







-
毛竹Phyllostachys edulis广泛分布于中国长江以南的丘陵低山山麓地带。毛竹用途广泛,经济、生态和社会价值高,是中国南方主要的经济林种之一。毛竹有其独特的生长方式,即“爆发式生长”,亦称“快速生长”。长期以来,不同学者们对毛竹快速生长期进行了细致的研究,从出笋期间各器官激素分布的研究[1],到光合色素与光合酶活性的变化[2],再到毛竹叶片的生理特性[3]。光合固碳方面,杨迪蝶等[4],许大全等[5],施建敏等[6]也从各个方面进行了细致的研究。针对快速生长期不同时期的气体交换,老竹对新竹营养传输的研究却鲜见报道,只有李洪吉等[7]对毛竹快速生长期间的光合固碳进行了研究。本研究通过测定毛竹快速生长期间不同竹龄毛竹的气体交换特征,试图为揭示毛竹快速生长的固碳机制提供理论依据。
HTML
-
试验研究地位于浙江省杭州市临安区东部的青山街道研口村(30°14′N,119°42′E)的浙江农林大学毛竹生理生态定位监测站。此地属于亚热带季风气候,温暖湿润,日照充足,降水充沛,四季分明。年平均气温为15.9 ℃,极端最高气温为41.2 ℃,最低气温达-13.1 ℃,全年降水量为1 427.0 mm,全年日照时数为1 920.0 h,无霜期为234.0 d。属低山丘陵区,海拔为48~49 m。土壤属棕红壤,土层深度均在60 cm以上。竹龄为1~6年生,竹高为9~16 m,胸径8~16 cm。
-
在试验研究地内搭建实验观测塔。选取观测塔周边自然生长的3年生和5年生毛竹分别进行测定。测定部位为离地高5~7 m的林冠中层。
-
使用Li-6400XT光合仪,从2016年3月下旬至5月中旬,在毛竹快速生长的前期(3月下旬),中期(4月中下旬)和后期(5月中旬),选择晴朗天早上(8:30-10:30),选取3年生和5年生毛竹林冠层中层向南生长的叶片测定光响应曲线。同期选取毛竹中层向南生长的叶片,选择晴朗天,在自然条件下测定净光合速率(Pn)和蒸腾速率(Tr)等主要生理指标的日变化特征,日变化隔1 h测定1次,取值3~5个·次-1。
-
对不同竹龄毛竹的气体交换情况进行单因素方差分析,对显著性进行检验。数据分析使用SPSS软件。
2.1. 试验材料
2.2. 试验方法
2.2.1. 光响应曲线的测定和日变化特征的测定
2.2.2. 数据分析
-
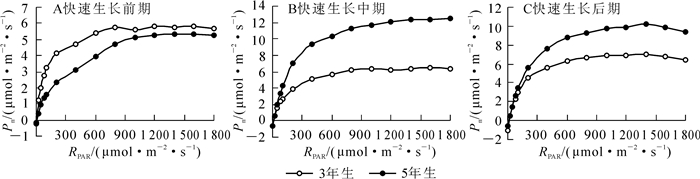
快速生长前期(图 1A),在光合有效辐射(RPAR)0~200 μmol·m-2·s-1之间3年生毛竹叶片净光合速率(Pn)迅速提高,在RPAR为200 μmol·m-2·s-1时,叶片Pn达4.150 7 μmol·m-2·s-1,之后随着光照强度的增加,Pn也随之升高,并在光照强度为800 μmol·m-2·s-1时达到光饱和状态,此时的Pn为5.775 5 μmol·m-2·s-1;而5年生毛竹的光合能力略弱于3年生毛竹,在光照强度为200 μmol·m-2·s-1时,5年生毛竹的Pn为2.369 5 μmol·m-2·s-1,相比于3年生毛竹低了约40%。在光照强度为1 200 μmol·m-2·s-1时,5年生毛竹叶片达到光饱和状态,峰值为5.258 1 μmol·m-2·s-1,低于3年生毛竹约9%。

Figure 1. Light response curve of Phyllostachys edulis in different stages of rapid growth period
快速生长中期(图 1B),3年生毛竹与5年生毛竹光合能力差距明显。3年生毛竹在光照强度为1 000 μmol·m-2·s-1时达到光饱和状态,峰值为6.327 2 μmol·m-2·s-1,而5年生毛竹在光照强度为1 200 μmol·m-2·s-1左右时达光饱和状态,峰值为12.080 2 μmol·m-2·s-1,比3年生毛竹高出了近91%。
在快速生长后期(图 1C),5年生毛竹的Pn依然高于3年生毛竹。在光照强度为1 200 μmol·m-2·s-1时,5年生毛竹的Pn为9.819 5 μmol·m-2·s-1,比3年生毛竹的6.934 0 μmol·m-2·s-1高出近30%。
-
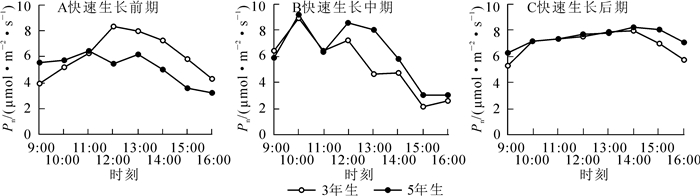
从图 2可以看出:3年生毛竹在快速生长前期(图 2A)Pn的变化规律呈“单峰型”曲线,不具有“光合午休”现象,峰值出现在13:00(8.362 1 μmol·m-2· s-1),而后Pn迅速下降。进入快速生长中期后(图 2B),Pn日变化呈“三峰型”曲线,峰值分别出现在10:00(8.920 3 μmol·m-2·s-1),12:00(7.245 7 μmol·m-2·s-1)和14:00(4.737 4 μmol·m-2·s-1),而在快速生长的后期(图 2C),Pn的变化与前期相似,呈“单峰型”曲线,峰值出现在14:00(7.925 5 μmol·m-2·s-1)。而5年生毛竹在快速生长前期则呈“双峰型”曲线,有明显的“光合午休”现象。上午毛竹叶片Pn随着光照强度的上升逐渐上升,11:00时Pn达到第1个峰值(6.481 7 μmol·m-2·s-1),随后开始出现明显的午休现象,而第2个峰值出现在13:00(6.190 9 μmol·m-2·s-1)。进入快速生长中期之后,毛竹叶片光和能力明显增强,在10:00达到第1个峰值(9.229 4 μmol·m-2·s-1),光合午休现象出现的时间更早,也更为明显,第2个峰值出现在12:00(8.614 6 μmol·m-2·s-1),随后光合速率明显下降。在快速生长后期,5年生毛竹Pn日变化与3年生毛竹相似,呈“单峰型”曲线,峰值出现在14:00左右(8.226 7 μmol·m-2·s-1),而后Pn迅速下降。将3年生毛竹和5年生毛竹的Pn与不同的生长时期进行方差分析发现(表 1),3年生毛竹不同时期变化差异不显著,而5年生毛竹Pn在前期显著低于中期和后期。将毛竹Pn与竹龄和快速生长的3个时期进行相关性分析(表 2)发现,这2个因素对毛竹叶片光合作用的影响均达极显著程度(P<0.01)。

Figure 2. Net photosynthetic rate diurnal variation of Phyllostachys edulis in different stages of rapid growth period
项目 前期 中期 后期 3年生毛竹 5.593 9 a 5.619 1 a 6.206 6 a 5年生毛竹 5.692 6 a 6.490 8 b 6.735 9 b 说明:不同字母表示均值差达显著水平(P < 0.05) Table 1. ANOVA between net photosynthetic rate and growth period in different ages of bamboo
项目 Pearson相关性 显著性(双侧) 样本数/个 生长期 0.164** 0.001 416 竹龄 0.132** 0.007 416 说明:**表示在0.01(双侧)水平上相关 Table 2. Correlational analysis of the relations between net photosynthetic rate and bamboo age and growth period
-
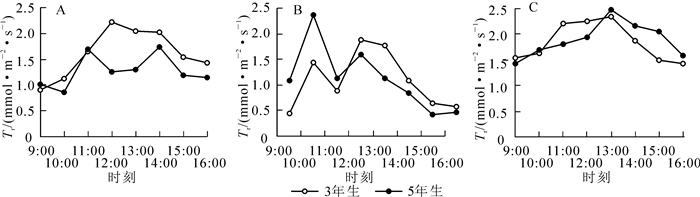
从图 3可以看出:与Pn日变化趋势相似,3年生毛竹在快速生长前期(图 3A)Tr日变化呈“单峰型”曲线,蒸腾强度随着时间的推移不断上升,在12:00达到峰值(2.240 5 mmol·m-2·s-1);中期(图 3B)Tr呈“双峰型”曲线,峰值分别出现在10:00(2.359 6 mmol·m-2·s-1)和12:00(1.590 3 mmol·m-2·s-1);到了快速生长后期(图 3C),Tr曲线重归“单峰型”,峰值与快速生长初期相似,出现在13:00(2.352 0 mmol·m-2·s-1)。而5年生毛竹蒸腾速率曲线与3年生毛竹略不同,在生长前期即呈“双峰型”曲线,峰值分别出现在11:00(1.713 1 mmol·m-2·s-1)和14:00(1.756 8 mmol·m-2·s-1);进入中期后,蒸腾速率日变化曲线同样为“双峰型”曲线,但峰值比前期略早,分别在10:00(1.442 3 mmol·m-2·s-1)和12:00(1.869 6 mmol·m-2·s-1)出现;快速生长后期蒸腾速率日变化曲线为“单峰型”,峰值在13:00(2.480 3 mmol·m-2·s-1)出现。在整个生长期间,蒸腾速率在中期最弱,后期最强,前期居中,且在中前期,3年生毛竹高于5年生毛竹,后期5年生毛竹高于3年生毛竹。对Pn和Tr以及生长时期和竹龄进行相关性分析发现(表 3),Pn和Tr趋势相同,且存在极显著的正相关关系(P<0.01),生长时期的变化对Tr的影响也达到了极显著程度(P<0.01)。竹龄对Tr的影响则没有达显著程度(P>0.05)。

Figure 3. Transpiration rate diurnal variation of Phyllostachys edulis in different stages of rapid growth period
项目 Pearson相关性 显著性(双侧) 样本数/个 净光合速率 0.688** 0.000 416 生长期 0.201** 0.000 416 竹龄 0.056 0.255 416 说明:**表示0.01水平(双侧)上极显著相关,*表示0.05水平(双侧)上显著相关 Table 3. Correlational ana1ysis of the re1ations between transpiration rate and net photosynthetic rate, growth period, and bamboo age
-
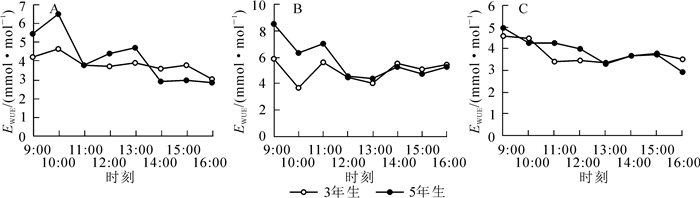
水分利用效率(EWUE)表示植物每消耗单位含水量而同化的二氧化碳的量,通过公式EWUE=Pn/Tr可计算出毛竹叶的水分利用效率(EWUE)[8]。从图 4可以看出:3年生毛竹在前期(图 4A)EWUE变化比较平滑,其峰值出现在10:00(4.649 5 mmol·mol-1),中午过后EWUE趋于稳定;而5年生毛竹EWUE变化起伏较大,峰值出现在10:00(6.490 8 mmol·mol-1)和13:00(4.691 9 mmol·mol-1)。到快速生长中期(图 4B),毛竹EWUE波动明显,3年生毛竹的峰值分别在9:00(5.942 5 mmol·mol-1),11:00(5.666 5 mmol·mol-1),14:00(5.571 4 mmol·mol-1)和16:00(5.565 3 mmol·mol-1)出现;而5年生毛竹在快速生长中期EWUE趋势与3年生毛竹相同,峰值同样出现在9:00(8.556 1 mmol·mol-1),11:00 (7.145 5 mmol·mol-1),14:00(5.309 1 mmol·mol-1)和16:00(5.335 6 mmol·mol-1);在快速生长后期(图 4C),2个竹龄的毛竹EWUE曲线趋势亦相似,峰值出现在9:00(3年生毛竹为4.589 8 mmol·mol-1,5年生毛竹为4.999 6 mmol·mol-1)和15:00(3年生毛竹为3.778 5 mmol·mol-1,5年生毛竹为3.750 3 mmol·mol-1)。通过将不同时期不同竹龄的毛竹EWUE进行方差分析发现(表 4),3年生毛竹在快速生长前期和后期与5年生毛竹在快速生长前期和后期的EWUE没有显著差异。3年生毛竹在快速生长中期的EWUE显著高于3年生毛竹快速生长前期(约46%),快速生长后期(约47%),5年生毛竹快速生长前期(约40%)和快速生长后期的EWUE(约36%),5年生毛竹在快速生长中期的EWUE又显著高于同时期的3年生毛竹(约19%)。将3年生毛竹与5年生毛竹在上午(12:00及之前)与下午(12:00之后)的EWUE进行方差分析(表 5)后发现:毛竹在上午的EWUE极显著高于下午(P<0.01)。5年生毛竹在上午(12:00及之前)的EWUE极显著高于3年生毛竹,下午(12:00之后)2个竹龄毛竹的EWUE差异不显著。

Figure 4. Water use efficiency diurnal variation of Phyllostachys edulis in different stages of rapid growth period
3年生毛竹EWUE 5年生毛竹EWUE 前期 中期 后期 前期 中期 后期 3.808 8 a 5.593 4 b 3.786 2 a 3.973 8 a 6.662 6 c 4.106 7 a 说明:不同字母表示均值差达显著水平(P < 0.01) Table 4. ANOVA with water use efficiency in different stages of rapid growth period of different ages of bamboo
3年生毛竹EWUE 5年生毛竹EWUE 上午(12:00及之前) 下午(12:00之后) 上午(12:00及之前) 下午(12:00之后) 4.650 4 a 3.754 2 b 5.784 0 c 3.738 5 b 说明:不同字母表示均值差达显著水平(P < 0.01) Table 5. ANOVA with water use efficiency between morning and afternoon of rapid growth period of different ages of bamboo
3.1. 快速生长期母竹的光响应曲线比较
3.2. 快速生长期母竹气体交换日变化特征的比较
3.2.1. 不同生长期不同竹龄光合作用日变化的比较
3.2.2. 不同生长期不同竹龄母竹蒸腾速率(Tr)的日变化比较
3.2.3. 水分利用效率(EWUE)日变化
-
快速生长是毛竹特有的生理特征,毛竹林有大小年之分,大年促进植物生长或分裂的吲哚乙酸(IAA),赤霉酸(GA3)和激动素(KT)含量高[1],出笋;小年抑制植物生长的脱落酸(ABA)在竹叶、竹竿中的含量高[1, 9],换叶。
已有研究表明:毛竹冬笋在蛋白质、氨基酸、脂肪以及糖类等营养物质的含量上均高于春笋[10];毛竹从竹笋长成到成竹的过程中,竹笋不仅消耗冬笋期储存的营养[10],毛竹母竹也会通过竹鞭将其器官中的非结构性碳水化合物传输给生长中的竹笋[11];毛竹林的碳通量为负值,且4月二氧化碳通量绝对值最大[12]。这说明在快速生长期间,毛竹母竹会大量同化二氧化碳,并向新竹输送营养和水分[13],而且这个水分和营养的输送在快速生长中期尤为明显[14]。本研究通过实测测得毛竹快速生长不同时期的净光合速率(Pn)和蒸腾速率(Tr),再经过换算得出叶片水分利用效率(EWUE),通过比较不同竹龄,不同时期毛竹叶片的气体交换能力,深刻认识毛竹林快速生长的生理动态、毛竹林固碳特征以及碳汇估算等。
本研究测定毛竹在快速生长不同时期光响应曲线。结果表现为:快速生长前期,3年生毛竹光合能力强于5年生毛竹;快速生长中后期,5年生毛竹光合能力强于3年生毛竹。这与李洪吉等[7]的研究结果——快速生长前期,光合能力Ⅱ度竹强于Ⅲ度竹强于Ⅰ度竹,而在中期和后期,Ⅰ度竹强于Ⅱ度竹强于Ⅲ度竹——有一定出入。可以看出:在快速生长前期,本研究与李洪吉等的论文结果相同,认为中年竹的光合能力强于老年竹;但在快速生长中后期,本研究的结果却与李洪吉等[7]的结果完全相反。造成这一结果的原因可能是因为李洪吉等对毛竹光响应曲线的测定是完成在快速生长小年,而本研究则是测定了快速生长大年毛竹母竹的光响应曲线变化。毛竹大年出笋,小年换叶的生长规律造成了2项研究在结果上的出入。
通过净光合速率(Pn)与蒸腾速率(Tr)计算出的毛竹叶片水分利用效率(EWUE)可以看出,5年生毛竹在整个快速生长期期间的水分利用效率均要高于3年生毛竹,尤其在快速生长中期。这说明无论是在竹笋刚刚破土而出的快速生长前期,还是在新竹开始其成竹生长的快速生长中期,或是新竹生长趋于停止的快速生长后期,毛竹母竹对新竹的营养供给都是由5年生毛竹发起,经由3年生毛竹而传输给新竹。可以认为,在快速生长前期竹笋刚刚破土而出还没开始其成竹生长的时候,由于冬笋期储存了大量营养物质供其消耗,母竹对新竹的营养和水分传输还不明显,因此,5年生毛竹的Pn要低于3年生毛竹;到了快速生长中期,新竹开始其成竹生长,对养分和水分的消耗量加大,便借由水势梯度[12]向母竹汲取营养和水分。因此,3年生毛竹和5年生毛竹都为了满足新竹的需要调整了其生存策略,加强了自身的光合速率,同时提高了其水分利用效率。在生长中期,3年生毛竹的EWUE高出快速生长前期和后期40%左右,而5年生毛竹则高出3年生毛竹约20%。
结合李洪吉等[7]和蔡先锋等[15]对毛竹快速生长期光合固碳特征及耗水特性做出的研究结果,袁佳丽等[14]对毛竹快速生长期的水势变化特征以及SONG等[11]、徐超等[12]对毛竹成竹快速生长时间的界定,可以认为,成竹通过竹鞭向新竹输送养分和水分的这个过程在快速生长中期最为剧烈,5年生毛竹是对新竹的营养传输的源头,是新竹快速生长最主要的养分来源。
 DownLoad:
DownLoad: