





-
中国竹类资源相对丰富。竹材刚度好、强度大,而且化学成分与木材类似,主要为纤维素、半纤维素和木质素[1-3],因此,竹材是木材最佳的代替品[4]。竹材易变形,抗腐霉能力差[5],借鉴木材热处理的方法对竹材进行高温热处理,以达到改良竹材的品质,降低竹材的吸湿性和吸水性,提高尺寸稳定性、生物耐腐性和耐候性。改性处理是纯物理热处理技术,与其他化学改性方法相比,生产过程中污染少,处理工艺较简单,且使用过程中不会因化学药剂的流失、挥发而降低防腐效果,也不会对人体造成伤害[6]。竹材化学成分以及分布是影响竹材材性和利用的重要因素[7],所以,本研究以竹材处理的温度和时间为因素,探讨分析热处理温度和时间对竹材中的主要化学成分:综纤维素、α-纤维素、木质素质量分数的影响。已有研究表明[8]:热处理温度对竹材性能的影响比时间更为显著。为了确定合理的热处理温度,在红外光谱实验时选择了4 h作为热处理时间来分析热处理后竹材官能团的变化,以期为热处理工艺对竹材力学性能影响的研究提供一定的参考,并对运用高温热处理改性竹材生产优质竹产品提供了借鉴。
-
毛竹Phyllostachys edulis材:取自临安市东湖村,取距地面约为2.0 m的整竹节处以1.5 m为长度,2个竹节间长的4年生竹材。将气干后的无节竹材制作成300片尺寸为600 mm × 20 mm × t(t为壁厚)的坯料试件,烘干,控制含水率在9 ~12 g·kg-1再进行热处理。热处理后竹材加工成尺寸为80 mm × 20 mm × t的竹片,共140个试样,供力学性能测试。毛竹粉:经过上述相同取材热处理后,放入粉碎机中将其磨成竹粉,获取过40目但不过60目筛的竹粉(约50 g),烘干,供化学成分分析及红外光谱使用。
-
中国上海梅特勒-托列多仪器有限公司AB204-N型万分之一电子分析天平,中国长春试验机研究所有限公司DNS50型微机控制电子式木材万能试验机,日本岛津公司IRPrestige-21型傅立叶变换红外光谱仪,自制小型木材热处理装置[9]等。
-
将烘干后的试件放入自制小型木材热处理装置[9]中采用温度为160,180和200 ℃,时间为2,4和6 h 进行热处理试验。
-
将未处理毛竹竹材与热处理毛竹竹材分别进行化学成分检测。综纤维素、α-纤维素和木质素3种成分分别参照GB/T 2677.10-1995《造纸原料综纤维素含量的测定》、GB/T 744-1989 《纸浆α-纤维素的测定》和GB/T 2677.8-1994 《造纸原料酸不溶木素含量的测定》[10]中规定的方法,进行2次测定,比较分析得出实验结果。
-
傅立叶变换红外光谱分析法能定性描述热处理材料官能团的变化,而材料官能团的变化与其自身的化学成分与物理力学性能有着内在的联系。为了考察热处理工艺对毛竹竹材性能的影响,本实验组主要选取160,180,200 ℃,4 h的竹粉进行红外光谱检测,来间接描述热处理竹材的性能变化。首先取少量改性毛竹竹粉在玛瑙缽中充分磨细,再加入干燥的溴化钾(KBr)粉末[m(竹粉)∶m(溴化钾)=1∶100)],继续磨研,直到完全混合均匀,并将混合物在红外灯下烘干,取混合物于压膜内,在压片机上用60 MPa的压力压2 min,然后泄压取出,即可得一透明薄片,把此薄片装于薄片夹持器上,然后在傅立叶变换红外光谱仪上进行测定分析。
-
第1组热处理毛竹竹材试件进行力学性能测试。热处理竹材的力学测试方法参照GB/T 15780-1995《竹材物理力学性质试验方法》,实验试样12个·组-1,比较分析得出实验结果。
-
未处理毛竹竹材木质素的质量分数为201.7 g·kg-1,综纤维素的质量分数为747.1 g·kg-1,α-纤维素的质量分数为453.5 g·kg-1。高温热处理后毛竹竹材的化学成分大致变化为:随着热处理温度的升高和热处理时间的延长,木质素的质量分数上升,综纤维素和α-纤维素的质量分数下降。
-
热处理后毛竹竹材综纤维素(纤维素和半纤维素)的质量分数变化见图 1。由图 1可知:热处理后毛竹竹材的综纤维素质量分数随着处理温度的升高和处理时间的延长而下降,主要因为综纤维素是多糖组成,在高温条件下这些多糖容易裂解为糖醛和某些糖类的裂解产物,随着高温热处理温度的升高裂解加剧[11]。
当热处理温度为160~180 ℃,毛竹竹材综纤维素的质量分数随着时间的增加而下降了71.1 g·kg-1,降幅较小,但其质量分数较未处理毛竹竹材均有所升高,其原因主要为热处理过程中水分蒸发,毛竹竹材平衡含水率降低以及部分易挥发性物质损失[12];而当热处理温度在180 ℃以上时,综纤维素分解加剧,其质量分数下降了73.6 g·kg-1;在处理时间超过4 h后其质量分数低于未处理材的。这可能是由于热处理过程中综纤维素基团受热,使分子内部脱水,其活性增强生成少量链引发基团,链引发基团大量分解,使得综纤维素质量分数低于未处理材。

图 1 热处理后毛竹竹材料综纤维素的变化
Figure 1. Changes of the content of holo-cellulose after heat treatment
-
热处理后毛竹竹材α-纤维素质量分数的变化见图 2。根据图 2所示:热处理温度对α-纤维素质量分数起决定性影响。当热处理的温度为160~180 ℃,α-纤维素变化不明显,主要是因为在160~180 ℃,半纤维素开始分解,而纤维素基本没有发生变化,对α-纤维素影响较小。当温度达到180 ℃以上时,随着热处理温度的升高和热处理时间的延长,纤维素也发生降解,导致α-纤维素质量分数呈现显著下降的趋势。当温度达到200 ℃时,其质量分数降低至未处理材的质量分数。热处理后竹材内部的纤维素的结晶结构减少,结晶区比重减少[13],此外,还会影响竹材的力学性能。

图 2 热处理后毛竹竹材α-纤维素的变化
Figure 2. Changes of the content of α-cellulose after heat treatment
-
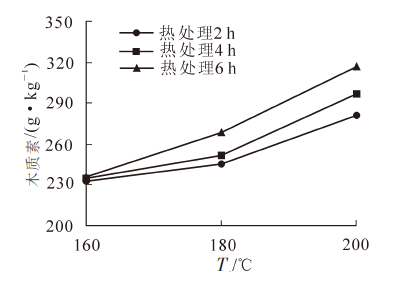
热处理后毛竹竹材木质素的质量分数变化见图 3。由图 3可知:当热处理温度为160~180 ℃时,热处理后毛竹竹材的木质素质量分数变化不明显,但高于未处理材。当热处理温度在180 ℃以上时,木质素质量分数随热处理温度的升高和热处理时间的延长呈显著上升的趋势。有学者认为:木质素在热处理的过程中发生了缩聚反应,聚合度和结构均发生了变化,并由此产生网状交联结构。也有研究表明,主要是由于半纤维素和纤维素发生降解造成木质素质量分数的增加。Elisabeth等[14]通过实验发现,木质素降解后生成木酚素,其木酚素使得热处理后的竹材耐久性能提高,聚糖质量分数的降低也有助于竹材的耐久性提高。

图 3 热处理后毛竹竹材木质素的变化(t=4 h)
Figure 3. Changes of the content of Klason-lignin after heat treatment (t=4 h)
-
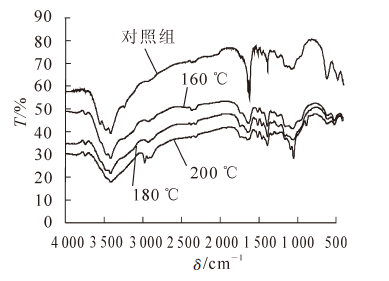
高温热处理前后毛竹竹材的红外光谱图见图 4。不同热处理工艺会造成毛竹竹材化学成分变化程度不同[15]。从图 4可见:3420 cm-1和1745 cm-1附近的吸收峰震动最为剧烈,高温热处理后竹材的羟基峰和羰基峰的吸收强度均呈现降低趋势。3420 cm-1附近的吸收峰为O-H伸展振动吸收峰,从图 4中可见:随着热处理温度的升高,与未处理材相比毛竹竹材羟基峰变弱,说明经过热处理后羟基数目减少。这主要是由于毛竹竹材纤维素、半纤维素、木质素上均含有大量的羟基基团,羟基数量减少,半纤维素和纤维素发生降解,从而质量分数下降。在高温作用下,羟基基团发生氧化反应,生成醛基、酮基或羧基从而使游离羟基的数量减少;此外,在酸性条件下(多聚糖水解产生醋酸),细胞壁物质中的木质素发生酯化反应,使得羟基数量再次减少。1745 cm-1附近的吸收峰为C-O伸缩振动(木聚糖乙酰基CH3C=O),随着热处理温度升高,吸收峰的吸收强度变弱,因为经过高温处理后木聚糖发生降解。当温度达到160 ℃或更高时羰基发生分裂。在上述因素共同作用下,高温热处理后毛竹竹材羟基峰和羰基峰的吸收强度均呈降低趋势。木质素在热处理过程中发生了化学变化,在红外光谱中1600 cm-1附近的吸收峰为木质素C=O伸缩振动吸收峰,从峰谱图可以看出吸收峰随温度的升高而下降。

图 4 高温热处理前后毛竹竹材的红外光谱图
Figure 4. Spectrogram of FT-IR specbefore and after heat treatment (t=4 h)
-
热处理温度对毛竹竹材绝干密度的影响见表 1。毛竹竹材在50 g·kg-1左右含水率的力学测试中可以看出,随着热处理温度的升高,热处理时间的延长,毛竹竹材的绝干密度呈下降的趋势。主要原因是毛竹竹材中化学成分发生了变化,三大素发生降解;并且红外光谱图显示羟基数量下降,则毛竹竹材的吸湿能力下降,从而影响密度。
热处理工艺不仅改变了毛竹竹材的化学成分,还影响了它的力学性能。本实验中毛竹竹材抗弯强度和抗弯弹性模量都是在一定的温湿度环境下测得。热处理温度对毛竹竹材抗弯强度和抗弯弹性模量的影响见表 1。可知,毛竹竹材的抗弯强度随热处理温度的升高与热处理时间的延长均呈下降趋势;而抗弯弹性模量随热处理温度的升高先增大后减小,随热处理时间的延长呈下降趋势。热处理工艺为200 ℃和6 h的改性毛竹竹材与未处理材相比,其抗弯强度减少84.5 MPa,抗弯弹性模量减小1.86 GPa。
竹材内部纤维素发生降解是竹材力学性能降低的最主要原因。由图 1,图 2和图 3可知:随着热处理温度的升高与热处理时间的延长,竹材内部主要化学成分均发生降解,热处理200 ℃和6 h的改性毛竹竹材与未处理竹材相比,综纤维素质量分数下降了53.5 g·kg-1,α-纤维素的质量分数下降了239.4 g·kg-1。由红外图谱可知,随着热处理温度的升高,竹材的羟基峰值下降,经过热处理后纤维素发生降解,羟基等强吸水性基团数目减少,在化学成分的作用下导致竹材结晶度减少,从而影响竹材的力学性能。
竹材的抗弯弹性模量的大小主要是由于密度的改变[16],继而影响内部的结晶结构。一般情况下,抗弯弹性模量与密度成正比。正如表 1所示:随着热处理温度升高,毛竹竹材内部结晶区晶层距离变小,结晶区变大,结晶结构变得更为紧凑,因而其抗弯弹性模量出现了增大的现象。当密度下降时,抗弯弹性模量出现下降的趋势。也有分析指出:竹材在160 ℃和2 h的热处理过程中半纤维素中的木聚糖与甘露聚糖产生了结晶化,因此导致结晶度增大[17-18],随着热处理时间的延长,化学成分降解剧烈,对力学性能的影响更为明显,从而导致了毛竹竹材抗弯弹性模量也先增大后减小。
表 1 热处理前后毛竹竹材物理力学性能的变化
Table 1. Changes of physical and mechanical properties of the bamboo before and after heat treatment
T/℃ t/h p/(g·cm-1) 抗弯弹性强度/MPa 抗弯弹性模量/GPa 室温 0 0.799 203.6 12.98 160 2 0.790 189.9 12.55 160 4 0.791 162.9 11.95 160 6 0.777 132.6 10.58 180 2 0.787 187.7 14.83 180 4 0.765 171.1 14.98 180 6 0.754 156.9 14.76 200 2 0.780 142.1 12.92 200 4 0.748 128.9 11.98 200 6 0.749 119.1 11.12 -
不同热处理工艺对毛竹竹材的化学成分和力学性能影响较大,利用热处理工艺可以制造不同性能的竹材产品,从而满足不同使用范围产品的需求。①随着热处理温度升高和热处理时间延长,毛竹竹材发生热降解、缩聚等化学反应,使得纤维素、半纤维素的质量分数下降,木质素的质量分数呈现上升的趋势。200 ℃和6 h的热处理竹材与未处理竹材相比,综纤维素质量分数下降了93.1 g·kg-1,α-纤维素的质量分数下降了239.4 g·kg-1,木质素质量分数上升了115.0 g·kg-1。②随着热处理温度升高和热处理时间延长,热处理竹材抗弯强度下降,抗弯弹性模量略微上升后呈下降趋势。其中200 ℃和6 h热处理毛竹竹材抗弯强度较未处理材减小了84.5 MPa,抗弯弹性模量较未处理材减小了1.86 GPa。热处理温度越高和热处理时间越长,毛竹竹材的密度,抗弯强度,抗弯模量下降的更为显著。
Phyllostachys edulis with high temperature heat treatments
-
摘要: 采用热处理温度为160,180,200℃,热处理时间为2,4,6 h的高温热处理工艺对毛竹Phyllostachys edulis竹材进行改性处理,分析不同热处理工艺对竹材化学成分和力学性能的影响,将分别在160,180,200℃下处理4 h后的竹材进行傅立叶变换红外光谱图表征。结果表明:热处理温度越高和时间越长,竹材中木质素质量分数也越高,综纤维素、α-纤维素质量分数呈现下降的趋势,竹材的纵向抗弯强度呈减小趋势,并且抗弯弹性模量呈减小趋势。200℃,6 h热处理竹材与未处理竹材相比,木质素质量分数上升了115.0 g·kg-1,综纤维素质量分数下降了93.1 g·kg-1,α-纤维素的质量分数下降了239.4 g·kg-1,毛竹竹材的抗弯强度较未处理材减小了84.5 MPa,抗弯弹性模量较未处理材减小了1.86 GPa。红外谱图中竹材表面羟基数目随热处理温度的上升和热处理时间的延长不断减少。Abstract: To determine changes in chemical components as well as physical and mechanical properties of Phyllostachys edulis, heat treatments of 160,180,and 200℃ for 2,4,and 6 h were applied to bamboo and compared with untreated bamboo. The treatments were analyzed with a spectrogram from a Fourier transform infrared spectroscopy. Results showed that as the heat treatment temperatures and times increased, the Klason-lignin content of Ph. edulis bamboo increased;whereas holo-cellulose content,α-cellulose,modulus of rupture (MOR),and modulus of elasticity (MOE) decreased. The 200℃ and 6 h heat-treatment compared to untreated bamboo increased the Klason-lignin content 115.0 g·kg-1 but decreased the holo-cellulose content 93.1 g·kg-1 and α-cellulose 239.4 g·kg-1. Bending strength decreased 84.5 MPa and the MOE decreased 1.86 GPa. The spectrogram also illustrated that the number of hydroxyls on the surface of the modified bamboo decreased.
-
Key words:
- woody science /
- Phyllostachys edulis /
- heat treatment /
- chemistry components /
- FT-IR spectrum /
- mechanical properties
-
图 1 热处理后毛竹竹材料综纤维素的变化
Figure 1 Changes of the content of holo-cellulose after heat treatment
图 2 热处理后毛竹竹材α-纤维素的变化
Figure 2 Changes of the content of α-cellulose after heat treatment
图 3 热处理后毛竹竹材木质素的变化(t=4 h)
Figure 3 Changes of the content of Klason-lignin after heat treatment (t=4 h)
图 4 高温热处理前后毛竹竹材的红外光谱图
Figure 4 Spectrogram of FT-IR specbefore and after heat treatment (t=4 h)
表 1 热处理前后毛竹竹材物理力学性能的变化
Table 1. Changes of physical and mechanical properties of the bamboo before and after heat treatment
T/℃ t/h p/(g·cm-1) 抗弯弹性强度/MPa 抗弯弹性模量/GPa 室温 0 0.799 203.6 12.98 160 2 0.790 189.9 12.55 160 4 0.791 162.9 11.95 160 6 0.777 132.6 10.58 180 2 0.787 187.7 14.83 180 4 0.765 171.1 14.98 180 6 0.754 156.9 14.76 200 2 0.780 142.1 12.92 200 4 0.748 128.9 11.98 200 6 0.749 119.1 11.12  下载: 导出CSV
下载: 导出CSV
-
[1] 杨淑惠. 植物纤维化学[M]. 北京:中国轻工业出版社,2001:65-67. [2] 张齐生. 中国竹材工业化利用[M]. 北京:中国林业出版社,1995:39-41. [3] 刘一星,赵广杰. 木质资源材料学[M]. 北京:中国林业出版社,2004:246-252. [4] 薛涵与. 竹纤维的表面改性及其复合材料的研究[D]. 福州:福建师范大学,2010. XUE Hanyu. Surface Modification of Bamboo Fiber and its Composites[D]. Fuzhou:Fujian Normal University,2010. [5] 张亚梅,于文吉. 热处理对竹材性能及变色的影响[J]. 木材加工机械,2008(4):40-47. ZHANG Yamei,YU Wenji. Research about heat-treatment effect on bamboo performance and discoloration[J]. Woodwork Mach,2008(4):40-47. [6] 李延军,唐荣强,鲍滨福,等. 高温热处理杉木力学性能与尺寸稳定性研究[J]. 北京林业大学学报,2010,32(4):232-236. LI Yanjun,TANG Rongqiang,BAO Binfu,et al. Mechanical properties and dimensional stability of heat-treated Chinese fir[J]. J Beijing For Univ,2010,32(4):232-236. [7] 熊文愈. 竹材构造和性质报告[J]. 竹类研究,1991,10(1):1-6. XIONG Wenyu. Structure and nature report of bamboo[J]. Bamboo Res,1991,10(1):1-6. [8] 张亚梅. 热处理对竹材颜色及物理力学性能影响的研究[D]. 北京:中国林业科学研究院,2010. ZHANG Yamei. Study on the Effect of Color and Physical-mechanical Properties for Heat-treated Bamboo[D]. Beijing:Chinese Academy of Forestry, 2010. [9] 李延军,唐荣强,鲍滨福,等. 高温热处理木材工艺的初步研究[J]. 林产工业,2008,35(2):16-18. LI Yanjun,TANG Rongqiang,BAO Binfu,et al. The preliminary study on high temperature heat-treated technology of wood[J]. For Prod Ind,2008,35(2):16-18. [10] 国家轻工业局行业管理质量标准处. 中国轻工业标准汇编:造纸卷[S]. 北京:中国标准出版社,1999. [11] 成俊卿. 木材学[M]. 北京:中国林业出版社,1985:325-332. [12] 张亚梅,余养伦,于文吉,等. 热处理对毛竹化学成分变化的影响[J]. 中国造纸学报,2011,26(2):6-10. ZHANG Yamei,YU Yanglun,YU Wenji, et al. Effect of heat treatment on the chemical composition of bamboo[J]. Trans China Pulp Paper,2011,26(2):6-10. [13] BHUIYAN M T R, HIRAI N, SOBUE N. Changes of crystallinity in wood cellulose by heated treatment under dried and moist conditions[J]. J Wood Sci,2000,46(6):431-436. [14] WINDEISEN E, STROBEL C, WEGENER G. Chemical changes during the production of thermo-treated beech wood[J]. Wood Sci Technol,2007,41(6):523-536. [15] 胡够英. 高温热处理竹材动态黏弹性研究[D]. 临安:浙江农林大学,2012. HU Gouying. Study on Dynamic Viscoelasticities of Heat-treated Bamboo[D]. Lin'an:Zhejiang A&F University,2012. [16] 林勇,沈钰程,于利,等. 高温热处理竹材的物理力学性能研究[J]. 林业机械与木工设备,2012,40(8):22-24. LIN Yong,SHEN Yucheng,YU Li,et al. The study of high temperature heat treatment on physical-mechanical properties of bamboo[J]. For Mach&Woodwork Equ,2012,40(8):22-24. [17] 龙超,郝丙业,刘文斌,等. 影响热处理木材力学性能的主要工艺因素[J]. 木材工业,2008,22(1):43-45. LONG Chao,HAO Bingye,LIU Wenbin,et al. Effect of heat treatment on mechanical properties of wood[J]. China Wood Ind,2008,22(1):43-45. [18] TJEERDSMA B F,MILITZ H. Chemical changes in hydrothermal treated wood:FTIR analysis of combined hydrothermal and dry heat-treated wood[J]. Holz Roh-Werkst,2005,63:102-111. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2014.02.001
 点击查看大图
点击查看大图

计量
- 文章访问数: 4756
- HTML全文浏览量: 875
- PDF下载量: 941
- 被引次数: 0
