






-
中国东北地区的针阔混交林是温带森林生态系统的核心组成部分,在维护国家生态安全、碳汇功能和保护生物多样性等方面具有不可替代的重要作用[1]。吉林蛟河针阔混交林是其典型代表,以红松Pinus koraiensis、色木槭Acer mono、紫椴Tilia amurensis、水曲柳Fraxinus mandshurica等乔木为主要树种,凭借复杂的群落结构与功能,为区域气候调节、水土保持提供了重要支撑[2]。针阔混交林是该地区的顶级群落类型。由于20世纪以来的过度采伐等干扰,当地针阔混交林资源被严重破坏,大量原始林退化为次生林,生态功能减弱[3],现仍处于演替早期阶段。现有研究认为:中低强度的采伐能够优化群落结构,有利于群落生产力与稳定性的提升;高强度的采伐会导致生态位的过度释放,对森林谱系多样性和功能多样性产生严重的负面影响[4]。鉴于仅有一定强度范围内的采伐才能有利于森林恢复,因此亟需相关研究为其恢复与可持续发展提供科学的理论支撑。
植物叶片对环境的变化极为敏感,能够通过调整生长策略达到与环境变化相适应的目的,因此叶片功能性状可以反映植株个体获取与保护资源的策略[5]。叶功能性状包括叶光合特性、叶结构性状、叶化学性状等[6]。当前,对于叶功能性状的研究多集中于叶对各种自然环境胁迫的响应。李昕蓉等[7]研究发现:叶结构性状受遗传调控为主,而叶化学性状更多受到环境驱动;刘阳等[8]对4种槭树Acer spp.的研究结果显示:槭树的比叶面积等结构性状随着气候变化而波动;崔晓阳等[9]研究发现:对于水曲柳幼苗,较低的土壤水势导致非气孔因素改变,进而主导了光合抑制现象;郝珉辉等[10]研究发现:吉林蛟河乔木叶性状受坡向、海拔等地形因子的显著影响。关于采伐等人为扰动对不同种间叶功能性状影响的研究相对较少。当前对采伐的研究集中于林分结构、物种组成、幼苗更新等宏观生态功能领域[11],而采伐强度对乔木叶、根等器官功能性状的研究则较为分散。巫志龙等[12]研究发现:对杉阔混交林进行中低强度采伐处理与重度采伐处理后,混交林中乔木细根生物量减少,并在2个处理间存在显著差异;米爽等[13]研究发现:重度采伐会导致幼苗的比叶面积显著下降。
国内外研究表明,采伐通过增加辐射有效性减少了各树种对资源的竞争,导致叶片变薄从而提升单位叶面积的光合速率[14]。采伐对叶功能性状的影响会因树种的不同而有所变化。欧洲黑松P. nigra在重度采伐(减少60%~70%胸高断面积)下叶干物质质量分数显著高于中度采伐(减少30%~40%胸高断面积),欧洲赤松P. sylvestris在重度采伐下光合速率显著高于对照组[15],上述研究揭示了采伐干扰对叶功能性状的影响,有助于了解养分循环的响应模式,确定最佳采伐强度并优化其树种配置方案[15]。因此,研究采伐强度对叶功能性状的影响具有重要的理论和实践意义[16]。
本研究以吉林蛟河针阔混交林中色木槭、水曲柳、紫椴和红松等4种主要树种为研究对象,探究不同采伐强度对各树种光合特性叶结构性状的影响,旨在揭示叶功能性状对不同采伐强度的响应,为该地区森林可持续经营提供理论支持,为量化采伐干扰的影响以及采伐后森林恢复管理提供科学依据。
-
研究区域位于吉林省吉林市蛟河市国有林保护中心监测样地(43°57′31″~43°58′03″N, 127°44′07″~127°44′40″E),海拔为430~457 m。该区域属于温带大陆性季风气候,夏季温暖多雨,冬季严寒干燥;年均气温为3.8 ℃,年均降水量为696.0 mm,降水集中于7—8月,冬季降雪显著,年平均积雪厚度为30~65 cm,年均无霜期为130.0 d,年均有霜期为230.0 d。土壤母质主要为花岗岩,林下土壤属于山地暗棕色森林土,平均厚度为30~80 cm,局部可达100 cm以上[17]。监测样地植被类型为典型的天然次生针阔混交林,主要乔木树种包括红松、紫椴、色木槭、水曲柳、春榆Ulmus japonica和蒙古栎Quercus mongolica等[17]。
-
2011年7月,在吉林蛟河针阔混交林成熟林中选取林分状况大体一致的4块面积为1 hm2的固定标准地,使用全站仪将每块样地划分为25个20 m×20 m的样方,调查样地内所有胸径≥1 cm 树木的物种名称、胸径、树高、冠幅和位置坐标等。同年12月,经上级林业主管部门批准后依据间密留匀、留优去劣的原则,根据保留木相邻距离、树种以及林木大小确定采伐木,尽量去除先锋树种或个体数量较多的树种,并将生长状况不良的树木优先去除[18]。根据胸高断面积确定的4种不同采伐强度分别为对照(ck,采伐强度为0)、轻度采伐 (T1,采伐强度为17.24%)、中度采伐(T2,采伐强度为34.74%)、重度采伐(T3,采伐强度为51.85%),获得4块采伐样地,于2013、2015、2018、2021 年对样地进行了复测。样地概况见表1。2023年7月,在4个采伐样地中,选取色木槭、水曲柳、紫椴和红松4种树种作为研究对象,每个树种分别选择胸径、树高、冠幅接近平均水平的3株个体为目标树作为试验重复样。目标树中色木槭平均树高为12.3 m,平均胸径为12.4 cm;水曲柳平均树高为33.0 m,平均胸径为21.7 cm;紫椴平均树高为21.0 m,平均胸径为35.2 cm;红松平均树高为26.0 m,平均胸径为17.5 cm。
采伐强度 海拔/m 坡度/(°) 密度/(株·hm−2) 胸高断面积/(m2·hm−2) 2011年采伐前 2011年采伐后 2021年复测 2011年采伐前 2011年采伐后 2021年复测 ck 453 1 987 987 952 28.875 28.875 31.266 T1 443 4 965 784 903 28.731 24.627 27.849 T2 430 5 947 653 798 29.968 21.151 23.601 T3 497 3 1152 586 655 30.247 15.007 18.433 Table 1. Survey of sample plots
-
2023年7—8月,选取所有晴天7:00—10:30,使用便携式光合作用测定系统(Li-6400XT, Li-COR, 美国)进行光合数据的测定。用高枝剪剪下冠层中部的二级枝后迅速插入装有清水的水桶内进行二次取枝以维持枝条内的水势,确保实验数据准确。每株阔叶样树选取3片完整健康的叶片,每株针叶样树选取4束共20根完整健康的针叶叶片,分别进行光合参数的测定。
使用Li-
6400 光合仪提供的光强为16 000 μmol·m−2·s−1的标准光强红蓝光源,设定相对湿度50%,控制叶室温度为25 ℃,使用外接压缩二氧化碳(CO2)小钢瓶控制CO2摩尔分数为400 μmol·mol−1,气体流速设定为500 μmol·s−1,由强至弱依次设定光量子通量密度为2 000、1 500、1 200、1 000、800、600、400、300、200、150、100、75、50、20、0 μmol·m−2·s−1。每次测量均在2 000 μmol·m−2·s−1光强下诱导30 min后再进行光合参数的测定,每一光强下的测定时间不少于120 s,不多于240 s。测量完毕后将数据导出至电脑,使用非直角双曲线模型[19]对测定的数据进行拟合,获得各树种的光响应曲线,并拟合出最大净光合速率(μmol·m−2·s−1)、暗呼吸速率(μmol·m−2·s−1)、表观量子效率(μmol·m−2·s−1)、光补偿点(μmol·m−2·s−1)和光饱和点(μmol·m−2·s−1)[20]。当叶室光强为1 000 μmol·m−2·s−1,环境CO2摩尔分数为400 μmol·mol−1时,记录4种主要树种的光合气体交换参数,包括净光合速率(μmol·m−2·s−1)、气孔导度(mol·m−2·s−1)、胞间CO2摩尔分数(μmol·mol−1)、蒸腾速率(mmol·m−2·s−1)和水分利用效率(mmol·mol−1)。 -
每株目标树随机选取15片健康完整的叶片,使用手持叶绿素计(SPAD-520PLUS)对红松以外的3个阔叶树种进行叶绿素相对含量(SPAD)的测定,测量时避开主脉,每个取样叶片测定3次并记录,随后立即装入密封袋中。返回实验室后使用分析天平称量阔叶与针叶的鲜质量(g)。使用扫描仪扫描叶片,使用Image J软件,利用像素格比对法计算出叶面积(cm2)。使用游标卡尺测量阔叶叶片厚度(cm),将15片叶片按照相同的方向叠放,测量时避开叶脉,每组叶片测量3次。利用排水法测量红松针叶的叶体积(cm3)。将叶片放入恒温70 ℃的烘箱内72 h后,测量叶干质量(g)。
为探讨每种采伐强度下不同树种的叶片结构性状,分别计算每个样品的比叶重(g·cm−2)、叶干物质质量分数(%)、阔叶叶组织密度(g·cm−3)和针叶叶组织密度(g·cm−3)[19]。
-
用SPSS 26.0对数据进行处理分析,通过单因素方差分析(one-way ANOVA)以及最小显著差异法(LSD)进行不同组间多重比较(P<0.05)。利用GraphPad Prism 9 制作图表。
-
阔叶红松林4种主要树种的光合气体交换参数如表2所示。从光合气体交换参数的种间比较来看:水曲柳的净光合速率在各采伐强度下均显著高于色木槭与红松,且在ck、T2、T3处理下显著高于紫椴(P<0.05),仅在T1处理下与紫椴无显著差异。在所有处理中,水曲柳的气孔导度和蒸腾速率均为最高且显著大于其他树种(P<0.05);紫椴的水分利用效率在T1处理中显著高于其他树种(P<0.05);对于胞间CO2摩尔分数,各采伐强度下普遍没有显著的种间差异,仅有色木槭在T2处理中显著低于水曲柳与紫椴(P<0.05)。
树种 采伐强度 净光合速率/
(μmol·m−2·s−1)气孔导度/
(mol·m−2·s−1)胞间CO2摩尔分数/
(μmol·mol−1)蒸腾速率/
(mmol·m−2·s−1)水分利用效率/
(mmol·mol−1)色木槭 ck 4.67±0.32 Ca 0.06±0.03 Bab 252.61±61.16 Aa 0.84±0.36 Ba 6.13±1.77 Aa T1 5.24±1.60 Ba 0.06±0.02 Bab 247.91±10.32 Aa 1.30±0.47 Ba 4.09±0.35 Ba T2 4.56±0.22 Ba 0.05±0.01 Bb 243.64±32.43 Ba 0.76±0.16 Ca 6.29±1.63 Aa T3 5.91±1.16 BCa 0.13±0.05 Ba 306.98±41.35 Aa 1.49±0.60 ABa 4.63±2.14 Aa 水曲柳 ck 11.22±0.27 Aa 0.20±0.01 Aa 295.74±5.76 Aa 3.03±0.86 Aa 3.87±1.18 ABa T1 8.84±0.92 Ab 0.16±0.07 Aa 281.43±52.43 Aa 2.80±0.93 Aa 3.51±1.43 Ba T2 12.03±0.84 Aa 0.29±0.15 Aa 307.33±36.75 Aa 2.54±0.35 Aa 4.77±0.55 Aa T3 10.94±0.50 Aa 0.24±0.03 Aa 314.74±11.77 Aa 2.09±0.30 Aa 5.35±0.87 Aa 紫椴 ck 4.78±0.61 Cc 0.10±0.02 ABa 310.12±10.67 Aa 2.01±0.33 ABa 2.40±0.24 Bb T1 9.19±0.06 Aa 0.13±0.01 ABa 268.08±7.40 Aa 1.19±0.11 Ba 7.72±0.74 Aa T2 5.46±1.01 Bc 0.10±0.02 Ba 299.53±19.92 Aa 1.75±0.78 ABa 3.81±2.06 Ab T3 7.08±0.53 Bb 0.16±0.06 Ba 311.17±23.44 Aa 2.20±0.60 Aa 3.37±0.73 Ab 红松 ck 6.35±0.54 Ba 0.11±0.06 ABa 279.23±39.75 Aab 1.46±0.68 Ba 5.04±1.88 ABab T1 5.46±0.93 Bab 0.07±0.02 Ba 253.30±32.27 Ab 0.81±0.21 Bb 7.06±1.66 Aa T2 5.25±1.11 Bab 0.08±0.02 Ba 281.77±16.99 ABab 0.97±0.20 BCab 5.49±0.78 Aab T3 4.75±1.12 Cb 0.09±0.04 Ba 306.67±15.67 Aa 1.02±0.40 Bab 4.91±0.79 Ab 说明:数据为平均值±标准差。不同小写字母表示不同采伐强度间差异显著(P<0.05),不同大写字母表示不同树种间差异显著(P<0.05)。 Table 2. Photosynthetically gas exchange parameters of 4 tree species under different logging intensities
从各树种在不同采伐强度间比较来看,色木槭和水曲柳的各项光合气体交换参数普遍较为稳定,仅有色木槭的气孔导度在T2处理中显著小于T3处理(P<0.05),水曲柳的净光合速率在T1处理中显著低于其他处理(P<0.05),其余参数随采伐强度变化不显著。紫椴的气孔导度、胞间CO2摩尔分数和蒸腾速率在各处理间展现出稳定性,而净光合速率与水分利效率均在T1处理中显著高于其他处理(P<0.05),可见,紫椴在面对轻度采伐时通过提升对水分的利用效率提升了其光合能力。红松的各项参数在各处理间差异显著(P<0.05):净光合速率随着采伐强度的增加而下降,T3处理显著小于ck处理;T3处理胞间CO2摩尔分数显著大于T1组,水分利用效率则呈现为T1处理显著大于T3处理(P<0.05); T1处理蒸腾速率显著小于ck处理(P<0.05);气孔导度在各处理间无显著差异。
-
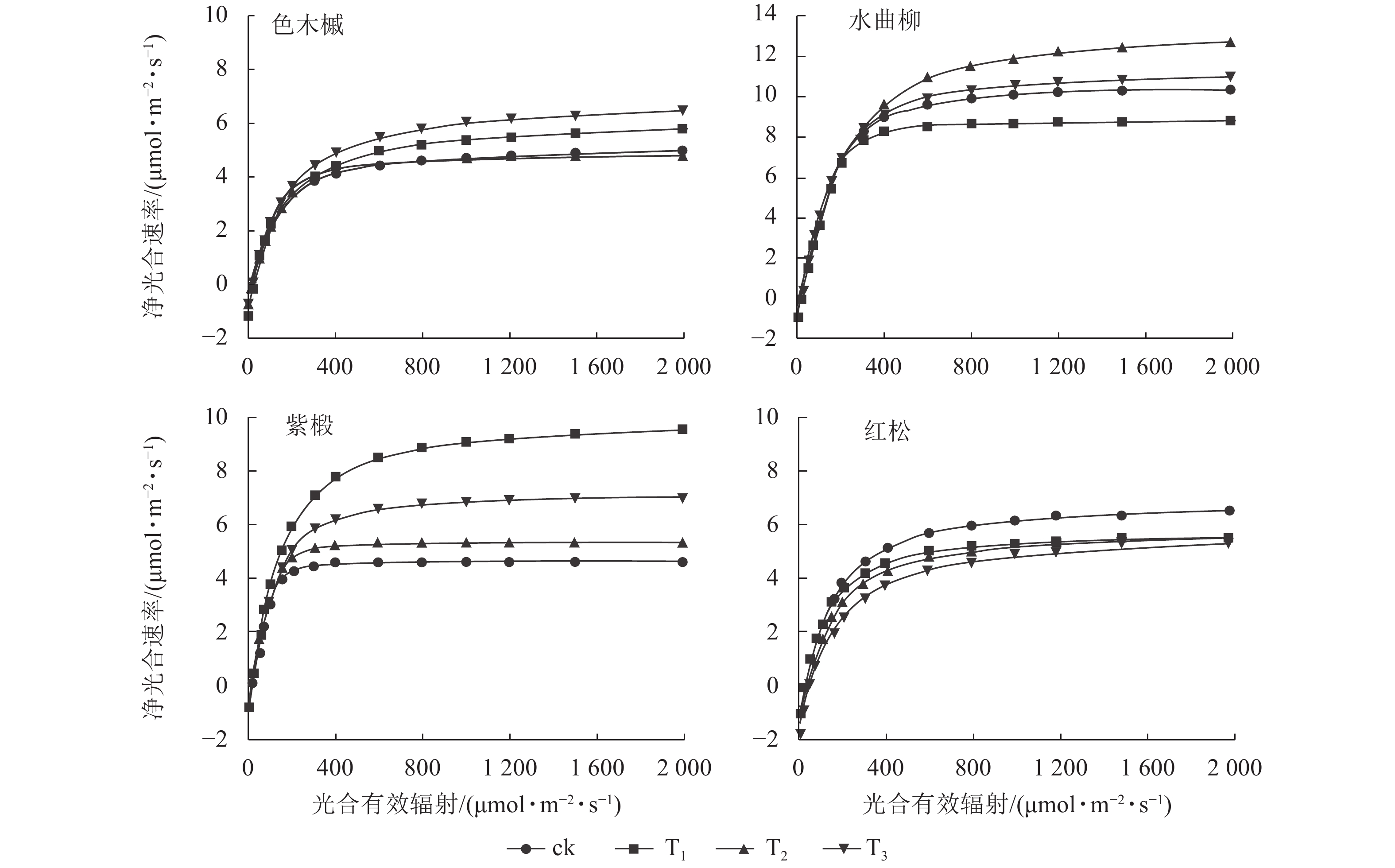
图1显示:随着光合有效辐射的增强,各树种净光合速率变化趋势较为一致。在光合有效辐射为0~200 μmol·m−2·s−1时,各树种的净光合速率均呈快速增长。随着光合有效辐射的增加,净光合速率的增长速率逐步放缓。当光合有效辐射达到光饱和点时,净光合速率增长趋于平缓。
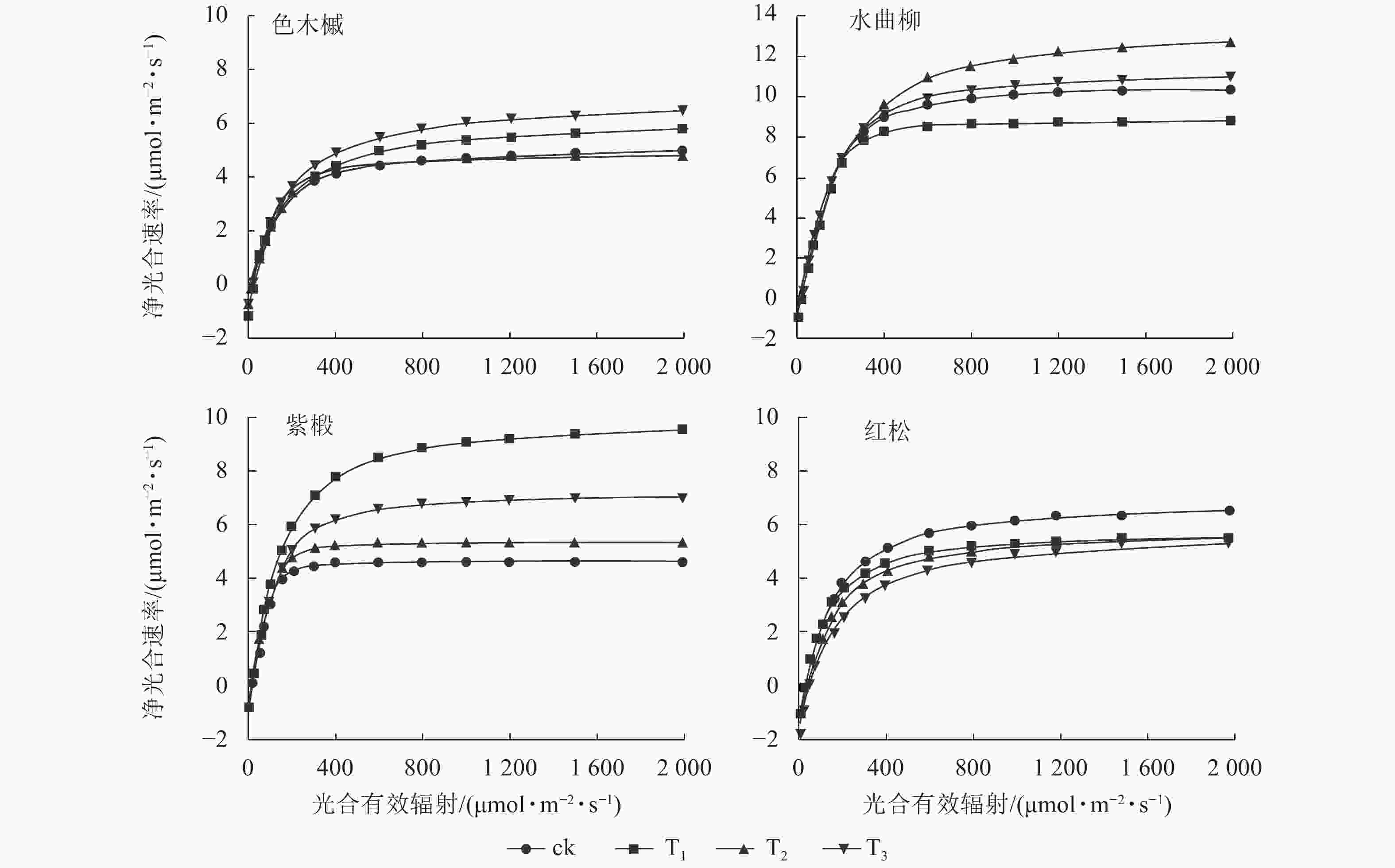
Figure 1. Net photosynthetic rates of the 4 major tree species at different logging intensities
通过非直角双曲线模型得到色木槭、水曲柳、紫椴、红松4个树种的光响应参数,结果如表3所示。从种间比较来看,在所有处理中水曲柳最大净光合速率与表观量子效率均显著大于其他树种(P<0.05),仅在T1处理中与紫椴差异不显著。在ck与T1处理中各树种间暗呼吸速率与光饱和点无显著差异;T2与T3处理中,红松的光饱和点显著高于其他3种阔叶树(P<0.05)。ck与T1处理中,各树种间光补偿点无显著差异;在T2处理中,红松光补偿点显著大于3种阔叶树(P<0.05);在T3组中红松和色木槭光补偿点显著大于水曲柳与紫椴(P<0.05)。
树种 采伐强度 暗呼吸速率/
(μmol·m−2·s−1)最大净光合速率/
(μmol·m−2·s−1)光饱和点/
(μmol·m−2·s−1)光补偿点/
(μmol·m−2·s−1)表观量子效率/
(μmol·m−2·s−1)色木槭 ck 0.68±0.22 Aa 4.94±0.28 Cab 1 538.21±661.82 Aa 20.27±22.30 Aa 0.02±0.01 Ca T1 1.19±0.14 Aa 6.17±1.09 Bab 1 341.36±103.88 Aa 22.28±1.94 Aa 0.02±0.00 Ba T2 0.78±0.42 Ba 4.87±0.68 Bb 1 719.69± 1150.68 Ba12.91±6.29 Ca 0.02±0.01 Ba T3 1.26±0.90 ABa 7.11±0.18 Ba 1 773.22±333.35 Ba 20.98±19.98 Aa 0.03±0.00 Ba 水曲柳 ck 1.51±0.53 Aa 10.57±1.40 Aab 1 335.97±473.77 Aab 21.73±10.80 Aa 0.04±0.01 Aa T1 1.35±0.35 Aa 9.17±0.67 Ab 928.98±85.38 Ab 17.12±4.20 Aa 0.04±0.00 Aa T2 1.37±0.20 ABa 12.26±0.72 Aa 1 505.02±199.00 BCa 20.83±3.18 Ba 0.04±0.00 Aa T3 0.99±0.26 Ba 10.96±0.62 Aa 1 253.41±113.48 Bab 13.30±3.51 Ba 0.04±0.00 Aa 紫椴 ck 1.08±0.16 Aa 4.95±0.57 Cc 699.23±36.82 Ab 14.18±3.48 Aa 0.03±0.00 Ba T1 0.81±0.10 Aa 9.52±0.25 Aa 1 859.17±703.38 Aa 11.56±0.42 Aa 0.03±0.00 Aa T2 1.10±0.28 ABa 5.76±1.15 Bc 780.74±168.10 Cb 15.16±4.26 BCa 0.03±0.01 ABa T3 1.15±0.37 Ba 7.28±0.51 Bb 1 084.89±370.58 Bb 15.30±1.98 Ba 0.03±0.01 Ba 红松 ck 1.24±1.07 Aa 6.61±0.66 Ba 2 953.14±971.84 Aa 25.80±23.92 Ab 0.03±0.00 Ba T1 1.15±0.38 Aa 5.65±0.70 Bb 2 132.14± 1377.96 Ab23.82±12.63 Ab 0.02±0.01 Ba T2 1.46±0.39 Aa 5.60±0.63 Bb 2 525.07± 1532.40 Aa35.53±9.64 Aab 0.02±0.01 Ba T3 2.28±0.99 Aa 5.89±0.32 Cab 2 858.82± 1587.13 Aa53.95±10.09 Aa 0.02±0.01 Ca 说明:数据为平均值±标准差。不同小写字母表示不同采伐强度间差异显著(P<0.05),不同大写字母表示不同树种间差异显著(P<0.05)。 Table 3. Light response parameters of 4 tree species under different logging intensities
从不同处理间的比较来看,各树种暗呼吸速率和表观量子效率在各处理间差异不显著。色木槭的光补偿点、光饱和点各处理间差异不显著, T3处理的最大净光合速率显著大于T2处理(P<0.05)。水曲柳的光补偿点在各处理间差异不显著, T2、T3处理的最大净光合速率显著大于T1处理,T2处理的光饱和点显著大于T1处理(P<0.05)。紫椴的光补偿点在各处理理间差异不显著, T1处理的最大净光合速率和光饱和点显著大于其他处理(P<0.05)。ck处理红松的最大净光合速率显著大于其他处理(P<0.05), T1处理光饱和点显著低于其他处理(P<0.05),T3处理光补偿点显著大于ck与T1处理(P<0.05)。
-
由表4可见:红松的比叶重、叶干物质质量分数与叶组织密度均显著高于其他3种阔叶树种(P<0.05),且红松的3种叶结构性状在各采伐强度下种内差异均不显著,展现出针叶叶片结构上的稳定性。从阔叶树种的种间对比来看,仅有T3处理下色木槭比叶重显著大于水曲柳(P<0.05),其他组内无种间差异。各阔叶树种叶干物质质量分数的变化规律一致,从大到小均为色木槭、紫椴、水曲柳。水曲柳的叶组织密度显著低于其他树种(P<0.05)。
树种 采伐强度 比叶重/(g·cm−2) 叶干物质质量分数/% 叶组织密度/(g·cm−3) 叶绿素相对含量 色木槭 ck 23.31±2.18 Bb 0.35±0.03 Bab 0.24±0.02 ABb 40.33±3.23 Aa T1 34.11±6.52 Ba 0.33±0.01 Bb 0.27±0.07 BCab 43.42±7.54 Aa T2 28.53±3.19 Bab 0.38±0.01 Aa 0.31±0.02 ABa 40.94±3.17 Aa T3 32.18±5.94 Ba 0.38±0.03 Aa 0.34±0.05 Aa 41.85±3.43 Aa 水曲柳 ck 20.95±2.48 Ba 0.22±0.01 Cb 0.17±0.02 Cb 34.98±2.68 Ba T1 29.24±4.86 Ba 0.25±0.01 Da 0.20±0.02 Ca 37.94±4.54 Aa T2 22.26±6.00 Ba 0.23±0.03 Bab 0.18±0.03 Cab 37.08±4.36 ABa T3 23.56±3.67 Ca 0.24±0.01 Bab 0.18±0.02 Bab 36.51±1.90 Ba 紫椴 ck 18.21±1.73 Bc 0.32±0.04 Bab 0.18±0.02 BCc 34.85±3.96 Ba T1 31.71±5.84 Ba 0.30±0.03 Cb 0.29±0.03 ABa 36.70±4.83 Aa T2 23.13±3.22 Bbc 0.34±0.04 Aab 0.25±0.03 BCb 34.16±2.72 Ba T3 27.45±3.81 BCab 0.35±0.04 Aa 0.25±0.02 Bb 38.26±2.46 ABa 红松 ck 122.04±2.23 Aa 0.39±0.01 Aa 0.29±0.08 Aa - T1 126.10±1.93 Aa 0.40±0.02 Aa 0.35±0.04 Aa - T2 125.42±6.30 Aa 0.39±0.04 Aa 0.39±0.14 Aa - T3 124.63±3.37 Aa 0.39±0.02 Aa 0.36±0.08 Aa - 说明:数据为平均值±标准差。不同小写字母表示不同采伐强度间差异显著(P<0.05),不同大写字母表示不同树种间差异显著(P<0.05)。-表示无此项。 Table 4. Leaf structural traits and SPAD of 4 tree species under different logging intensities
从不同处理间比较来看,3种阔叶树种的比叶重呈相同的变化规律,从大到小均为T1、T3、T2、ck。色木槭与紫椴的叶干物质质量分数最大值出现在T3处理,且显著大于T1处理(P<0.05)。水曲柳的叶干物质质量分数与之相反,最大值出现在T1处理。色木槭的叶组织密度随采伐强度的增加而增加,在T3处理显著大于ck处理(P<0.05),水曲柳与紫椴的叶组织密度最大值出现在T1处理,且均显著大于ck处理。
ck处理的色木槭叶绿素相对含量显著大于紫椴和水曲柳(P<0.05);T2处理的色木槭叶绿素相对含量显著大于紫椴(P<0.05);T3处理中色木槭叶绿素相对含量显著大于水曲柳(P<0.05)。各树种的叶绿素相对含量在不同采伐强度下种内差异均不显著。
-
采伐是一种重要的森林经营措施,合理强度的采伐能够减弱竞争、改善光照条件和地下水肥条件,从宏观上促进森林生产力的提升[21]。本研究发现:从光合生理方面来说,采伐对4种树种的光合特性均产生了显著影响,且各树种对不同采伐强度的响应模式不同,体现出不同树种面对环境变化时光合策略的差异。
色木槭的光合能力(最大净光合速率和净光合速率)在重度采伐条件下显著高于其他处理,而其他光合参数均不随采伐强度产生显著变化,表明内源性因素并非驱动其光合速率提升的主要原因。采伐会显著提高林下光照水平。陈涵等[22]研究发现:随着采伐强度的增大,冠层开度、直射光和散射光均有显著增加,因此重度采伐可能使林下光照接近或达到色木槭的光饱和点,从而提升了其光合速率。黄一鑫等[19]研究也发现:在光照强度大的林窗环境下,色木槭最大净光合速率显著高于光强较弱的林下和林缘,为本研究推断提供了进一步佐证。色木槭在中度采伐条件下净光合速率显著低于其他采伐强度,胞间CO2摩尔分数和蒸腾速率最低,水分利用效率最高,证明气孔限制了叶片与外界的气体交换和蒸腾作用,植物通过提升水分利用效率来适应环境,保障光合作用的产出[23]。
水曲柳的净光合速率、气孔导度、蒸腾速率、最大净光合速率及表观量子效率在各个采伐强度下均显著高于其他树种,表现出极强的光合能力和对环境的适应能力。水曲柳的光合速率和气孔开闭随环境变化改变可塑性极强,且两者的响应存在显著耦合关系,这增强了水曲柳在面临不同强度采伐时对环境的适应性[24]。孔鑫等[25]研究也发现:水曲柳幼苗的光合和水力性状对光照强度的可塑性较强,使得水曲柳在森林异质性较高的光环境中的生存能力显著提升,从侧面印证了本研究的观点。
紫椴的最大净光合速率与水分利用效率均是在未采伐状态下最低,轻度采伐时最高。在本研究采伐样地的另一项研究表明:未采伐林分中冠层郁闭导致光合有效辐射低于紫椴最适区间,可能引发其光限制效应[26]。此外,王宣璎等[27]研究还发现:紫椴对光适应范围极为狭窄,因此导致紫椴的光合能力在强度采伐时上升后又在中度与重度采伐时产生显著的光抑制。
红松的光合特性对采伐强度的响应呈现出复杂的生态适应机制。随着采伐强度增加,红松的净光合速率显著下降,对照组较高的最大净光合速率表明,完整的林冠结构通过适度遮荫维持了稳定的光温条件,避免了强光抑制和水分过度蒸散,从而优化了红松的光合碳同化能力[28]。重度采伐组的光饱和点显著高于对照组和轻度采伐组,这可能是红松面对强光胁迫的适应性调整,通过提升光能利用上限来补偿因林分结构破坏导致的散射光减少[29]。然而,这种高光饱和点并未转化为实际光合效率的提升,暗示重度干扰可能引发光系统损伤或代谢失衡[30]。胞间CO2摩尔分数在各处理间的差异则揭示了非气孔限制的存在:尽管气孔导度未显著变化,但重度采线的CO2摩尔分数显著低于轻度采伐组,结合水分利用效率的逆向响应,表明叶肉导度降低可能是主要内源性限制因素[31]。蒸腾速率在轻度采伐时下降则可能源于被动的气孔调节,这种水分保存策略虽提升水分利用效率却牺牲了光合速率[32]。这些结果共同印证了适度采伐的生态合理性,既能通过林隙形成促进光能利用,又可避免高强度的采伐引发红松适应不良[33]。
-
植物通过选择不同的响应策略有针对性地改变叶片结构,从而对环境表现出不同的适应性[7]。本研究结果显示:采伐改变了3种阔叶树种的各项叶结构性状。阔叶乔木的比叶重反映其资源利用效率与光合能力,高比叶重体现为叶片结构紧密、光合组织密集,适应干旱或强光环境从而提升资源利用效率[34]。本研究中,3种阔叶树种的比叶重均在轻度采伐时提升最为显著,反映植物对资源分配的优化策略:轻度干扰下光资源增加而胁迫较弱,叶片通过适度增厚提高光能捕获效率[4];中重度采伐下光照强度虽持续增加,但可能因机械损伤或水分胁迫抑制比叶面积的进一步提升[35]。此外,对照组因冠层郁闭度高、光限制显著,叶片倾向于降低构建成本,符合叶片经济谱理论[36]。从树种角度来看,色木槭的比叶重在重度采伐条件下显著高于水曲柳与紫椴。这可能与色木槭具有更强的光适应能力有关:色木槭在干扰后通过增加叶片厚度及调整结构比例来提升抗逆性[37],而水曲柳和紫椴因生态策略差异未形成类似响应。
叶干物质质量分数表征碳投资策略,高叶干物质质量分数指示叶片抗逆性强、代谢缓慢,通过增加木质素等结构物质延长寿命,但牺牲生长速率[38]。在各采伐强度下,3种阔叶树种的叶干物质质量分数从大到小均为色木槭、紫椴、水曲柳,体现出一定的稳定性,这可能是由于色木槭的策略倾向于保守,通过更高的叶干物质增强养分保留能力,从而适应资源贫瘠或扰动强烈的竞争环境[39];水曲柳作为开拓性树种,低的叶干物质质量分数表明其倾向于快速生长来提升在竞争环境中的适应能力[40]。各树种对采伐的响应也具有差异:在轻度采伐下,水曲柳叶干物质达到峰值,这可能与水曲柳对光照变化较为敏感有关[28]。作为喜光树种,在轻度采伐下水曲柳所处的光照环境得到改善,叶片能够吸收更多的光能进行光合作用,合成更多有机物并储存在叶片中,进而使得叶干物质显著增加[30]。紫椴在轻度采伐下叶干物质质量分数最低。前人研究表明:叶干物质通常与光合速率呈负相关。本研究也发现:紫椴光合速率与水分利用效率在轻度采伐下达到最大,与叶干物质质量量分数呈现负相关,与前人观点一致[41]。在重度采伐下,色木槭与紫椴叶干物质质量分数达最大值,可能源于高强度干扰加剧资源竞争和土壤养分限制,高叶干物质质量分数通过提升组织密度增强对物理损伤的抗性,同时延长叶片寿命以适应资源胁迫[42]。同时,色木槭的光饱和点也在重度采伐强度下达到最大,代表着色木槭通过增加叶组织的机械强度更有效地拦截和利用强光,从而提高对高强度环境胁迫的耐受性[28]。
叶组织密度关联叶片机械强度与光合效率,高叶组织密度通过增厚栅栏组织或细胞密度增加光能捕获率,但可能限制气体交换,体现生长与抗胁迫间的平衡[43]。水曲柳的叶组织密度总是显著低于紫椴和色木槭,与其比叶重、叶干物质质量分数的变化规律保持一致,共同提示水曲柳倾向于快速光合而非增强机械防御的生长策略[44]。紫椴的叶组织密度在轻度采伐时达到峰值,可能作为耐荫树种,适度干扰既缓解竞争又不致暴露于过强光照,使紫椴在光能获取与叶结构稳定间达到最优平衡[17]。徐程扬[45]研究也发现:紫椴幼苗能在环境光强增加时通过改变冠层结构提升其光截获能力,从冠层结构的角度补充支持了本研究结果。比叶重、叶干物质质量分数与叶组织密度三者共同反映了植物在光合产物分配、环境适应及资源获取-保存策略间的平衡[46]。
叶绿素相对含量的增加是植物为了提高光能捕获效率而做出的适应性调整[47]。不同的采伐强度对各树种的叶绿素相对含量的影响不大,这可能是由于叶绿素作为光合作用的关键色素,其相对含量受到多种因素的共同调控。在采伐过程中,虽然光照、温度、水分等环境因子会发生变化,但这些变化可能并未达到足以显著影响叶绿素合成的阈值[48]。
-
本研究检验了不同采伐强度对吉林蛟河4种主要树种叶光合特性、叶结构性状以及叶绿素相对含量的影响。主要结论如下:①采伐改善了3种阔叶树种的光合能力,色木槭、水曲柳、紫椴分别在重度、中度、轻度采伐强度下碳同化能力最强,采伐普遍提升了3种阔叶树种的比叶重、叶干物质质量分数和叶组织密度。②红松的光合能力随采伐强度增加而下降,叶结构性状不随采伐强度增加而变化,体现出红松针叶适应环境变化的保守策略。③紫椴与水曲柳通过提升光饱和点来适应中低强度的采伐,体现出这2个树种对于强光的捕获潜能。④采伐强度对各树种叶绿素相对含量、暗呼吸速率影响不显著。总体而言,中低强度的采伐可促进针阔混交林中阔叶树种的光合能力及资源配置效率,但高强度的采伐会造成红松适应不良。
Impact of logging intensity on leaf functional traits of conifer-broadleaf mixed forest in Jiaohe, Jilin Province
doi: 10.11833/j.issn.2095-0756.20250155
- Received Date: 2025-02-24
- Accepted Date: 2025-06-30
- Rev Recd Date: 2025-06-22
- Available Online: 2026-01-27
- Publish Date: 2026-02-20
-
Key words:
- logging intensity /
- conifer-broadleaf mixed forest /
- leaf functional traits /
- photosynthetic properties /
- leaf structural traits
Abstract:
| Citation: | YANG Zhiyi, FENG Xinyi, SHU Ting, et al. Impact of logging intensity on leaf functional traits of conifer-broadleaf mixed forest in Jiaohe, Jilin Province[J]. Journal of Zhejiang A&F University, 2026, 43(1): 33−44 doi: 10.11833/j.issn.2095-0756.20250155

|
 DownLoad:
DownLoad: