







-
灌木层是森林生态系统的重要组成部分,它参与森林生态系统物质循环和能量转化[1]。林下微环境是不断变化的,它对森林生态系统的稳定发展、演替进程及物种多样性起着重要作用[2]。中国灌木林面积约为2.97×107 hm2[3],与乔木层相比,林下植被生物量仅占总森林生物量的10%~30%,处于次要地位[4]。虽然灌木生物量占整个森林生物量比例较小,但由于灌木层在维持森林群落稳定性、物种多样性及水土保持效益上的重要作用,其生物量的估算与研究对森林生态群落物质的循环与固定、积聚与消耗以及能量转换具有相当重要的意义[5-9]。探讨森林群落生物量的方法主要有样地全收获、直接收获和拟合植物生物量与其形态参数的生物量模型来推算[10]。直接收获法浪费人力物力,且对林下微环境也存在很大的不确定性,样株数又很难符合统计学大样本的要求[11]。通过拟合生物量最优模型的方法来估算林下灌木的生物量不仅能够降低对林下植被的破坏,而且还可长期追踪样地林下灌木生物量的变化[12-13]。特别是大范围森林生物量调查,利用生物量模型能大大减少调查工作量[14-15]。且拟合适用于大范围的通用性立木生物量模型,是开展全国森林碳储量监测必不可少的基础工作[16]。目前,生物量模型主要分为线性、非线性以及多项式[17]。国内外学者对吉林省金沟岭林区林下植被生物量研究集中在地上地下生物量的研究中[18-19],很少考虑物种间和各物种不同器官生物量最优模型的差异及建立单一物种混合最优模型,样本量及物种个数范围较小[13, 20-21]。本研究利用野外调查的大量实测数据,涵盖了该区出现频率较高的所有灌木,且运用灵活性更高的R软件[22]拟合该区出现频率较高的13个物种生物量模型,并估算了不同密度冷杉Abies nephrolepis,鱼鳞云杉Picea jezoensis和红皮云杉Picea koraiensis林(以下称云冷杉林)灌木层的生物量,为研究金沟岭不同林型不同密度林分与灌木层生物量的关系提供基础依据。
HTML
-
研究区位于吉林省汪清县金沟岭林场(130°10′E,43°22′N),经营森林面积为16 286 hm2。该区属低山丘陵地貌,海拔为300~1 200 m,坡度5°~25°。全年平均气温约为3.9 ℃,7月气温最高,在22.0 ℃左右;年降水量为600~700 mm,多集中于7月。研究区主要的乔木有冷杉,鱼鳞云杉,红皮云杉,红松Pinus koraiensis,白桦Betula platyphylla,枫桦Betula costata等。主要林下灌木有毛榛Corylus mandshurica,暴马丁香Syringa amurensis,忍冬Lonicera japonica,溲疏Deutzia scabra等。主要地被植物为少数灌木和禾本科Poaceae草类,如珍珠梅Sorbaria sorblia,柳叶绣线菊Spiraea salicifolia等。
-
以云冷杉林为研究对象,设置3种郁闭度(0.6,0.8,1.0) 20 m × 20 m的样地,灌木样方5 m × 5 m设置在每个样地的中心及4个拐角,测量每株灌木的地径(D,mm)或胸径、冠幅(C,m )和株高(H,m)。根据灌木调查结果,在样地附近选择分布较广的13个物种,取15株·种-1左右。样株要能代表研究灌木的地径范围,取样之前测地径D,株高H,冠幅C1和C2,再将整株全挖出,地上分为树干、树枝、树叶分别称出鲜质量,将根系土全部清除后再称量,将称量的各器官装入信封带回实验室,在85 ℃烘箱内烘干得到各器官干质量和含水率。取样灌木的基本参数见表 1。
物种 样本数/个 地径/mm 株高/m 冠幅/m 灌木类型 平均值 标准差 平均值 标准差 平均值 标准差 土庄绣线菊 14 4.786 2.292 1.003 0.542 0.384 0.206 典型灌木 刺五力口 Acanthopanax senticosus 12 12.131 2.113 1.482 0.387 0.597 0.204 典型灌木 溲疏! 18 6.941 2.894 0.932 0.312 0.432 0.256 典型灌木 野蓄薇%osa :u;ti/Zora 8 5.601 1.557 0.721 0.231 0.379 0.157 典型灌木 山梅花 Philadelphus incanus 11 11.305 2.757 1.830 0.511 0.865 0.318 乔木型灌木 茶薦子 %i+es mandshuricum 10 5.518 1.612 1.154 0.518 0.551 0.333 典型灌木 珍珠梅 Sorbaria sorbifolia. 14 12.731 6.017 1.393 0.524 0.388 0.207 典型灌木 青措械—,er te>mentosum 17 10.473 7.372 1.398 1.234 0.555 0.486 乔木型灌木 花措械—,er u?urunduense 11 8.583 7.325 1.204 0.875 0.492 0.713 乔木型灌木 暴马了香 Syninga reticulata var. mandsshuca 14 11.456 6.478 1.387 0.753 0.909 0.851 乔木型灌木 忍冬 Lonicerajaponica 14 11.721 4.713 1.679 0.644 0.945 0.363 乔木型灌木 毛棒子 Corylus mandshurica 17 11.798 8.212 1.613 1.188 0.799 0.675 乔木型灌木 惚木 Aralia chinensis 14 15.454 6.763 1.259 0.533 0.943 0.349 乔木型灌木 Table 1. General parameters of sampling shrubs
-
本次生物量模型的建立,选用了单因子地径D和复合因子冠幅乘株高CH,植冠面积AC(AC=πC1C2/4)及D2H为自变量,通过阅读相关文献[13, 15, 24-28],建立灌木生物量模型。不同物种生长规律不同,植物形态也会有所不同。因灌木各测树因子间存在着很强的关联性,所以希望得到灌木的生长变化和某种函数趋势接近,且选取方程和自变量时要考虑植物的形态。所以选用了指数、幂函数、二元一次和一元二次函数。选用二元一次函数的生物学意义是影响灌木生长的因素可能不止一个。在R软件中采用非线性最小二乘法拟合了灌木不同器官(干、枝、叶、根)的生物量模型和单一物种生物量混合模型。模型均进行t检验(P<0.05),分别使用判定系数R2,剩余标准差ESEE,平均相对误差E以及F检验值作为评价模型优劣的指标。选出R2和F最大,ESEE和E值最小的方程作为最优方程。将选出的模型应用于不同密度的云冷杉林,计算出灌木层的生物量,而其他没有建立生物量模型的灌木采用何列艳等[16]建立的乔木型和典型灌木混合模型计算出生物量。4种函数形式如下[14-27]:W=axb,W=aebx,W=a+bx+cx2,W=a+bx1+cx2。a,b,c为各模型参数,x为建模因子。利用Excel 2013和R软件[22]对数据进行统计分析。
1.1. 研究区概况
1.2. 数据的收集
1.3. 数据分析
-
根据实测因子和生物量分布情况,利用R软件[23]的处理结果,得到13个物种的生物量模型。由表 2知:除溲疏根以外,其他物种不同器官模型和单一物种混合模型的回归关系显著(P<0.001 0),模型均具有较高的R2,F值和较小的ESEE,E值,R2为0.626 2(溲疏根)~0.997 9(花楷槭干),溲疏除干和叶的R2达0.900 0以上,其他器官模型R2相对较低。F值为20.650 0(溲疏根)~4 040.340 0(花楷槭干),F值越大,模型精度相对越高。ESEE值为0.000 7(绣线菊干)~0.027 5(楤木干),各物种不同器官模型的ESEE值除青楷槭根外,其他物种不同器官模型的ESEE值均小于各自单一物种混合模型的ESEE值。E值为-7.48×10-17(刺五加干)~0.000 3(茶藨子枝),E值也较小,说明预估精度相对越高。各物种不同器官的最优模型除青楷槭的叶和根,花楷槭的干为幂函数,其他物种不同器官最优模型多为一元二次或二元一次函数,说明青楷槭叶和根、花楷槭干的生长分布近似于幂函数的分布趋势;其他近似于一元二次函数分布,模型为二元一次函数的说明影响该物种生物量模型的因子至少有2个且生长趋势与其相近。另外,茶藨子的各个模型均为指数函数,但不适用于地径较大的植株。干、枝最优模型的自变量除绣线菊为AC,花楷槭为CH外,其他多为D2H和CH,即影响干枝生物量的因子多与灌木的地径和高有关;叶最优模型的自变量,绣线菊、刺五加和青楷槭采用因子AC和CH,其他物种多采用D和D2H,即不同植物叶片不同,影响叶生物量的因子也会有所不同;灌木根系多采用D2H,说明在此地区,影响植物根系生物量因子多为此复合因子。综合各个模型可知,影响灌木生物量模型的因子多为D和复合因子D2H,AC。
-
单一物种混合模型R2为0.837 1(溲疏)~0.997 0(花楷槭),且除溲疏和土庄绣线菊以外,其他混合模型R2都达到了0.920 0以上,F值的变化规律与R2相似,R2越大F值相对越大,单一物种生物量模型拟合越好。单一物种混合模型的ESEE为0.008 2(茶藨子)~0.056 9(楤木),除忍冬和楤木的ESEE值达到0.050 0以外,其他物种的ESEE值均较小且E值也较小。单一物种混合模型除土庄绣线菊和茶藨子为指数函数,自变量为AC和D外,其他多为一元二次或二元一次函数,说明土庄绣线菊和茶藨子在一定的时间范围内,生物量增加的方式类似指数分布。溲疏、青楷槭最优方程采用自变量D2H,花楷槭、暴马丁香、忍冬和毛榛采用自变量为D2H和CH,除忍冬、溲疏外,其他物种的植物形态较为相似,且大部分都采用了因子D2H,说明植株形态相似,影响其生物量的因子也较为相似,综合这13个物种的单一生物量最优模型知,影响灌木生物量的因子多为复合因子D2H。从表 2中还可以看出:青楷槭、毛榛、忍冬的单一物种生物量混合模型的判定系数高于它们不同器官生物量模型的判定系数。其他物种混合模型的判定系数介于各器官模型的判定系数之间。
物种及地径范围 最佳模型 参数 E ESEE R2 F值 显著性 a b c 土庄绣线菊0.21>0.98cm W干=aeb(AC) 0.018 1 2.031 2 1.8500E-05 0.003 7 0.8517 58.263 1 0.000 0 W枝=a+b(AC)+c(AC)2 0.018 4 0.000 2 0.116 1 -5.84E-10 0.002 0 0.9452 134.7740 0.000 0 W叶=a+b(AC)+c(CH) 0.018 1 0.024 2 -0.003 1 -9.66E-10 0.000 7 0.8989 87.819 1 0.000 0 W根=a+b(D)+c(AC) 0.015 3 0.000 9 0.026 9 -1.09E-10 0.003 6 0.7458 29.893 4 0.000 0 W总=aeb(Ac) 0.071 1 1.609 9 0.0000424 0.008 7 0.8882 80.560 4 0.000 0 刺五加0.28~1.58cm W干=a+b(AC)+c(D2H) 0.015 7 0.026 5 0.000 1 -7.4800E-17 0.004 6 0.9626 403.133 7 0.000 0 W枝=a+b(AC)+c(CH) 0.011 7 -0.063 1 0.045 9 6.9700E-10 0.005 8 0.8302 84.057 4 0.000 0 W叶=a+b(AC)+c(CH) 0.013 9 -0.068 5 0.046 5 7.9900E-10 0.006 5 0.7765 67.985 2 0.000 0 W根=a+b(cH)+c(CH)2 0.045 4 -0.036 6 0.036 3 2.8300E-09 0.007 5 0.8860 116.2644 0.000 0 W总=a+b(AC)+c(CH) 0.059 4 -0.135 8 0.173 0 2.2400E-09 0.018 8 0.9242 221.681 6 0.000 0 溲疏0.22~1.35cm W干=a+b(D2H)+c(D2H)2 -0.000 7 0.000 2 -1.760 0E-07 4.4100E-11 0.002 2 0.9727 482.007 7 0.000 0 W枝=a+b(D2H)+c(D2H)2 -0.0022 0.000 3 -4.600 0E-07 -7.4100E-11 0.007 0 0.7257 35.492 0 0.000 0 W叶=a+b(D)+c(D2H) 0.001 8 -0.0004 7.7300E-05 8.6000E-12 0.001 4 0.9025 109.219 0 0.000 0 W根=a+b(D2H)+c(D2H)2 -0.003 2 0.000 5 -1.260 0E-06 1.7900E-11 0.011 2 0.6262 20.650 5 0.001 0 W总=a+b(D2H)+c(D2H)2 -0.006 0.001 0 -1.8500E-06 9.7300E-12 0.018 6 0.8374 70.028 2 0.000 0 青楷槭2.98~23.31cm W干=a+b(Ac)+c(CH) -0.002 4 -0.051 9 0.0523 -5.6000E-09 0.008 2 0.9827 728.609 3 0.000 0 W枝=a+b(D)+c(D)2 0.005 4 -0.002 3 0.003 -2.56E-11 0.008 8 0.9452 223.781 7 0.000 0 W叶=aDb 2.740 0E-05 2.309 0 9.6600E-05 0.002 7 0.9796 707.487 3 0.000 0 W根=aDb 2.510 0E-14 9.110 5 0.0101 0.023 5 0.9527 154.291 8 0.000 0 W总=aD(D2H)+c(D2H)2 0.0021 0.003 -7.67E-08 -1.100 5E-09 0.0140 0.9892 1 242.204 7 0.000 0 花楷槭2.88~28.90 mm W干=a(CH)b 0.0250 1.1229 -0.0007 0.0034 0.9979 4050.3455 0.0000 W枝=a+b(Ac)+c(CH) -0.0010 0.0194 0.0046 -5.2000E-09 0.0032 0.9932 1070.311 0.0000 W叶=a+b(D2H)+c(D2H)2 0.0012 2.4600E-05 2.5900E-09 1.3800E-11 0.0032 0.9872 545.4946 0.0000 W根=a+b(Ac)+c(CH) -0.0044 -0.0378 0.0408 1.2000E-11 0.0052 20.9897 723.6879 0.0000 W总=a+b(D2H)+c(CH) 0.0060 4.2900E-05 0.0634 -6.5800E-11 0.0108 0.9970 2476.331 0.0000 暴马丁香4.20~23.13 mm W干=a+b(D2H)+c(CH) 0.0115 0.0001 0.0103 -1.8202E-12 0.0119 0.9772 282.1242 0.0000 W枝=a+b(CH)+c(D2H) 0.0106 0.0001 0.0103 -1.8200E-12 0.0119 0.9577 210.5066 0.0000 W叶=a+b(Ac)+c(CH) 0.0134 -0.0087 0.0197 -7.0101E-10 0.0038 0.9772 282.1242 0.0000 W根=a+b(D2H)+c(CH) 0.0265 0.0001 0.0087 2.3200E-11 0.0192 0.8654 63.0999 0.0000 W总=a+b(D2H)+c(CH) 0.0618 0.0003 0.0489 1.4000E-07 0.0397 0.9656 257.5155 0.0000 忍冬3.84~20.75 mm W干=a+b(D2H)+c(CH) -0.0219 -0.0001 0.0869 1.5400E-10 0.0271 0.8881 67.8130 0.0000 W枝=a+b(CH)+c(CH)2 -0.0131 0.0630 -0.0058 9.4200E-09 0.0283 0.7431 25.9984 0.0000 W叶=a+b(D)+c(Ac) 0.0152 0.0007 0.0125 7.3400E-10 0.0040 0.8830 65.2186 W根=a+b(Ac)+c(CH) -0.0023 -0.0416 0.0608 1.2700E-10 0.0220 0.8608 49.4119 0.0000 W总=a+b(D2H)+c(CH) 0.0002 -0.0001 0.1784 9.9900E-08 0.0549 0.9205 101.6929 0.0000 毛榛子2.19~26.58mm W干=a+b(Ac)+c(CH) -0.008 -0.0136 0.0439 4.1300E-09 0.0184 0.9690 285.0368 0.0000 W枝=a+b(CH)+c(CH)2 -0.0025 0.0157 -0.0007 2.3500E-10 0.0094 0.8983 97.4586 0.0000 W叶=a+b(AC)+c(D2H)2 -9.6400E-05 1.4600E-14 -6.8800E-08 -7.2600E-11 0.0096 0.9041 101.2987 0.0000 W根=a+b(D2H)+c(D2H)2 -9.6400E-05 1.460E-04 -6.8800E-08 -7.2600E-11 0.0096 0.9041 101.2987 0.0000 W总=a+b(D2H)+c(CH) 0.0035 6.7500E-05 0.0470 3.0000E-09 0.0235 0.9798 540.8142 0.0000 茶藨子2.82~7.60mm W干=aeb(D) 0.00003 1.0541 -0.0002 0.0088 0.8996 48.3694 0.0000 W枝=aeb(D) 3.000E-6 1.3083 0.00036 0.9645 124.6582 0.0000 W叶=aeb(D) 0.0002 0.4997 -0.0001 0.0008 0.9145 81.3773 0.0000 W根=aeb(D) 0.0007 0.5182 -0.0005 0.0036 0.9206 93.0817 0.0000 W总=aeb(D) 0.0002 0.8198 0.0002 0.0082 0.9812 309.8734 0.0000 楤木6.10~27.33 W干=a(D2H)b 0.0002 1.0251 -1.3100E-03 0.0275 0.9506 202.9919 0.0000 W枝=aeb(D) 0.0014 0.1567 -1.7500E-05 0.0123 0.8760 70.6008 0.0000 W叶=a+b(D)+c(Ac) -0.0037 0.0008 0.0205 -1.000E-7 -1.1600E-10 0.0181 0.8430 57.4457 0.0000 W根=a+b(D2H)+c(D2H)2 0.0013 0.0002 -1.0000E-7 -1.1600E-10 0.0181 0.8430 57.4457 0.0000 W总=a(D2H)b 0.0002 1.1742 6.4400E-04 0.0280 0.9419 128.7706 0.0000 山梅花5.72~15.70mm W干=a+b(Ac)+c(D2H) -0.0111 0.0102 0.0002 -3.0100E-11 0.0070 0.9771 302.6944 0.0000 W枝=a+b(Ac)+c(D2H) -0.0131 0.0092 0.0002 3.1900E-11 0.0186 0.8169 31.6464 0.0000 W叶=a+b(CH)+c(CH)2 0.0082 -0.0100 0.0044 6.2400E-13 0.0023 0.8112 3.8272 0.0000 W根=a+b(D2H)+c(CH) 0.0080 0.0002 -0.0125 2.8100E-12 0.0066 0.9553 135.1367 W总=a(D2H)b 0.0002 1.1742 6.4400E-04 0.0280 0.9419 128.7706 0.0000 野蔷薇3.66~8.33mm W干=a+b(D2H)+c(D2H)2 -0.0011 0.0003 -6.0000E-7 3.9800E-12 0.0007 0.9925 611.4494 0.0000 W枝=a+b(D)+c(D)2 -0.0168 0.0059 -0.0004 -4.7000E-14 60.7921 17.2780 0.0000 W叶=a+b(AC)+c(CH) -0.0023 0.0078 0.0252 -2.6900E-10 0.0013 0.9755 177.1763 0.0000 W根=a+b(D)+c(Ac) -0.0128 0.0041 0.1099 2.6100E-09 0.0058 0.9349 67.8752 0.0000 W总=a+b(D)+c(Ac) -0.0386 0.0108 0.1533 2.3000E-09 0.0054 0.9823 2.688038 0.0000 珍珠梅4.21~24.53mm W干=a+b(D2H)+c(D2H)2 -0.0109 0.0004 0.0000 1.0700E-09 0.0199 0.9430 164.9001 0.0000 W枝=a+b(D)+c(Ac) -0.0193 0.0022 0.0720 8.5500E-09 0.0093 0.8962 88.9694 0.0000 W叶=a+b(Ac)+c(D2H) -0.0008 0.0442 0.00003 2.4400E-10 0.0059 0.9466 172.1479 0.0000 W根=a+b(D)+c(CH) -0.0288 0.0057 -0.0168 -1.0600E-09 0.0164 0.7612 34.1580 0.0000 W总=a+b(D2H)+c(D2H)2 -0.0055 0.0006 -2.0000E-7 4.6500E-11 0.0262 0.9735 389.0325 0.0000 Table 2. Best-fit component biomass models for specific species and mixed models for single species
-
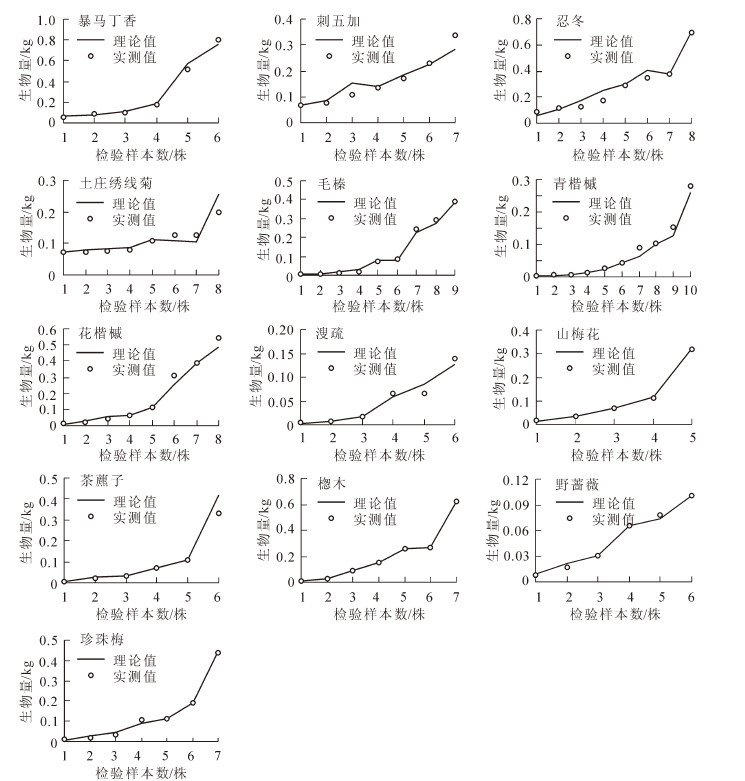
用2015年7-8月的实测数据作为检验样本(5~10株)对各模型进行检验,检验结果如图 1和表 3,各物种生物量实测值与理论值除溲疏和毛榛中的一个拟合率为60.00%,其余均达到70.00%以上,且有7个物种的拟合率为90.00%以上。13个物种的单一物种混合模型平均拟合率均达到82.00%以上,范围为82.77%(溲疏)~98.19%(楤木),溲疏和忍冬的平均拟合率比其他物种要低,因为溲疏生物量模型的判定系数比其他物种也相对较低。楤木的平均相对误差只有1.82%,且平均拟合率达到98.19%,模型的拟合度很高;茶藨子、野蔷薇和山梅花的平均相对误差也只约5.00%,模型拟合度也较高;除忍冬和溲疏约18.00%,这是由于样本数量较小导致精度并不理想,其他均为11.00%左右,模型拟合精度满足估算要求,模型可用于本地区生物量估算。
灌木名称 平均相对误差/% 平均拟合率/4 暴马丁香(6) 10.29 90.56 刺五加(7) 11.61 90.43 忍冬(8) 18.28 84.97 绣线菊(>) 11.85 89.32 茶薦子(5) 5.32 95.66 山梅花(6) 4.37 95.64 珍珠梅(7) 14.43 87.53 毛榛(9) 11.43 90.75 青揩槭(10) 14.74 86.51 花楷槭(8) 12.22 89.56 溲疏(6) 18.29 82.77 惚木(7) 1.82 98.19 野蔷薇(6) 6.38 94.47 说明:括号里数据为检验样本个数。 Table 3. Verification of different species biomass models

Figure 1. Verification of different species biomass models
-
将单一物种混合模型应用于金沟岭林场不同密度的云冷杉林林下灌木层生物量的估算,结果见表 4。冷杉林下灌木生物量随着密度的不同差异显著,随着林分密度减小,云冷杉林下灌木生物量出现先减小后增大的趋势,即W(0.6)>W(1.0)>W(0.8)。当郁闭度为0.8时,灌木生物量最小。调查可知,这是因为此密度下有较多的冷杉更新苗。灌木株数相对较小,导致此时灌木的生物量也较小。
灌木名称 生物量/kg 暴马丁香 16.036 6 4.412 9 0.485 3 刺五加 2.4149 2.679 3 3.697 7 花楷槭 3.322 3 0.186 2 0.000 0 青楷槭 5.741 6 0.577 5 2.955 1 忍冬 0.172 4 11.227 2 18.153 7 毛榛子 0.969 4 0.809 1 0.256 1 溲疏 0.000 0 0.152 1 1.852 3 绣线菊 0.000 0 5.308 8 5.265 6 其他 4.792 1 2.348 1 0.475 5 合计 33.449 2 27.701 1 33.141 3 说明:郁闭度 0.6 生物量为 836.230 0 kg.hm -2,郁闭度 0.8 为 692.527 5 kg.hm-2,郁闭度 1.0 为 828.535 2 kg.hm-2。 郁闭度0.6 郁闭度0.8 郁闭度1.0 Table 4. Biomass results of shrubs under three density spruce-fir forest
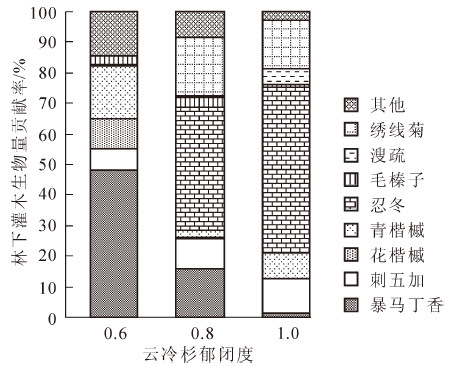
不同密度云冷杉林下灌木生物量贡献率用Origin 85制图(图 2)。对郁闭度为0.6的林分林下灌木生物量而言,暴马丁香的贡献率最大,达47.94%,其次是青楷槭,达17.16%,且其他灌木的贡献率达到14.32%;郁闭度为0.8时,灌木生物量贡献率最大的为忍冬,达40.53%,其次为绣线菊、暴马丁香;郁闭度为1.0时,灌木生物量贡献率最大的也为忍冬,达53.96%,其次为绣线菊、刺五加,分别达到了15.65%和10.99%。

Figure 2. Ratio of shrub biomass under different density spruce-fir forest
2.1. 灌木生物量最优模型
2.1.1. 各物种不同器官生物量模型
2.1.2. 单一物种生物量混合模型
2.1.3. 单一物种生物量混合模型的检验
2.2. 各物种生物量模型的应用
-
本研究拟合了金沟岭林场灌木层出现频率较高的13个植物的单一物种生物量混合模型和各物种不同器官生物量最优模型,并估算了3种不同密度云冷杉林下灌木层的总生物量。结果表明:每个模型均具有较高的判定系数R2值、F值和比较小的剩余标准差(ESEE)值、平均相对误差E值,R2为0.626 2(溲疏根)~0.997 9(花楷槭干),除了青楷槭的根外,其他物种不同器官模型的ESEE值均小于各自单一物种混合模型的ESEE值;各物种混合模型除了溲疏和土庄绣线菊以外,其他物种的生物量混合模型R2都达到了0.920 0以上;单一物种混合模型的 ESEE和平均相对误差E值均较小。通过对模型的检验,模型的平均相对误差和平均拟合率也较好,综合各项指标,本次拟合的65个模型精度均较高,满足估算要求,为今后测量森林生物量提供了可靠的方法依据。不同器官生物量最优模型除青楷槭的叶根,花楷槭干为幂函数,其他不同器官最优模型多为一元二次或二元一次函数。干、枝最优模型的自变量多为D2H和CH;叶最优模型的自变量,绣线菊、刺五加和青楷槭多采用AC和CH,其他物种多采用D和D2H;根系多采用的因子为D2H。单一物种混合模型除了土庄绣线菊为指数函数,其他的多为一元二次或者二元一次函数。将拟合的模型应用于3种密度的云冷杉林,分别得到郁闭度为0.6,0.8,1.0的林分林下灌木生物量为836.230 0,692.527 5,823.535 2 kg·hm-2。天然云冷杉林林下灌木生物量(W)随着林分郁闭度减小出现先减小后增大的趋势,即W(0.6)>W(1.0)>W(0.8)。郁闭度为0.8时,灌木生物量最小,是因为此密度下有较多的冷杉更新苗,导致此时灌木的生物量较小。郁闭度为0.6的林分,暴马丁香的贡献率最大,达47.94%;郁闭度为0.8和1.0时,贡献率最大的为忍冬,分别达40.53%和53.96%。在主林层密度较大时,忍冬对灌木层生物量的贡献要大于其他物种。
金沟岭林场林下灌木种类丰富,很多物种属于大灌木或者小型乔木,若采用直接收获的方法,很难估算出像青楷槭等乔木型灌木的生物量,构建这些灌木的生物量模型,不仅节省了人力物力,还有效地保护了林下微环境。本研究拟合的13个物种灌木生物量模型,涵盖了金沟岭林场出现频率较高的绝大多数物种。本研究所拟合的生物量模型仅适用于长白山区金沟岭林场林下灌木,若在研究区以外运用该模型需要通过验证,且模型中自变量超过该研究范围时也需经过验证,特别是地径较大的茶藨子。同时,各器官生物量分量总和与单一物种总生物量之间存在一定的偏差,即各器官生物量模型与总生物量模型的相容性有待进一步研究。
 DownLoad:
DownLoad: