








-
林木竞争指生长在同一生境中的林木为争夺共需的有限资源而阻碍其他林木正常生长发育,甚至致使其死亡的行为[1-3]。林木竞争导致个体间差异,进而动态调节林分结构及其空间关系,决定林木竞争格局和空间生态位[4],从而推动群落演替。林木竞争研究能为调节林分结构和保护濒危物种提供依据,有利于维持生物多样性和生态系统的稳定性[1, 5-6],一直是生态学研究的核心内容之一。竞争指数的提出为定量分析林木竞争强度提供了途径,根据其与距离的关系分为完备型和非完备型竞争指数[7]。非完备型竞争指数主要有树高断面积竞争指数、树冠表面积外露竞争指数等[8],仅与对象木信息或周围环境信息有关;完备型竞争指数充分结合对象木信息与周围环境信息,主要有基于树冠面积重叠的Bella竞争指数[9]、Hegyi竞争指数[10]和基于交角的竞争指数[11]等。其中,Hegyi竞争指数及其改进模型因简单易算,适应性较强而被广泛运用[1, 3, 12-15],但该指数却未考虑冠幅和树高在竞争中的影响;而树冠面积重叠竞争指数和基于交角的竞争指数则分别涉及了冠幅与树高的影响,亦被广泛运用[16-18]。油松Pinus tabuliformis是中国北方重要造林树种[19],第八次北京市园林绿化资源普查成果报告显示,按优势树种划分,北京油松林面积和蓄积量分别约8.45万hm2和213.19万m3,占北京森林面积与蓄积量的16.76%和15.16%,是北京森林生态系统的重要组成部分。因经营管理不到位,油松林单位面积蓄积量仅为25.22 m3·hm-2,低于北京森林平均水平(27.88 m3· hm-2);林分现存结构不合理,林木竞争激烈,天然更新困难,林分质量、生物多样性和稳定性均较低[20],亟需开展科学的经营管理。为改善油松人工林结构,首先应筛选一种更适合油松的林木竞争模型来了解该区域油松人工林林内竞争机制,为后期结构调整提供基础数据和理论支撑。如何在众多模型中选用最适模型来量化林分林木竞争强度一直是研究的重点[11, 21]。目前油松林竞争研究多为竞争模型的构建及改进、种内与种间竞争关系分析、林分结构与生物多样性探讨等[14, 19-20, 22],鲜有不同类型竞争模型对油松的适用性评价。基于此,本研究以京西九龙山油松人工林为对象,采用回归分析及k折交叉验证法[23],开展不同类型竞争指数模型对油松的适用性评价研究,筛选出更适于油松人工林的竞争模型,试图初步揭示其竞争机制,为后期经营管理提供依据。
HTML
-
研究区位于北京市门头沟九龙山(39°54′~39°59′N,115°59′~116°07′E),该区属暖温带大陆东岸半湿润大陆性季风气候,年平均气温为11.8 ℃,年平均降水量为623 mm,主要集中在6-9月,年平均蒸发量为1 870 mm,无霜期为216 d。土壤属于山地褐土,土层普遍较薄,含石量高[24]。研究区于20世纪60年代开始封山育林,同期选用2~3 a油松幼苗进行大面积的鱼鳞坑整地造林,油松人工林初始密度为5 000株·hm-2。在海拔100~997 m分布着典型落叶阔叶林和温性针叶林[24],主要树种有油松,栓皮栎Quercus variabilis,侧柏Platycladus orientalis,白蜡树Fraxinus chinensis,榆树Ulmus pumila,北京丁香Syringa pekinensis,朴树Celtis sinensis,臭椿Ailanthus altissima,杜梨Pyrus betulifolia和桑Morus alba等。
-
于2017年6月,在研究区内设置1个100 m × 100 m油松人工林样地(39°55′08″N,116°01′18″E)。该样地平均海拔750 m,坡度18°,北坡,林龄约53 a。对样地内胸径大于5 cm的林木进行每木检尺,记录树种名称,用围径尺测量胸径、用皮尺测量冠幅、用激光测高仪测量树高和枝下高,并用TOPCON全站仪进行定位。样地基本概况见表 1,林木冠幅及位置分布见图 1。
郁闭度 公顷断面积/(m2·hm-2) 蓄积量/m3 密度/(株·hm-2) 树种数 胸径/cm 树高/m 冠幅/m 0.80 26.88 113.58 1 159 13 16.39 ± 5.15 7.80 ± 1.79 3.25 ± 1.08 Table 1. Stand survey factors

Figure 1. Crown width and position distribution of the trees in plots
-
采用8邻域大样地平移法进行边缘校正[15],将原样地内所有油松作为对象木,并与其4株最近相邻木构成最佳空间结构单元[8]。
-
采用裴雅茹[25]的北京油松生物量研究模型对样地内油松生物量进行估算,公式为:
式(1)中:W为生物量(kg),D为林木胸径(cm),H为树高(m)。以Dj为竞争木j的胸径(cm),Di为对象木i的胸径(cm),dij为对象木i与竞争木j之间的距离(m),n为竞争木数量,得到Hegyi竞争指数[10]:
式(2)中:CHg_i为Hegyi竞争指数。Bella竞争指数(CIOi)[9]公式为:
式(3)中:Co_i为Bella竞争指数,ZO_ij为对象木i与竞争木j树冠投影重叠面积(m2),ZA_i为对象木i的树冠投影面积(m2)。基于交角的林木竞争指数(d_α_CIi)公式为:
式(4)~(6)中:Cd_α_i[17]为总竞争指数,Cd_α_i上和Cd_α_i侧则分别为上方遮盖和侧方挤占;α1和α2为对象木与竞争木之间的高度交角(°),如果相邻木j高于对象木i(图 2A),α1=arctg(Hi/dij),α2=arctg[(Hj-Hi)/dij],cij=1;否则(图 2B)α1=arctg(Hj/dij),α2不存在,cij=0。对象木的各竞争木(n=4)竞争强度的均值为竞争指数,在竞争指数计算公式(2),(3),(5),(6)中,竞争木j的竞争强度可描述为:

Figure 2. Schematic diagram of the over shading and the lateral extruding
-
本研究的油松人工林为同龄林,油松生物量可较好反映油松所受竞争强度。为筛选更适合油松人工林的竞争指数,分别采用指数、对数、多项式、线性和幂函数等模型作油松生物量与该3类竞争指数的回归分析,获取决定系数(R2)最大的最优模型,线性转换后,进行n次k折交叉验证(n和k均为10),平均n次k折交叉验证的R2作为最终的验证R2。k折交叉验证,即将样本分为分割成k个子样本,轮流将(k-1)个子样本组合作为训练集,另外的1个样本作为验证模型的保留集,k次的平均交叉验证识别正确率即为模型检验结果[23]。k折交叉验证的验证R2较回归R2损失得越少,模型泛化能力越强,以此结合决定系数(R2)来筛选竞争指数,并用该竞争指数对林分现状作进一步分析。
-
采用SVMS 1.0软件对采集的油松人工林数据进行预处理,获取对象木与4株竞争木所构竞争单元基本信息,通过Excel 2010完成各对象木生物量和竞争指数的计算,并运用R软件编程获取生物量与竞争指数间的最优模型、决定系数(R2)及n次k折交叉验证R2,在SPSS 20.0软件中完成对象木径级和竞争指数的相关分析。
2.1. 样地调查
2.2. 数据分析方法
2.2.1. 对象木和竞争木的确定
2.2.2. 生物量模型及竞争模型

2.2.3. 模型适用性评价及油松人工林竞争现状分析
2.2.4. 数据分析
-
样地内胸径5 cm以上林木共1 159株13种,其中油松对象木1 054株。油松对象木径级分布为正态分布(偏度0.093,峰度-0.197;P=0.373>0.05),正态分布曲线略正偏。油松对象木及竞争木概况如表 2所示。油松对象木和竞争木胸径主要集中在10~25 cm,分别占88.24%和85.08%,其中5~10 cm径级的油松对象木数量较少,仅占8.16%。除大于等于30 cm的径级外,同径级对象木平均胸径大于竞争木,但同径级对象木平均树高皆小于等于竞争木。
径级/cm 对象木 竞争木 株数/株 百分比/% 平均胸径/cm 平均树高/m 株数/株 百分比/% 平均胸径/cm 平均树高/m 5~10 86 8.16 8.36 5.48 489 11.60 8.03 5.52 10~15 298 28.27 12.82 7.07 1 266 30.03 12.76 7.15 15~20 399 37.86 17.53 8.17 1 536 36.43 17.47 8.17 20~25 233 22.11 22.01 8.95 785 18.62 21.98 8.95 25~30 33 3.13 26.57 9.47 100 2.37 26.45 9.49 ≥30 5 0.47 31.48 10.80 40 0.95 32.10 11.23 合计 1 054 100.00 4 216 100.00 Table 2. The status of objective tree of Pinus tabuliformis and competitor
-
林木生物量累积速率受竞争强度影响,所受竞争强度越小,其生物量累积速率越快[18]。
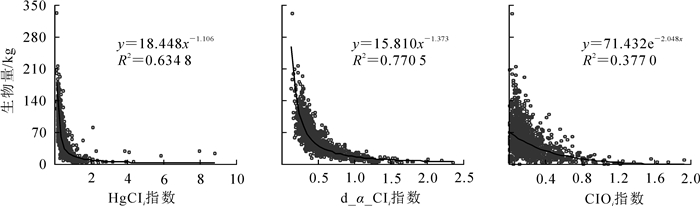
采用回归分析法对油松生物量与其竞争指数进行分析,结果表明(表 3):Hegyi竞争指数和d_α_CIi的最优模型为幂函数,CIOi的最优模型则为指数函数,且均通过模型的显著性检验(P<0.01)。10次10折交叉验证结果显示:各模型的交叉验证R2较决定系数R2损失均小于0.005;其中,d_α_CIi与生物量决定系数R2最高,R2损失量最小,分别为0.770 5和0.001 0,Hegyi竞争指数次之,CIOi最差。各竞争指数与生物量的关系如图 3所示,油松生物量均随各竞争指数的增大而减小;其中,CIOi和Hegyi竞争指数较d_α_CIi存在较多的离群点。综上所述,d_α_CIi能更好反映油松所受竞争强度。
竞争指数 最优模型 决定系数R2 验证R2 ΔR2 P HgCIi W=18.448x-1.106 0.634 8 0.631 2 0.003 6 <0.01 d_α_CIi W=15.810x-1.373 0.770 5 0.769 5 0.001 0 <0.01 CIOi W=71.432e-2.048x 0.377 0 0.373 4 0.003 6 <0.01 说明:ΔR2=决定系数R2-验证R2;W代表对象木生物量,x代表竞争指数 Table 3. Biomass response model of objective tree and cross-validation results under different competition indices

Figure 3. Correlation analysis of different competition indices with biomass
-
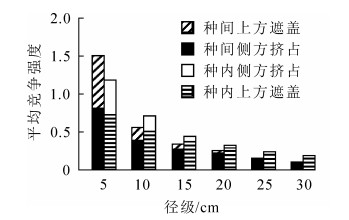
从图 4可知:小径级油松受侧方挤占和上方遮盖比例相差不大,随着径级的增大,侧方挤占逐渐主导竞争,大径级油松几乎不受上方遮盖影响。油松所受平均竞争强度随径级增大而减少,上方遮盖随径级的增大而迅速减小。除5~10 cm径级外,油松种间平均竞争强度均小于种内。对油松对象木的竞争指数作径级分析,因其径级竞争指数未满足方差齐性检验,故选用非参数检验进行分析。非参数检验的Kruskal-Wallis检验结果表明油松径级间竞争指数存在显著差异(P<0.05),由图 5可知:油松胸径>20 cm,各径级间差异均不显著,具体表现为20~25 cm与25~30 cm,20~25 cm与≥30 cm及25~30 cm与≥30 cm径级间差异不显著,其余各径级间差异均显著(调整后P<0.05)。这表明油松胸径<25 cm时,其竞争指数随胸径的增大而显著减少,但胸径≥20 cm的各径级油松个体竞争指数随径级变化不显著,占油松总体的25.71%。

Figure 4. Side and above competition varies with diameter class of objective tree

Figure 5. Competition index of objiective tree of P. tabuliformis varies with diameter class
-
从表 4可见:竞争木胸径越大,其竞争强度越大,上方遮盖占总竞争强度的比例也越大。当竞争木胸径为15~20 cm时,其种内、种间竞争强度最大,分别为868.87和32.26,种内竞争远大于种间。种内竞争的竞争木株数、竞争强度随径级呈先增大后减小的趋势,而种间竞争的竞争木株数、竞争强度总体上随径级呈先减小后增大的趋势,总体上各径级的种间竞争木株数和竞争强度远小于种内。
径级/cm 竞争来源 种内竞争 种间竞争 株数/株 竞争强度 合计 平均竞争强度 株数/株 竞争强度 合计 平均竞争强度 5~10 侧方挤占 342 75.31 83.12 0.24 147 23.67 24.97 0.17 上方遮盖 47 7.80 10 1.30 10~15 侧方挤占 1 176 375.33 448.50 0.38 90 26.15 32.12 0.36 上方遮盖 377 73.18 34 5.97 15~20 侧方挤占 1 483 663.86 868.87 0.59 53 23.04 32.26 0.61 上方遮盖 800 205.01 34 9.22 20~25 侧方挤占 771 422.68 593.64 0.77 14 8.05 11.57 0.83 上方遮盖 541 170.96 9 3.51 25~30 侧方挤占 99 68.75 104.89 1.06 1 0.66 1.16 1.16 上方遮盖 82 36.13 1 0.50 ≥30 侧方挤占 13 9.97 17.02 1.31 27 23.41 41.55 1.54 上方遮盖 11 7.05 27 18.14 总计 2 116.03 143.63 Table 4. Distribution of the intraspecific and interspecific competition intensity according to diameter class of competitive tree
-
由表 5可见:油松种内竞争强度为2 116.03,占93.64%,远大于种间,这与其多为纯林的特性相符。油松人工林种内及主要伴生种间的竞争强度顺序从大到小依次为油松种内,白蜡,榆树,北京丁香,朴树,杜梨,臭椿,桑,槐Sohpora japonica,栓皮栎,山杏Armeniaca sibirica,山楂Crataegus pinanatifida,白梨Pyrus bretschneideri。
物种 竞争来源 株数/株 竞争强度 总竞争强度 平均竞争强度 竞争强度排名 油松Pinus tabuliformis 侧方挤占 3 884 1 615.89 2 116.03 0.54 1 上方遮盖 1 858 500.13 白蜡树Fraxinus chinensis 侧方挤占 65 20.24 26.26 0.40 2 上方遮盖 20 6.03 榆树Ulmus pumila 侧方挤占 21 13.34 24.54 1.17 3 上方遮盖 18 11.20 北京丁香Syringa pekinensis 侧方挤占 63 17.70 22.43 0.36 4 上方遮盖 13 4.73 朴树Celtis sinensis 侧方挤占 77 16.89 19.47 0.25 5 上方遮盖 18 2.58 杜梨Pyrus betulifolia 侧方挤占 17 10.68 17.07 1.00 6 上方遮盖 10 6.40 臭椿Ailanthus altissima 侧方挤占 38 9.62 12.14 0.32 7 上方遮盖 18 2.53 桑Morus alba 侧方挤占 24 8.41 11.79 0.49 8 上方遮盖 9 3.38 槐Sophora japonica 侧方挤占 3 1.65 2.79 0.93 9 上方遮盖 3 1.14 栓皮栎Quercus variabilis 侧方挤占 12 2.26 2.62 0.22 10 上方遮盖 3 0.37 山杏Armeniaca sibirica 侧方挤占 3 1.40 1.65 0.55 11 上方遮盖 2 0.25 山楂Crataegus pinnatifida 侧方挤占 5 1.89 1.95 0.39 12 上方遮盖 1 0.06 白梨Pyrus bretschneideri 侧方挤占 4 0.90 0.90 0.22 13 上方遮盖 0 0.00 合计 4 216 2 259.66 2 259.66 Table 5. The competition according to species
3.1. 油松对象木与竞争木概况
3.2. 油松人工林不同竞争指数的适用性分析
3.3. 油松对象木的竞争强度分析
3.3.1. 竞争强度随油松对象木胸径的变化

3.3.2. 油松种内、种间竞争随竞争木径级的变化
3.3.3. 油松所受竞争强度随树种的分布规律
-
本研究的油松生物量与d_α_CIi,Hegyi竞争指数服从幂函数关系,与CIOi为指数函数。大部分学者认为Hegyi竞争指数与林木大小参数服从幂函数[1, 3, 11-12, 15, 26],本研究支持了该观点。而惠刚盈等[11]建立的基于交角的竞争指数与林木大小参数间拟合的最优模型却不为幂函数,与本研究结果不同,这可能是本研究选取竞争指数的胸径比参数替换了其所选用的大小比数造成的,不同结构的竞争指数所拟合的最优模型不同。
d_α_CIi,CIOi和Hegyi竞争指数均反映了油松生物量受竞争强度增大而减小的规律,3类竞争指数的R2损失量均较小,说明其回归模型泛化能力均较强;其中,d_α_CIi的决定系数最高,损失量最少,与生物量拟合的离群点较少,说明其对油松所受竞争强度的量化稳定,能较好处理如多主干油松所受竞争强度等特殊情况。因此,d_α_CIi能更好量化北京典型油松人工林竞争强度。
CIOi表现最差,这可能受油松树冠特征影响。油松人工林多为同龄林,林木大小分化不明显,作为喜阳物种,油松树冠的中下层侧枝易在激烈的竞争中因光环境过弱而枯死,而其上层侧枝因生长年限不足而偏短,从而导致其冠幅较小,树冠重叠面积偏少,所计算竞争指数可能低于林木实际所受竞争强度。且林木间剧烈的竞争易导致树冠的可塑性变异[27],令测量的冠幅大小和空间位置的准确性偏低,从而使竞争强度量化不够准确。因此,该竞争指数表现较差。
北京地区油松人工林因长期缺乏经营管理,其林分密度过大,林木间距不足,光环境过弱,为提高竞争力获取更多光照,林木将更多的能量用于树高生长[28]。因此,北京油松人工林中林木树高是竞争力的重要表现形式。本研究中选取的基于交角的竞争指数d_α_CIi优于Hegyi竞争指数,这与惠刚盈等[11]建立的基于交角的林木竞争指数优于Hegyi竞争指数的结果相似,可能树高与间距比构成的交角因子更好地反映了林木在水平和垂直方向上的竞争压力,所以竞争指数d_α_CIi效果更为理想。
不同竞争指数侧重点和适用性不同[26],d_α_CIi在量化油松纯林竞争强度上表现较好,能具体分析其上方遮盖和侧方挤占的竞争强度,但是否对油松混交林也能保持较高的适用性有待进一步深入研究。
-
油松对象木径级正态分布曲线略正偏,这主要是林木对光等有限资源的不对称竞争引起的[29]。在温带森林里竞争压力通常是阻碍林木生长甚至导致其死亡的主要因素[30],油松人工林过强的竞争压力,致使油松总体胸径偏小,小径级油松受竞争影响不断死亡。而林内过高的郁闭度、过厚的林下凋落叶层[31],可能共同导致了其林下更新困难,小径级林木不断死亡却难以得到补充,林分难以进一步形成近自然的异林龄结构。
采用d_α_CIi对1 054株油松对象木和4 216株竞争木竞争强度的量化研究表明:油松所受竞争强度随其胸径增大而减小,小径级油松受侧方挤占和上方遮盖的强度均较大,且两者比例相差不大,随胸径的增大,其来自上方遮盖的比例逐渐减小甚至消失。当胸径≥20 cm(约油松林中前1/4),油松所受竞争强度随径级的增大变化不显著,这与项小燕等[1]对大别山五针松Pinus dabeshanesis,曾思齐等[3]对青冈栎Cyclobalanopsis glauca,沈琛琛等[26]对蒙古栎Quercus mongolica和邢海涛等[13]对马尾松Pinus massoninana的研究结果相似。这可能是因为小径级油松获取光照、水分等资源的能力较弱,所受压力较大,而大个体油松居于上层林冠,资源获取能力更强,因而受其他林木竞争影响较小,并能有效抑制竞争木生长,从而进一步稳定其竞争优势。除5~10 cm径级外,油松种间平均竞争强度均小于种内,说明油松个体水平种内竞争更激烈。
油松种内竞争强度为2 116.03,占93.64%,所受竞争压力主要来自种内,这与油松林中其他林木数量过少有关。在油松人工林的种内及主要伴生种间的竞争强度排序中,榆树的数量仅约占北京丁香和朴树的1/3,但受林木种类各自生长特征影响,其个体较大,竞争强度普遍较高,进而导致榆树排序反而靠前,说明林木竞争受数量、大小和种类共同影响,这与项小燕等[1]的研究结果相同。
种内竞争的竞争木株数、竞争强度随竞争木径级呈先增大后减小的趋势,而种间竞争的竞争木株数、竞争强度总体上呈先减小后增大的趋势,且各径级的种内竞争木株数和竞争强度总体上远大于种间。这印证了在许多植物群落中种内竞争往往比种间竞争更强是局域尺度上物种稳定共存的重要条件,也是相邻尺度上稳定共存机制运作的普遍规律[32],亦表明油松在竞争中占主导地位,其他林木受其竞争排挤,大多以小径级或大径级的形式在油松林垂直空间的两端获取生存空间,两者形成物种间的空间分离。这种空间分离能减弱种间竞争,阻止或减缓竞争优势物种对竞争劣势物种的竞争排除作用,有利于维持物种共存,促进生物多样性维持[33],这也说明生态位差异在相邻空间尺度上的群落动态中发挥着重要作用[32]。
林分密度过高易导致生物量及生产力下降[34],为改善油松林结构,提高其质量,针对其缺乏幼苗、林分密度和竞争强度过高的现状,应合理稀疏和开辟林窗促使幼苗树种的形成及林木间的空间分离,以降低林木竞争强度、提高生物多样性。基于油松林竞争强度的空间分布对局部竞争过强的林木进行合理稀疏,重点围绕胸径<20 cm的目标树开展择伐释压经营管理措施,使其在中小径级时获得足够的空间、光照、水分等资源,减少竞争木对有限资源的掠夺,促使小径级林木的正常生长和林下更新,以改善油松人工林的结构,提高其生态服务功能。
-
d_α_CIi竞争指数能比其他竞争指数更准确的量化北京典型油松人工林的竞争强度。林木竞争受数量、大小和种类共同影响。油松种内竞争大于种间及两者间的垂直空间分离是林内物种共存的重要条件。基于油松林竞争指数的空间分布,对目标树周围胸径<20 cm的竞争木进行合理抚育间伐,能降低目标树所受竞争强度,改善油松林结构,提高油松林生态系统服务功能。
 DownLoad:
DownLoad: