCHER F. L'appareil stridulatoire de I'Hydrophilus piceus et celui de Berosus aericeps[J]. Ann Biol Lacustre, 1912, 5:215-217. [22] BUHK F. Stridulationsapparat bei Spercheus emarginatus Schall[J]. Zeitschrift Für Witsenschaftliche Insektenbiologie, 1910, 6:342-346. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2019.01.018
 点击查看大图
点击查看大图

图(6) / 表(1)
计量
- 文章访问数: 3581
- HTML全文浏览量: 810
- PDF下载量: 138
- 被引次数: 0
