




-
营养元素是植物生长的基础,不同器官营养元素的含量和积累体现了植物对其的需求量和吸收能力,植物营养元素的积累与分布是研究森林生态系统物流和能流的基础,直接影响森林生产力,一定程度上制约着地力的演变,对维持林地的养分平衡有重要作用[1]。竹子是集经济、生态和社会效益于一体的优良林种,是区域社会经济发展和生态环境保护的重要刚性资源,被誉为21世纪最有发展前景的植物类型[2]。了解营养元素在竹子各器官中的分布、积累及分配规律对于指导竹林生产经营、提高系统的养分利用率和最大限度地提高林地生产力和可持续经营都具有重要意义。黄甜竹Acidosasa edulis隶属于禾本科Gramineae竹亚科Bambusoideae酸竹属Acidosasa,是一种优质笋用竹种。黄甜竹笋期为4−6月,竹笋产量高、品质佳,是已知竹笋中营养成分最丰富最合理的一种[2]。但目前对黄甜竹的研究主要集中于栽培技术、生长发育规律、竹笋营养成分和次生代谢产物[3-6]等方面,关于竹类植物营养元素的研究主要集中在毛竹Phyllostachys edulis[7-10]上,其他竹种如雷竹Phyllostachys violascens[11]、苦竹Pleioblastus amarus[12]、绿竹Bambusa oldhamii[13]、黄竹Dendrocalamus membranaceus[14]、青皮竹Bambusa textilis[15]等也有涉及,但黄甜竹的相关报道尚未见。本研究通过分析黄甜竹地上器官中氮(N)、磷(P)、钾(K)、钙(Ca)、镁(Mg)、铁(Fe)、锰(Mn)、铜(Cu)和锌(Zn)等9种营养元素,了解黄甜竹不同年龄和不同器官的营养元素的变化规律,为黄甜竹资源开发利用提供科学基础。
-
试验地位于浙江省丽水市莲都区丽水市林业科学研究院百果园科研基地(28°28′ N,119°53′ E),平均海拔为150 m,平均坡度为25°。该区属浙西南低山丘陵地带,年平均气温为18.1 ℃,年平均日照时数为l 783 h,年降水量1 427 mm,无霜期为256 d,极端最高气温为43.2 ℃,极端最低气温为−7.5 ℃。土壤为红壤,pH 5.1,有机质为29.8 g·kg−1,碱解氮、有效磷和速效钾分别为134.1、3.2和124.3 mg·kg−1。试验地于1990年建园,长期人工经营,黄甜竹林面积20 hm2,林分长势良好,立竹密度10 250株·hm−2,平均胸径为5.2 cm,林分保留3年生及以下的立竹,竹林结构1年生∶2年生∶3年生=1∶1∶1,4年生及以上竹子全部采伐。每年夏进行竹林垦复并施复合肥[m(氮)∶m(磷)∶m(钾)=18∶5∶8] 1次,施肥量1 500 kg·hm−2。
-
2018年10月,在人工种植的黄甜竹纯林中,设立20 m×20 m标准样地3个,立竹密度(10 250±267)株·hm−2。在标准地内每株检尺,按不同竹龄测量并计算黄甜竹平均胸径;各样地中分别选取1年生、2年生、3年生平均胸径的标准竹各4株,采用全收获法砍伐。
-
参照WU等[16]方法进行样品采集和生物量调查,按样地中各年龄标准株生物量和各年龄株数[15]计算地上部生物量。叶、枝、秆样品分别置于样品袋中,带回实验室分析;分别用去离子水清洗后,在105 ℃下杀青30 min,然后在80 ℃烘干至恒量,粉碎后测定元素质量分数。氮(N)的测定:浓硫酸-双氧水(H2SO4-H2O2)消化后采用凯氏半微量法(全自动凯氏定氮仪,FOSS)测定。磷(P)、钾(K)、钙(Ca)、镁(Mg)等的测定:采用微波消解法消解样品,用IRIS/AP型全谱直读电感耦合等离子体发射光谱仪测定(ICP-OES法)[17]。计算营养元素积累量(吸收量)=营养元素质量分数×干物质量;植物地上部某器官营养元素分配=某器官营养元素积累量/地上部积累量×100%[13]。
-
使用Excel 2003和SPSS 22分析软件进行数据处理。
-
由表1可知:不同竹龄黄甜竹地上部单株生物量均以秆最高(4 107.78 g),叶最低(661.11 g)。单株叶、枝、秆的生物量分别占地上部生物量的11.56%、16.56%和71.86%,说明黄甜竹生物量主要集中在秆。随竹龄增大,不同部位生物量均逐渐增大,其中3年生黄甜竹秆、枝、叶生物量比1年生分别提高17.24%、38.43%和137.40%;随竹龄增长,叶和枝生物量占地上部生物量的比例有所提升。
表 1 黄甜竹标准株生物量
Table 1. Biomass of A. edulis standard plant
竹龄 秆/(g·株−1) 枝/(g·株−1) 叶/(g·株−1) 合计/(g·株−1) 1年生 3 866.67±359.49 bA 806.67±70.24 bB 410.00±26.46 cC 5 086.67 2年生 3 923.33±101.16 bA 916.67±40.41 abB 600.00±45.83 bC 5 440.00 3年生 4 533.33±446.58 aA 1 116.67±140.12 aB 973.33±45.09 aB 6 623.33 平均 4 107.78 946.67 661.11 5 716.67 说明:小写字母表示不同竹龄间差异显著(P<0.05),大写字母表示不同部位间差异显著(P<0.05) -
植物不同器官生理机能不同,不同营养元素在植物体内的功能也不同,因此营养元素在植物不同部位及不同营养元素在同一部位中的分布具有差异性[18-19]。由表2可知:黄甜竹地上部营养元素最高为叶(48.73 g·kg−1),其次是枝(17.61 g·kg−1),秆中最低(16.30 g·kg−1),与毛竹[8]、雷竹[11]、青皮竹[15]、高节竹Phyllostachys prominens[20]、慈竹Neosinocalamus affinis[21]等的研究结果一致。对不同部位营养元素分布的方差分析可知:除Zn外,叶中其他8种营养元素质量分数均显著高于秆和枝(P<0.05)。N、P和K质量分数分别为31.20、1.18和7.49 g·kg−1,分别比枝高133.13%、178.27%和172.03%,比秆高215.79%、81.59%和62.80%;Ca比枝和秆分别高971.70%和952.40%,Mg比枝和秆分别高出856.62%和470.50%;Fe、Mn和Cu质量分数分别比枝高208.29%、192.26%和116.53%,比秆高132.50%、247.14%和197.07%。叶和秆中Zn质量分数分别为37.27和37.55 mg·kg−1,两者差异不显著(P>0.05),但均显著高于枝中(P<0.05)。枝和秆中营养元素质量分数高低不一,N质量分数表现为枝中高于秆中(P<0.05),P、K和Zn表现为秆中高于枝中(P<0.05),Ca、Mg、Fe、Mn和Cu在枝和秆中差异不显著(P>0.05)。
表 2 不同竹龄黄甜竹各部位营养元素质量分数分析
Table 2. Nutrient content in different parts of A. edulis with different bamboo ages
部位 竹龄 N/(g·kg−1) P/(g·kg−1) K/(g·kg−1) Ca/(g·kg−1) Mg/(g·kg−1) 叶 1年生 31.45±2.76 aA 1.14±0.03 aA 7.81±0.39 aA 5.40±1.01 bA 1.65±0.06 aA 2年生 30.20±3.57 aA 1.21±0.08 aA 7.33±0.24 aA 6.26±0.91 abA 1.39±0.23 aA 3年生 31.94±1.77 aA 1.18±0.27 aA 7.34±0.80 aA 8.00±1.42 aA 1.51±0.32 aA 平均 31.20±1.53 A 1.18±0.07 A 7.49±0.06 A 6.55±0.53 A 1.52±0.17 A 枝 1年生 15.88±1.48 aB 0.58±0.08 aB 4.42±0.24 aB 0.67±0.18 aB 0.26±0.01 aB 2年生 13.70±0.32 bB 0.47±0.13 aC 2.37±0.17 bC 0.49±0.10 aB 0.10±0.03 bB 3年生 10.56±1.09 cB 0.22±0.04 bC 1.47±0.07 cC 0.68±0.15 aB 0.11±0.05 bB 平均 13.38±0.52 B 0.42±0.06 C 2.75±0.13 C 0.61±0.07 B 0.16±0.02 B 秆 1年生 9.52±1.79 aC 0.54±0.11 aB 6.30±0.53 aA 0.26±0.02 bB 0.18±0.01 bB 2年生 9.33±1.21 aC 0.79±0.28 aB 4.03±0.32 bB 0.47±0.10 bB 0.21±0.04 abB 3年生 10.79±0.84 aC 0.61±0.12 aB 3.48±0.37 bB 1.13±0.52 aB 0.40±0.16 aB 平均 9.88±1.17 C 0.65±0.17 B 4.60±0.18 B 0.62±0.15 B 0.27±0.06 B 部位 竹龄 Fe/(mg·kg−1) Mn/(mg·kg−1) Cu/(mg·kg−1) Zn/(mg·kg−1) 叶 1年生 282.01±24.01 aA 551.68±53.53 aA 7.22±0.36 aA 38.65±7.87 aA 2年生 177.98±30.25 bA 519.09±53.53 aA 6.43±0.33 bA 35.16±6.50 aA 3年生 152.25±10.06 bA 567.65±34.83 aA 5.72±0.18 bA 38.00±8.42 aB 平均 204.08±21.44 A 546.14±47.30 A 6.46±0.29 A 37.27±7.6 A 枝 1年生 39.45±10.03 bC 165.74±50.11 aB 3.98±1.13 aB 25.31±2.75 bB 2年生 38.02±6.89 bC 166.21±89.96 aB 2.86±0.32 abB 23.07±4.33 bB 3年生 121.13±38.06 aA 228.66±111.23 aB 2.10±0.08 bB 36.88±3.33 aB 平均 66.20±18.33B 186.87±83.76B 2.98±0.51 B 28.42±3.47 B 秆 1年生 88.13±20.34 aB 46.39±12.99 bC 2.37±0.15 aC 29.86±5.76 aB 2年生 95.86±19.35 aB 124.30±34.72 abB 2.36±0.38 aB 33.67±6.68 aA 3年生 79.34±6.61 aB 301.28±79.33 aB 1.79±0.26 aB 49.11±7.58 aA 平均 87.78±15.43 B 157.32±42.34 B 2.17±0.27 B 37.55±6.67 A 说明:小写字母表示不同竹龄间差异显著(P<0.05),大写字母表示不同部位间差异显著(P<0.05) 同一部位中不同营养元素质量分数也不同,叶中各元素质量分数从高到低依次为N、K、Ca、Mg、P、Mn、Fe、Zn、Cu,枝中依次为N、K、Ca、P、Mn、Mg、Fe、Zn和Cu,秆中依次为N、K、P、Ca、Mg、P、Mn、Fe、Zn、Cu。不同部位中各营养元素质量分数大致相似,最大的2种元素均为N和K,最小的2种元素均为Cu和Zn,与葛高波等[14]在黄竹上的研究结果相似。
竹龄对不同部位营养元素质量分数影响不同。总体上看,叶受竹龄影响最小,秆其次,枝受竹龄影响最大。具体看,叶中N、P和K质量分数受竹龄影响不大,枝中N、P、K质量分数随竹龄增大而降低,秆中N、P质量分数受竹龄影响不大,但K质量分数随竹龄增大而降低,其中1年生显著高于2年生、3年生立竹(P<0.05)。叶和秆中Ca质量分数随竹龄增大而增大,叶中Mg质量分数受竹龄影响不大,枝中表现为1年生最高,显著高于2年生和3年生(P<0.05),秆中则表现为逐年增大。叶中Fe和Cu表现为随竹龄增大而降低,Mn和Zn较稳定,受竹龄影响不大;枝中Fe和Zn表现为3年生高于1年生和2年生,Cu表现为逐年下降;秆中Mn质量分数表现逐年增高,各竹龄差异显著(P<0.05),Zn随竹龄增大而逐年增高,但差异不显著(P>0.05),而Fe和Cu受竹龄影响不显著(P>0.05)。
-
从表3可知:黄甜竹地上部9种营养元素积累总量为1 186.69 kg·hm−2,秆中的积累量最大(687.38 kg·hm−2),其次是叶(333.01 kg·hm−2),枝积累量最小,仅166.29 kg·hm−2,与绿竹[13]研究结果相同。不同营养元素累积量从大到小依次为N、K、Ca、P、Mg、Mn、Fe、Zn、Cu。
表 3 黄甜竹不同部位各营养元素的积累量
Table 3. Accumulation of nutrient elements in different parts of A. edulis
部位 N/
(kg·hm−2)P/
(kg·hm−2)K/
(kg·hm−2)Ca/
(kg·hm−2)Mg/
(kg·hm−2)Cu/
(kg·hm−2)Zn/
(kg·hm−2)Fe/
(kg·hm−2)Mn/
(kg·hm−2)合计/
(kg·hm−2)叶 212.18 8.00 50.37 47.00 10.19 0.04 0.25 1.27 3.72 333.01 枝 126.98 3.90 25.22 5.87 1.47 0.03 0.28 0.69 1.85 166.29 秆 417.91 27.21 191.10 27.34 11.50 0.09 1.61 3.68 6.95 687.38 合计 757.06 39.11 266.69 80.21 23.16 0.16 2.14 5.63 12.52 1 186.69 如图1可知:9种营养元素在黄甜竹叶、枝、秆中的分配率不同,除Ca外,其他元素的分配率均表现为秆中最大,其次是叶,枝中最小;秆中分配率最低的为Ca(34.09%)和Mg(49.65%),另外7种元素均大于50%。叶中分配率最高的为Ca和Mg,占营养元素总量的58.59%和43.99%。
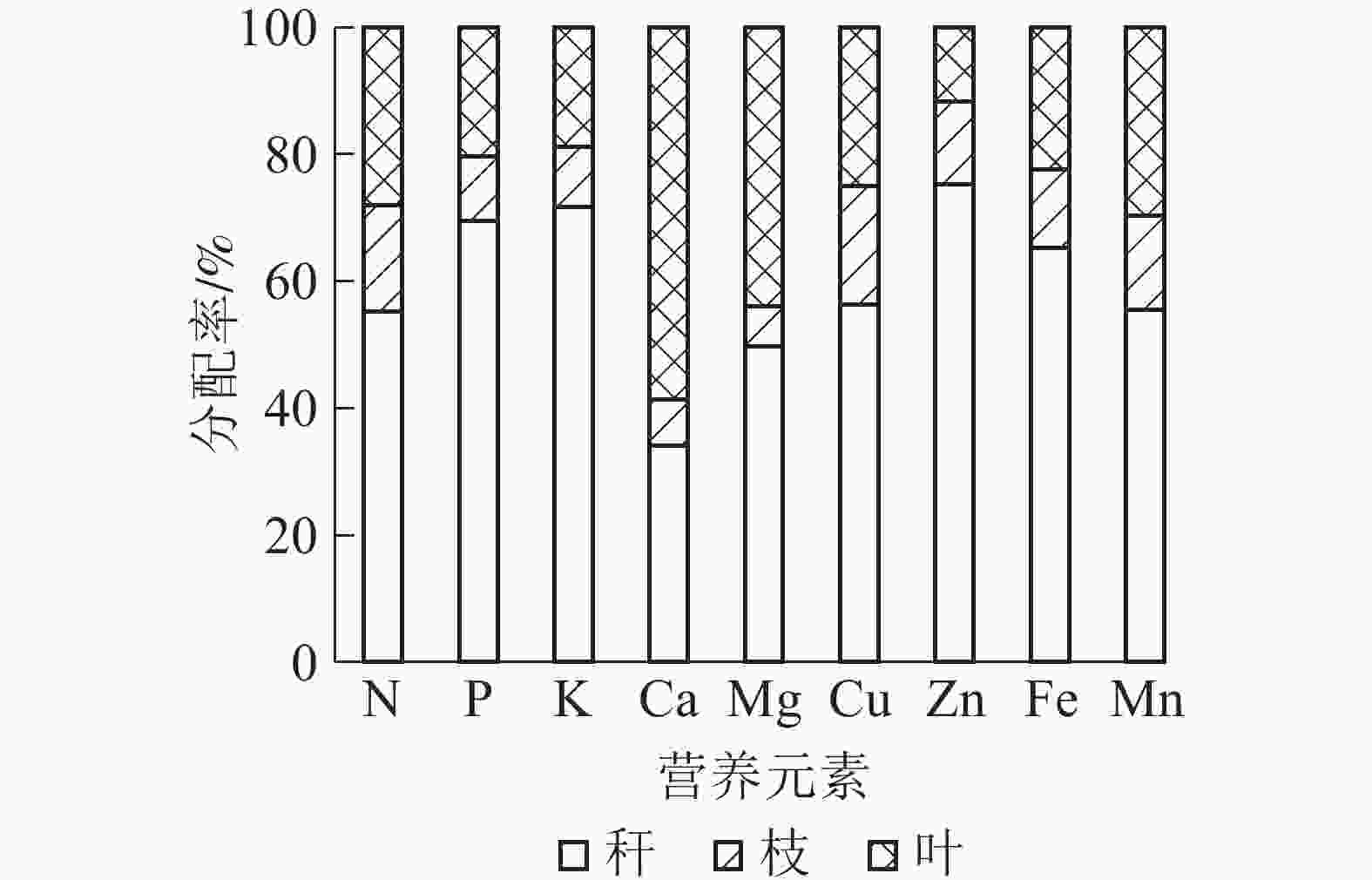
图 1 黄甜竹营养元素在各部位中的分配
Figure 1. Distribution of nutrient elements in aboveground parts of A. edulis
-
植物营养元素含量是植物生物学特性与生态环境相统一的结果,不但反映了植物自身的特征,也反映了植物在一定生境下从土壤中吸收和蓄积养分的能力[22]。竹子依靠根鞭吸收养分,按各器官所需运输、分配并积累;长期的自然进化导致竹叶作为有机物合成的“加工厂”,代谢活动最为活跃,营养元素需求量也最多,这也是黄甜竹竹叶中大部分营养元素远高于枝和秆的原因,与秦武明等[23]、刘鹏等[24]认为代谢越活跃的部位营养元素含量越高的观点一致。从营养元素功能的角度看,氮、磷、镁等是光合色素的重要组成元素,钾参与光合酶活化、促进碳水化合物合成、加速光合产物的流动,是确保光合过程高效运行的重要因子,因此,N、P、K、Mg等元素在光合器官叶中的质量分数显著高于非光合器官枝和秆。各营养元素在地上部分布规律大体相似,不同部位质量分数最大的2种元素均为N和K,最小的均为Cu和Zn,可能与物种的遗传特性密切相关[25]。对不同经营方式下毛竹各器官营养元素的研究表明[9],毛竹各器官营养元素在集约经营和粗放经营中差异并不显著,表现出了较高的遗传稳定性。
黄甜竹不同器官通过有机配合进行物质的循环和能量的流动,以适应生长发育需要。随着竹龄的增长,竹叶通过周期性换叶增加竹叶数量实现叶生物量的积累,而竹枝和竹秆主要是靠竹壁增厚和竹壁密度的增加来实现生物量的积累,因此不同部位营养元素质量分数受竹龄的影响不同。本研究发现:不同竹龄的竹叶中营养元素质量分数差别不大,而秆和枝则受年龄的影响较大;说明不同竹龄竹叶生理状态和有机物合成能力相似,尽管竹龄增加了,但由于周期性脱落换新,叶龄相近。不同器官对养分需求量有差异,不同竹龄黄甜竹生物量增长不同,1年生单株枝和秆的生物量增长远高于2年生和3年生,与黄张婷等[11]和杨杰等[26]的研究结果相似,说明竹类植物在不同生长阶段竹叶对养分的合成能力差异不大,但在物质分配和积累上有较大差异。
黄甜竹地上部营养元素主要积累在秆中,其次为叶,枝中最少,积累量高低与生物量大小不完全一致,差异主要体现在叶和枝上,原因在于生物量的构成不仅仅包括上述9种营养元素,还包括大量的碳(C)、氢(H)、氧(O)等其他元素;叶中营养元素含量远远高于枝,因此叶中营养元素的积累量高于枝。就各营养元素积累量来看,黄甜竹N积累量高达63.80%,表现出了极强的N吸收能力;以此计算林地需肥量,适时补充N肥,为黄甜竹林地养分管理和定向培育提供数据支撑。
竹叶中含多种矿质元素,其中很多有一定的生理和药理作用,是竹叶保健功能的物质基础之一[27]。K在人体中可维持碳水化合物、蛋白质代谢,维持细胞正常渗透压;Ca参加骨骼和牙齿的组成;Fe参与血红蛋白形成,促进造血;Mn被认为是抗癌元素,是多种酶的催化剂;Zn参与多种酶的合成,促进人体生长发育,增强创伤组织再生[28]。本研究发现:黄甜竹叶中K、Ca、Fe、Mn、Zn等元素极为丰富,如K高达7.49 g·kg−1,Ca高达6.55 g·kg−1,是潜在的保健食品原料。在植物生长代谢过程中,营养元素不仅是细胞生长的物质来源,而且参与植物初生代谢和次生代谢,影响植物体内生理活性物质的组分和含量[29],在日益关注健康的当下,挖掘竹子的保健功能已成为竹产业发展的新业态与新趋势[29],分析营养元素的积累与分配可为黄甜竹竹叶资源综合开发利用提供理论基础和方向。
Content and distribution of the main nutrient elements in aboveground parts of Acidosasa edulis in southwest of Zhejiang Province
-
摘要:
目的 研究浙西南地区不同年龄黄甜竹Acidosasa edulis地上部营养元素质量分数及分配特性,为黄甜竹林地土壤养分管理和资源开发利用提供依据。 方法 采用全收获法计算生物量,测定不同年龄(1~3 a)和不同地上部位(叶、枝、秆)的植株样品中氮(N)、磷(P)、钾(K)、钙(Ca)、镁(Mg)、铁(Fe)、锰(Mn)、铜(Cu)和锌(Zn)等9种营养元素。 结果 ①黄甜竹地上部营养元素质量分数最高为叶(48.73 g·kg−1),其次为枝(17.61 g·kg−1),秆中最低(16.30 g·kg−1);叶中各营养元素从高到低依次为N、K、Ca、Mg、P、Mn、Fe、Zn、Cu,枝中为N、K、Ca、P、Mn、Mg、Fe、Zn、Cu,秆中为N、K、P、Ca、Mg、P、Mn、Fe、Zn、Cu。各部位营养元素质量分数受竹龄影响不同,其中枝受竹龄影响最大,秆次之,叶最小。②营养元素积累量秆中最大(687.38 kg·hm−2),其次是叶(333.01 kg·hm−2),枝中最小(166.29 kg·hm−2)。不同营养元素的积累量从大到小依次为N、K、Ca、P、Mg、Mn、Fe、Zn、Cu。③不同部位营养元素分配率不同,总体表现为秆中最高(57.92%),其次是叶(28.06%),枝中最低(14.01%)。 结论 黄甜竹中N元素积累量最高,表明黄甜竹有较强的氮吸收能力,生产上要适时补充氮肥。竹叶中营养元素全面,有较高的开发利用价值。图1表3参29 Abstract:Objective With an investigation of the content and distribution characteristics of the main nutrient elements in the aboveground parts of Acidosasa edulis of different ages located in the southwest of Zhejiang Province, this study is aimed to provide a scientific basis for the management and exploitation of soil nutrients, as well as the rational utilization of Phyllostachys pubescens resources. Method Total harvest method was employed to estimate the biomass, and the contents of nitrogen (N), phosphorus (P), potassium (K), calcium (Ca), magnesium (Mg), iron (Fe), manganese (Mn), copper (Cu) and zinc (Zn) in plant samples of different ages (1−3 a) in different parts (leaves, branches and culms). Result a) In the aboveground parts of A. edulis, the total contents of the nine nutrient elements were as follows: leaf (48.73 g·kg−1), branch (17.61 g·kg−1) and culm (16.30 g·kg−1); whereas the contents of nutrient elements in leaves, branches and culms were as follows: N, K, Ca, Mg, P, Mn, Fe, Zn and Cu in leaf; N, K, Ca, P, Mn, Mg, Fe, Zn and Cu in branch; N, K, P, Ca, Mg, P, Mn, Fe, Zn and Cu in culm. b) The contents of nutrient elements in different parts were affected differently by the bamboo age which means that branches were most affected, culms were the second, and leaves were the least affected. c) The accumulation of the nine kinds of nutrient elements in culms was the highest (687.38 kg·hm−2), followed by that of the leaves (333.01 kg·hm−2), and the branches (166.29 kg·hm−2). d) The accumulation of different nutrient elements in A. edulis is as follows: N, K, Ca, P, Mg, Mn, Fe, Zn and Cu. e) The distribution rate of nutrient elements was different in leaves, branches and culms with the total distribution rate as follows: culm (57.92%), leaf (28.06%) and branch (14.01%). Conclusion The accumulation of N is the highest, indicating that A. edulis has strong nitrogen absorption capacity. Thus, extra attention should be paid to the timely supplement of nitrogen fertilizer to ensure a better production. Also, the leaves of A. edulis have high utilization value because of their comprehensive nutrient elements for human health, thus highly exploitable. [Ch, 1 fig. 3 tab. 29 ref.] -
Key words:
- Acidosasa edulis /
- nutrient elements /
- accumulation /
- distribution
-
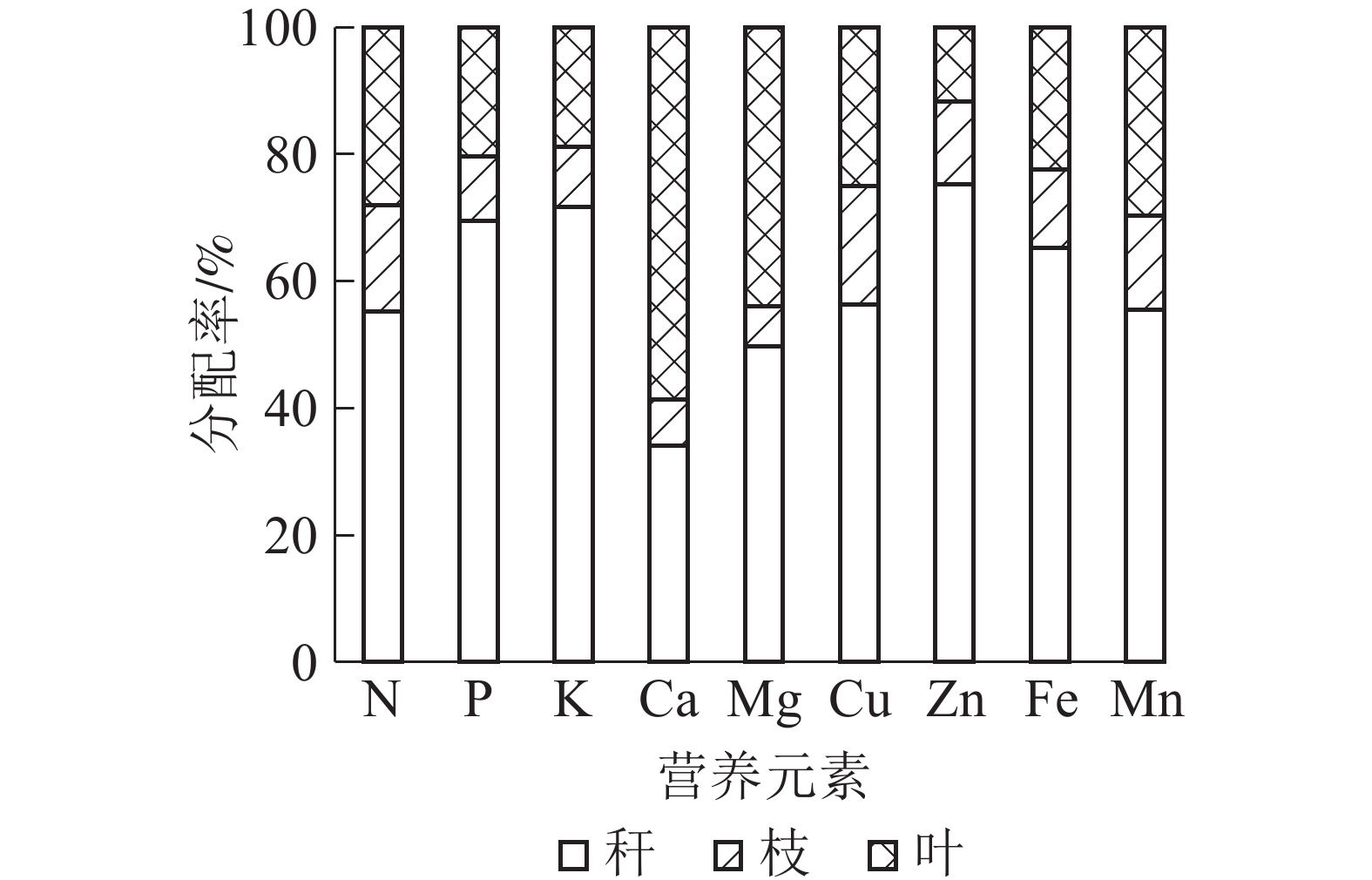
图 1 黄甜竹营养元素在各部位中的分配
Figure 1 Distribution of nutrient elements in aboveground parts of A. edulis
表 1 黄甜竹标准株生物量
Table 1. Biomass of A. edulis standard plant
竹龄 秆/(g·株−1) 枝/(g·株−1) 叶/(g·株−1) 合计/(g·株−1) 1年生 3 866.67±359.49 bA 806.67±70.24 bB 410.00±26.46 cC 5 086.67 2年生 3 923.33±101.16 bA 916.67±40.41 abB 600.00±45.83 bC 5 440.00 3年生 4 533.33±446.58 aA 1 116.67±140.12 aB 973.33±45.09 aB 6 623.33 平均 4 107.78 946.67 661.11 5 716.67 说明:小写字母表示不同竹龄间差异显著(P<0.05),大写字母表示不同部位间差异显著(P<0.05)  下载: 导出CSV
下载: 导出CSV
表 2 不同竹龄黄甜竹各部位营养元素质量分数分析
Table 2. Nutrient content in different parts of A. edulis with different bamboo ages
部位 竹龄 N/(g·kg−1) P/(g·kg−1) K/(g·kg−1) Ca/(g·kg−1) Mg/(g·kg−1) 叶 1年生 31.45±2.76 aA 1.14±0.03 aA 7.81±0.39 aA 5.40±1.01 bA 1.65±0.06 aA 2年生 30.20±3.57 aA 1.21±0.08 aA 7.33±0.24 aA 6.26±0.91 abA 1.39±0.23 aA 3年生 31.94±1.77 aA 1.18±0.27 aA 7.34±0.80 aA 8.00±1.42 aA 1.51±0.32 aA 平均 31.20±1.53 A 1.18±0.07 A 7.49±0.06 A 6.55±0.53 A 1.52±0.17 A 枝 1年生 15.88±1.48 aB 0.58±0.08 aB 4.42±0.24 aB 0.67±0.18 aB 0.26±0.01 aB 2年生 13.70±0.32 bB 0.47±0.13 aC 2.37±0.17 bC 0.49±0.10 aB 0.10±0.03 bB 3年生 10.56±1.09 cB 0.22±0.04 bC 1.47±0.07 cC 0.68±0.15 aB 0.11±0.05 bB 平均 13.38±0.52 B 0.42±0.06 C 2.75±0.13 C 0.61±0.07 B 0.16±0.02 B 秆 1年生 9.52±1.79 aC 0.54±0.11 aB 6.30±0.53 aA 0.26±0.02 bB 0.18±0.01 bB 2年生 9.33±1.21 aC 0.79±0.28 aB 4.03±0.32 bB 0.47±0.10 bB 0.21±0.04 abB 3年生 10.79±0.84 aC 0.61±0.12 aB 3.48±0.37 bB 1.13±0.52 aB 0.40±0.16 aB 平均 9.88±1.17 C 0.65±0.17 B 4.60±0.18 B 0.62±0.15 B 0.27±0.06 B 部位 竹龄 Fe/(mg·kg−1) Mn/(mg·kg−1) Cu/(mg·kg−1) Zn/(mg·kg−1) 叶 1年生 282.01±24.01 aA 551.68±53.53 aA 7.22±0.36 aA 38.65±7.87 aA 2年生 177.98±30.25 bA 519.09±53.53 aA 6.43±0.33 bA 35.16±6.50 aA 3年生 152.25±10.06 bA 567.65±34.83 aA 5.72±0.18 bA 38.00±8.42 aB 平均 204.08±21.44 A 546.14±47.30 A 6.46±0.29 A 37.27±7.6 A 枝 1年生 39.45±10.03 bC 165.74±50.11 aB 3.98±1.13 aB 25.31±2.75 bB 2年生 38.02±6.89 bC 166.21±89.96 aB 2.86±0.32 abB 23.07±4.33 bB 3年生 121.13±38.06 aA 228.66±111.23 aB 2.10±0.08 bB 36.88±3.33 aB 平均 66.20±18.33B 186.87±83.76B 2.98±0.51 B 28.42±3.47 B 秆 1年生 88.13±20.34 aB 46.39±12.99 bC 2.37±0.15 aC 29.86±5.76 aB 2年生 95.86±19.35 aB 124.30±34.72 abB 2.36±0.38 aB 33.67±6.68 aA 3年生 79.34±6.61 aB 301.28±79.33 aB 1.79±0.26 aB 49.11±7.58 aA 平均 87.78±15.43 B 157.32±42.34 B 2.17±0.27 B 37.55±6.67 A 说明:小写字母表示不同竹龄间差异显著(P<0.05),大写字母表示不同部位间差异显著(P<0.05)
下载: 导出CSV
表 3 黄甜竹不同部位各营养元素的积累量
Table 3. Accumulation of nutrient elements in different parts of A. edulis
部位 N/
(kg·hm−2)P/
(kg·hm−2)K/
(kg·hm−2)Ca/
(kg·hm−2)Mg/
(kg·hm−2)Cu/
(kg·hm−2)Zn/
(kg·hm−2)Fe/
(kg·hm−2)Mn/
(kg·hm−2)合计/
(kg·hm−2)叶 212.18 8.00 50.37 47.00 10.19 0.04 0.25 1.27 3.72 333.01 枝 126.98 3.90 25.22 5.87 1.47 0.03 0.28 0.69 1.85 166.29 秆 417.91 27.21 191.10 27.34 11.50 0.09 1.61 3.68 6.95 687.38 合计 757.06 39.11 266.69 80.21 23.16 0.16 2.14 5.63 12.52 1 186.69
下载: 导出CSV
-
[1] SHARMA J C, SHARMA Y. Nutrient cycling in forest ecosystems: a review [J]. Agric Rev, 2004, 25(3): 157 − 172. [2] 何林. 中国黄甜竹[M]. 北京: 中国林业出版社, 2012. [3] 谢益贵, 叶树军, 张世平. 高山引种黄甜竹试验初报[J]. 浙江林业科技, 2001, 21(4): 38 − 40. XIE Yigui, YE Shujun, ZHANG Shiping. Preliminary report on introduction of Acidosasa edulis on hills [J]. J Zhejiang For Sci Tech, 2001, 21(4): 38 − 40. [4] 陈松河, 郑清芳. 黄甜竹笋用林丰产培育技术模式的研究[J]. 竹子研究汇刊, 2001, 20(1): 61 − 67. CHEN Songhe, ZHENG Qingfang. Technical study on high-yield management pattern of shoot stand of Acidosasa edulis [J]. J Bamboo Res, 2001, 20(1): 61 − 67. [5] 何林, 傅冰, 王军峰, 等. 黄甜竹丰产林地下竹鞭结构生长规律研究[J]. 竹子研究汇刊, 2011, 30(3): 17 − 21. HE Lin, FU Bing, WANG Junfeng, et al. Growth pattern of the rhizome system in high-yield stands of Acidosasa edulis [J]. J Bamboo Res, 2011, 30(3): 17 − 21. [6] 吕兆林. 竹叶黄酮和挥发油的制备及生物活性的研究[D]. 北京: 北京林业大学, 2009. LÜ Zhaolin. Characterization, Production and Bioactivity of Flavonoids and Volatile Oil from Bamboo Leaves[D]. Beijing: Beijing Forestry University, 2009. [7] 周芳纯. 竹林培育学[M]. 北京: 中国林业出版社, 1998: 397 − 399. [8] 夏传格, 宁晨, 罗赵慧, 等. 不同年龄毛竹林养分分布及生物循环特征[J]. 生态学报, 2020, 40(11): 3715 − 3725. XIA Chuange, NING Chen, LUO Zhaohui, et al. Nutrient distribution and biochemical cycling in different aged moso bamboo (Phyllostachys pubescens) ecosystems [J]. Acta Ecol Sin, 2020, 40(11): 3715 − 3725. [9] 吴家森, 周国模, 钱新标, 等. 不同经营类型毛竹林营养元素的空间分布[J]. 浙江林学院学报, 2005, 22(5): 486 − 489. WU Jiasen, ZHOU Guomo, QIAN Xinbiao, et al. Distribution of nutrient elements in different organs of Phyllostachys pubescens under different managements [J]. J Zhejiang For Coll, 2005, 22(5): 486 − 489. [10] 宋艳冬, 金爱武, 金晓春, 等. 施肥对毛竹叶片光合生理的影响[J]. 浙江林学院学报, 2010, 27(3): 334 − 339. SONG Yandong, JIN Aiwu, JIN Xiaochun, et al. Physiology of leaf photosynthesis with fertilization in Phyllostachys pubescens [J]. J Zhejiang For Coll, 2010, 27(3): 334 − 339. [11] 黄张婷, 姜培坤, 宋照亮, 等. 不同竹龄雷竹中硅及其他营养元素吸收和积累特征[J]. 应用生态学报, 2013, 24(5): 1347 − 1353. HUANG Zhangting, JIANG Peikun, SONG Zhaoliang, et al. Uptake and accumulation characteristics of silicon and other nutritional elements in different age Phyllostachys praecox plants [J]. Chin J Appl Ecol, 2013, 24(5): 1347 − 1353. [12] 刘力, 林新春, 金爱武, 等. 苦竹各器官营养元素分析[J]. 浙江林学院学报, 2004, 21(2): 172 − 175. LIU Li, LIN Xinchun, JIN Aiwu, et al. Analysis of nutrient elements in various organs of Pleioblastus amarus [J]. J Zhejiang For Coll, 2004, 21(2): 172 − 175. [13] 叶晶, 陶立华, 柯和佳, 等. 绿竹地上部营养元素的吸收、积累和分配特性[J]. 浙江农林大学学报, 2015, 32(4): 545 − 550. YE Jin, TAO Lihua, KE Hejia, et al. Uptake, accumulation, and distribution of the main elemental nutrients in Dendrocalamopsis oldhami [J]. J Zhejiang A&F Univ, 2015, 32(4): 545 − 550. [14] 葛高波, 高智群, 刘欢, 等. 黄竹地上部分营养元素的吸收、积累和分配特性[J]. 竹子研究汇刊, 2016, 35(1): 41 − 45, 57. GE Gaobo, GAO Zhiqun, LIU Huan, et al. Uptake, accumulation and distribution of main elemental nutrients in the aboveground parts of Dendrocalamus membranceus [J]. J Bamboo Res, 2016, 35(1): 41 − 45, 57. [15] 叶晶, 葛高波, 应雨骐, 等. 青皮竹地上部营养元素的吸收、积累和分配特性研究[J]. 植物营养与肥料学报, 2015, 21(1): 154 − 160. YE Jing, GE Gaobo, YING Yuqi, et al. Absorption, accumulation and distribution of bamboo (Bambusa textilis) to the main nutrients [J]. J Plant Nutr Fert Sci, 2015, 21(1): 154 − 160. [16] WU Jiasen, XU Qiufang, JIANG Peikun, et al. Dynamics and distribution of nutrition elements in bamboos [J]. J Plant Nutr, 2009, 32(3): 489 − 501. [17] 石元值, 马立峰, 韩文炎, 等. 茶叶中磷、钾、铅、锌等17种元素的快速测定方法研究[J]. 食品科学, 2006, 27(1): 193 − 196. SHI Yuanzhi, MA Lifeng, HAN Wenyan, et al. Study on simultaneous determination of phosphorus, potassium, lead, zinc etc. altogether 17 elements in tea [J]. Food Sci, 2006, 27(1): 193 − 196. [18] 陈迪, 潘伟槐, 周哉材, 等. 植物营养元素运输载体的功能及其调控机制研究进展[J]. 浙江大学学报(农业与生命科学版), 2018, 44(3): 283 − 293. CHEN Di, PAN Weihuai, ZHOU Zaicai, et al. Research progresses on functional roles and regulation mechanisms of nutrient element transporters in plants [J]. J Zhejiang Univ Agric Life Sci, 2018, 44(3): 283 − 293. [19] 佟志龙, 陈奇伯, 王艳霞, 等. 不同林龄云南松林营养元素积累与分配特征研究[J]. 西北农林科技大学学报(自然科学版), 2014, 42(6): 100 − 106, 114. TONG Zhilong, CHEN Qibo, WANG Yanxia, et al. Accumulation and distribution characteristics of nutrients in Pinus yunnanensis forests with different ages [J]. J Northwest A&F Univ Nat Sci Ed, 2014, 42(6): 100 − 106, 114. [20] 储晓燕, 杨杰, 叶柳欣, 等. 高节竹地上器官氮、磷、钾含量及积累特性[J]. 福建林业科技, 2018, 45(3): 19 − 21. CHU Xiaoyan, YANG Jie, YE Liuxin, et al. Content and accumulation of N, P, K in above-ground organs of Phyllostachys prominens [J]. J Fujian For Sci Tech, 2018, 45(3): 19 − 21. [21] 陈瑞国, 彭超, 刘广路, 等. 不同年龄慈竹养分地上空间分布特征及叶养分与土壤肥力关系[J]. 四川农业大学学报, 2017, 35(3): 353 − 358. CHEN Ruiguo, PENG Chao, LIU Guanglu, et al. Above-ground spatial distribution feature of nutrients in different ages of Neosinocalamus affinis and correlation between soil and leaves nutrients [J]. J Sichuan Agric Univ, 2017, 35(3): 353 − 358. [22] 史瑞和. 植物营养原理[M]. 南京: 江苏科学技术出版社, 1989: 217 − 398. [23] 秦武明, 何斌, 覃世赢, 等. 厚荚相思人工林营养元素生物循环的研究[J]. 水土保持学报, 2007, 21(4): 103 − 107. QIN Wuming, HE Bin, QIN Shiying, et al. Biological cycling of nutrients in Acacia crassicarpa plantation [J]. J Soil Water Conserv, 2007, 21(4): 103 − 107. [24] 刘鹏, 郝朝运, 陈子林, 等. 不同群落类型中七子花器官营养元素分布及其与土壤养分的关系[J]. 土壤学报, 2008, 45(2): 304 − 312. LIU Peng, HAO Chaoyun, CHEN Zilin, et al. Nutrient element distribution in organs of Heptacodium miconioides in different communities and its relationship with soil nutrients [J]. Acat Pedol Sin, 2008, 45(2): 304 − 312. [25] 孙明茂, 洪夏铁, 李圭星, 等. 水稻籽粒微量元素含量的遗传研究进展[J]. 中国农业科学, 2006, 39(10): 1947 − 1955. SUN Mingmao, HONG Hachoel, LEE Kyuseong, et al. Progress of genetic research on trace minerals content in rice seed [J]. Sci Agric Sin, 2006, 39(10): 1947 − 1955. [26] 杨杰, 吴家森, 姜培坤, 等. 苦竹林植硅体碳与硅的研究[J]. 自然资源学报, 2016, 31(2): 299 − 309. YANG Jie, WU Jiasen, JIANG Peikun, et al. Study on phytolith-occluded organic carbon and silicon in a Pleioblastus amarus forest [J]. J Nat Resour, 2016, 31(2): 299 − 309. [27] 魏琦, 姚曦, 汤锋, 等. 苦竹属竹叶矿质元素比较研究[J]. 竹子研究汇刊, 2014, 34(3): 43 − 46. WEI Qi, YAO Xi, TANG Feng, et al. Mineral elements in bamboo leaves of ten Pleioblastus species [J]. J Bamboo Res, 2014, 34(3): 43 − 46. [28] WALLACE T C, MCBURNEY M, FULGONI V L. Multivitamin/mineral supplement contribution to micronutrient intakes in the United States, 2007−2010 [J]. J Am Coll Nutr, 2014, 33(2): 94 − 102. [29] 何跃君. 竹叶挥发油化学成分及其生物活性研究[D]. 北京: 中国林业科学研究院, 2009. HE Yuejun. Chemical Components and Biological Activities of Essential Oils from Bamboo Leaves[D]. Beijing: Chinese Academy of Forestry, 2009. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200646
 点击查看大图
点击查看大图
计量
- 文章访问数: 1575
- HTML全文浏览量: 355
- PDF下载量: 20
- 被引次数: 0
