





-
山核桃Carya cathayensis隶属于胡桃科Juglandaceae中的山核桃属Carya,是一种著名的干果树种,因其干果富含营养和经济价值较高而得到了广泛栽培[1]。山核桃干腐病是山核桃生产上一种重要病害,不但影响山核桃的产量,而且会削弱树势,严重时则导致树木过早死亡,并造成重大经济损失[2]。2011年首次报道山核桃干腐病病原菌为Botryosphaeria dothidea(Moug. ex Fr.) Ces. & De Not,属于子囊菌门葡萄座腔菌科Botryosphaeriaceae的葡萄座腔菌属Botryosphaeria[3]。葡萄座腔菌科真菌是农业和林业上重要病原菌、内生真菌或潜在的致病菌,主要引起树木溃疡病。葡萄座腔菌属真菌广泛分布于世界各地,而且寄主范围广泛,是森林生态系统中的重要真菌类群[4]。该菌存在有性型和无性形阶段,其主要形态分类特征为子座、子囊、子囊孢子以及分生孢子的形状、纹饰、颜色、分隔、长宽比、大小及壁厚度等[5-6]。另外,培养菌落颜色、气生菌丝生长情况及子囊孢子表面超微结构(纹饰)也可用于葡萄座腔菌科真菌的分类和鉴定[7]。随着分子生物学的快速发展,越来越多的基因序列分析方法应用于葡萄座腔菌科真菌的分类鉴定及系统发育分析,如核糖体小亚基基因(SSU),核糖体大亚基基因(LSU),延长因子α基因(EF1-a),核糖体DNA内转录间隔区(rDNA-ITS),几丁质合酶基因,β微管蛋白(β-tubulin),A-肌动蛋白(A-actin)基因,钙调蛋白(calmodulin)基因等基因[8]。尤其rDNA-ITS序列是应用最普遍的基因序列,已广泛地应用于很多真菌目、科、属、种等的分类鉴定及系统发育分析。本研究采用形态学特征与与rDNA ITS相结合的方法对分离自中国山核桃的干腐病菌进行了鉴定研究。
-
分别从浙江省临安市的昌化镇和横路镇,淳安县,桐庐县和安徽省宁国市等山核桃产区采集干腐病标本(枝条和树干)带回实验室,然后在实验室进行病原菌分离与纯化培养。具体分离方法:首先选取发病枝条和树干,用乙醇对病健交接处的组织进行表面消毒30 s,然后剪成约5.0 mm × 5.0 mm大小的组织块,在体积分数为75.0%的乙醇中浸泡5 s,用无菌水浸洗3次,再用质量分数为1.0%的次氯酸钠浸泡1 min,最后用无菌水清洗3次。用灭菌的滤纸吸干水分,将组织块置于马铃薯葡萄糖琼脂培养基(PDA)平板上,在25 ℃恒温培养箱中进行培养,2 d后挑取菌落边缘的菌丝进行转接培养、纯化,并进行编号和转管保存。
-
将供试菌株在马铃薯葡萄糖琼脂(PDA)平板上活化培养3~4 d后,用经灭菌的直径为7.0 mm的打孔器打取菌饼。室外选取健康的山核桃,采用丁字型接种方法,将菌饼正面朝向伤口,用已浸湿无菌水脱脂棉保湿。设置重复15个·菌株-1,并设置空白PDA作为对照。接种15 d后观察发病情况,记录不同菌株的病斑数和发病级别,并计算不同菌株的感病指数。将病斑分为3级,15 d后病斑大小在10.0 mm以上的代表数值为“3”,10.0 mm以下5.0 mm以上代表数值的为“2”,5.0 mm以下的代表数值为“1”,不发病的代表数值为“0”。感病指数等于各病级的总代表数值(病斑分级的代表数值与该级标准株数之积)相加,再除以最高一级的代表数值与总株数之积,再乘以100。感病指数越高表示该菌株的致病性越强。对发病病斑进行组织分离,分离得到与接种菌株培养特征一致的菌株确定为该病的病原菌。
-
将分离获得的病原菌菌株在PDA平板上培养5 d后,观察菌落培养特性,将菌株接种到由树皮煎汁或松针等制成的培养基上诱导孢子产生。制作徒手切片后,在光学显微镜下观察和测量病原菌分生孢子器、分生孢子梗及分生孢子等特征。根据形态特征进行病原菌种类鉴定。
-
将病原菌转接到PDA平板上,于25 ℃培养3 d后,刮取约200.0 mg气生菌丝于灭菌后的1.5 mL的离心管中,-20 ℃冰冻过夜,经带研磨杵的电钻研磨破壁后,采用基因组DNA提取试剂盒(北京宝锐通生物科技有限公司)提取真菌基因组DNA。
-
PCR扩增和纯化 采用真菌rDNA-ITS区域通用引物ITS1(5′TCCGTAGGTGAACCTGCGG3′)和ITS4(5′TCCTCCGCTTATTGATATGC 3′)进行聚合酶链式反应(PCR)扩增。PCR反应体系总体积为25.0 μL,包括12.5 μL PCR脱氧核糖核苷三磷酸(dNTP),9.5 μL 双蒸水(ddH2O),引物ITS1/ITS4各1.0 μL,模板DNA 1.0 μL。反应程序:94.0 ℃预变性2 min;94.0 ℃变性30 s;57.3 ℃退火30 s;72.0℃延伸40 s,30个循环;72.0 ℃延伸10 min。经质量浓度为15.0 g·L-1琼脂糖凝胶电泳检测PCR扩增产物,将条带清晰的PCR扩增产物送交北京宝锐通生物科技有限公司进行双向测序。
-
将测序结果在美国生物技术信息中心(NCBI) GenBank ()中进行同源性比对,下载参比序列,采用clustalX和Mega 5.0软件进行分析比对,采用PAUP 4.0和Mrbayes 3.0软件构建最大简约法(MP)和贝叶斯法(BI)系统发育树。
-
本研究分离得到129株菌株,选取7株作为实验菌株,室外接种健康山核桃枝条,保湿培养15 d后,所有实验菌株均具有致病性,枝条上出现黑色病斑,对病斑进行组织分离得到了相应的病原菌。根据不同菌株枝条发病数及其感病级数,计算得到各病原菌的感病指数。其中CXY1565,CXY1566,CXY1567致病性相近,其病情指数为77.8,其致病性最强(图 1F),其次为CXY1568和CXY1569,病情指数为15.6(图 2F),CXY1570和CXY1571菌株致病性最差,其病情指数为6.7(图 3F)。

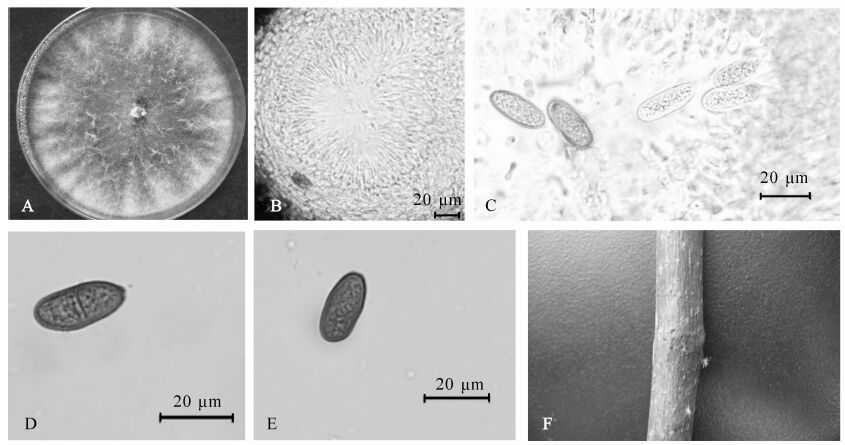
图 1 茶薦子葡萄座腔菌
Figure 1. Botryosphaeria dothidea (CXY1567)

图 2 Botryosphaeria fab icercianum(CXY1568)
Figure 2. Botryosphaeria fab icercianum(CXY1568)

图 3 Botryosphaeria obtusa(CXY1570)
Figure 3. Botryosphaeria obtusa(CXY1570)
-
以上7株病原菌形态学鉴定后共分为3种,一种是Botryosphaeria dothidea (Moug. ex Fr.) Ces. & De Not,以CXY1567为代表菌株,分离频率为71.4%;另一种是B. fabicercianum sp. Nov.,以CXY1568为代表菌株,分离频率为14.3 %;第3种是B. obtusa De Not.,以CXY1570为代表菌株,分离频率为14.3%。
Botryosphaeria dothidea:在PDA上25 ℃菌落生长速率较快,3~4 d几乎布满平板(直径90.0 mm);初期菌落白色或无色,气生菌丝棉絮毛状,较稀疏;培养1~2 d后,有白色或黄色小点沿菌丝分布,菌落中间有墨绿色色素;随着色素的沉积,黄色色素逐渐被掩盖,最后整个菌落变为灰褐色;后期菌落边缘气生菌丝倒伏紧贴培养基,菌落背面黄绿色色素呈点状不均匀分布;随着色素沉积,整个培养皿背面逐渐变为墨绿色或黑色。分生孢子器沿菌落边缘生长,表生,球形或不规则形;多腔室,分生孢子梗着生于腔室内壁细胞上,无色,杆状。分生孢子无色,无隔,锤形,顶部钝圆,基部比顶部稍尖。大分生孢子18.0~22.2 μm × 4.6~6.9 μm,平均为21.5 μm × 5.6 μm。长/宽比为3.0~4.0,小分生孢子直径为4.0~6.0 μm(图 1)。
Botryosphaeria fabicercianum:在PDA上菌落生长迅速,5 d可长满培养皿(直径90.0 mm)。菌落初呈白色,菌丝绒毛状或棉絮状,4~6 d菌落中央呈烟灰色,边缘菌丝紧贴培养基;12~16 d气生菌丝由灰绿色变为橄榄绿色,最后变为墨绿色。分生孢子器表生,散生或聚生,深褐色,球状,表面有菌丝覆盖。分生孢子器壁分3层,外层厚,深褐色或浅棕色,角质状;中层细胞薄壁,浅棕色;内层细胞薄壁,无色。分生孢子梗缺。产孢细胞圆柱形或烧瓶形,无色,光滑,薄壁,顶端产生单个分生孢子。侧丝无。分生孢子薄壁,光滑,无色,单胞,纺锤形,中间至中上1/3处最宽,顶端尖锐,基部平截,边缘具一个细小褶皱。分生孢子萌发前形成1~2个隔膜。大分生孢子17.3~24.3 μm × 4.5~7.5 μm,平均为22.7 μm × 6.1 μm,长/宽比为3.5~4.5,小分生孢子直径为3.8~6.3 μm[9](图 2)。
Botryosphaeria obtusa:在PDA上25 ℃生长迅速,3 d布满平板(直径90.0 mm),菌落初为白色,气生菌丝稀疏不发达,较短,细绒毛状,边缘整齐;2 d后有墨绿色色素沉积。后期气生菌丝分布于菌落边缘,稀疏且长势较弱,中央无气生菌丝,或气生菌丝平铺。10 d后由于色素沉积,菌落变为墨黑色,有时具反光;菌落背面由灰黑色变为墨绿色或者黑色。分生孢子器散生,表生,多腔室,腔室圆形或近圆形,无褶皱,内壁上着生分生孢子梗。分生孢子初无色,单胞,后呈褐色,卵形;大分生孢子为17.3~22.5 μm × 8.8~11.3 μm,平均为21.9 μm ×10.2 μm,长/宽比为1.8~2.3,小分生孢子直径3.0~4.0 μm[10](图 3)。
-
通过对供试7个菌株的rDNA-ITS序列测定和在GenBank中进行BLAST搜索和比对,结果表明:这些菌株均为葡萄座腔菌科真菌,分别为Botryosphaeria dothidea,B. fabicercianum和B. obtusa菌株(表 1)。
表 1 供试菌株rDNA-ITS序列与GenBank相关菌株的相似率
Table 1. Similarity of rDNA-ITS sequences of the tested fungal strains with related strains blasted in GenBank
菌株编号 菌株接收号 GenBank中相似的种类 相似率最高菌株 最高相似率/% 相似菌株接收号 CRY1567 TC527826 Botryosphaeria dothidea CBS 121484 99 EU650670 CRY1566 TC527822 B. dothidea CBS116743 99 AY786322 CRY1565 TC527825 B. dothidea CMW800 99 AY236949 CRY1568 TC527823 B. fabicercianum CMW24703 100 HQ332195 CRY1569 TC527824 B. fabicercianum CMW24703 99 HQ332195 CRY1570 TC527827 B. obtusa CBS119049 99 DQ458889 CRY1571 TC527828 B. obtusa CBS119049 99 DQ458889 -
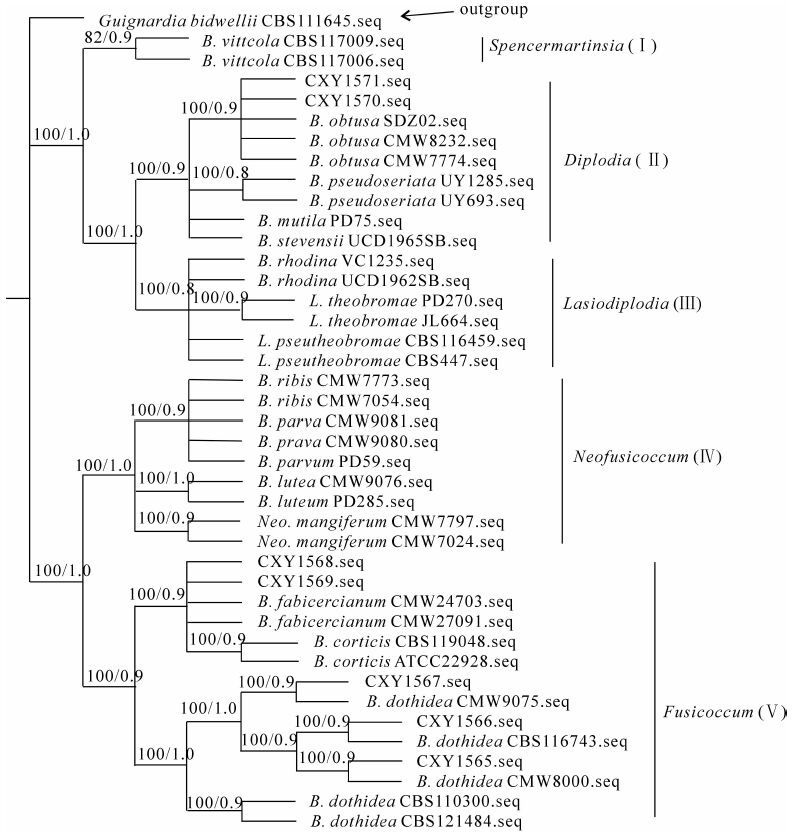
根据同源性比对的结果,从GenBank中下载33个与供试菌株关系相近的ITS序列和1个Guignardia philoprina(球痤菌属)序列作为外群,将所有序列整理后进行比对分析。用PAUP 4.0b10对比对结果进行最大简约法分析,将所有的614个特征视为无序且权重相同,其中120个恒量特征,和63个无效的变量特征Bootstrap法重复1 000次评估得到各节点支持率(BS)。利用最大简约法构建合议树步长(tree length)为327,一致性指数(consistency index,CI)0.8,保留指数(retention index,RI)0.9,趋同性指数(homoplasy index,HI)0.2,可调一致性指数(rescaled consistency index,RC)0.7;利用MrModeltese 3.7分析后,在AIC(Akaike Information Criterion)标准下,获得最佳模型TIM+G。贝叶斯方法采用马氏链蒙特卡罗(MCMC)算法,共运行500万代,所得9 902个树的合议树中各支的拓扑学结构与简约法基本一致,后验概率(PP)为节点支持率为PP。最后节点支持率为BS/PP(图 4)。

图 4 基于rDNA-ITS序列构建的系统发育树
Figure 4. The dendrogram constructed based on rDNA-ITS sequences
基于rDNA-ITS序列构建的系统发育树,结果将供试7个菌株与相关葡萄座腔菌科真菌分为2个大的类群,其中第1个类群包括分支Ⅰ,分支Ⅱ,分支Ⅲ,第2个类群包括分支Ⅳ和分支Ⅴ。
分支Ⅰ包括无性型为Spencermartinsia viticola等2个菌株,系加利福尼亚柑橘枝干溃疡病病菌[11]。供试菌株CXY1570和CXY1571位于分支Ⅱ中,其无性型为Diplodia。这2个菌株与B. obtusa菌株聚集在同一分支,与 CBS119049的菌株的最大相似率为99%。分支Ⅴ为无性型Fusicuccom类群的菌株,包括Botryosphaeria dothidea,B. fabicercianum和B. cortici。最大简约法和贝叶斯法的分析结果均表明,中国山核桃干腐病菌包括 Botryosphaeria dothidea,B. fabicercianum和B. obtusa。分子鉴定结果与形态学鉴定结果一致。
-
本研究从山核桃干腐病发病组织上分离得到了3种葡萄座腔菌属真菌,其中优势菌株为Botryosphaeria dothidea,而且致病性最强;而B. fabicercianum和B. obtusa分离频率较低,致病性较弱。
Botryosphaeria是重要的子囊菌,其无性型包括Diplodia,Dothiorella,Fusicoccum,Lasiodiplodia,Sphaeropsis等[12],是形态分类中最困难的真菌类群之一。关于山核桃干腐病的研究报道较少,而且关于病原菌种类不明确。杨淑贞等[13]提出该病病原真菌的有性态为B. fusisporae,无性态为Macrophoma caryae。也有研究认为山核桃溃疡病病原属于半知菌亚门腔胞纲球壳孢科小穴壳菌Dothiorella gregaria,并指出该病的病原与杨树溃疡病和桃树溃疡病的病原相同[14]。张传清等[15]认为山核桃干腐病菌为B. dothidea。田甜等[16]也认为山核桃干腐病病原菌是B. dothidea。本研究结果认为山核桃干腐病菌包括B. dothidea,B. fabicercianum及B. obtusa,但以B. dothidea为优势病菌。
Botryosphaeria是常见的林木干腐和枯梢病菌,尤其B. dothidea是发生最普遍和危害最重的病原菌[17]。Smith等[18]报道,B. dothidea在南非引起桉树溃疡病。另外,该菌是桉属Eucalyptus和松属Pinus植物上的内生真菌[19]。由于Botryosphaeria属真菌在自然条件以无性型最为常见,而且形态特征有限,所以,仅依靠形态特征难于进行种类鉴定。另外,该属真菌在人工培养条件下很难产生分生孢子器,而且耗时长。采用分子生物学技术不失为一种有效方法[20]。
Identification of the pathogens causing stem canker on Carya cathayensis
-
摘要: 葡萄座腔菌科Botryosphaeriaceae真菌是很多木本植物溃疡病和枯梢病的重要病原菌。已有报道认为:山核桃干腐病病原菌为茶藨子葡萄座腔菌Botryosphaeria dothidea,但对于葡萄座腔菌科其他真菌与山核桃干腐病的关系还没有深入研究。分离得到了山核桃干腐病相关病原菌,通过比较其形态学特征和核糖体DNA内转录间隔区(rDNA ITS)序列特征,将分离得到的供试菌株分为3组。根据形态学特征和基于ITS序列的系统发育分析,3组菌株分别鉴定为Botryosphaeria dothidea,B.fabicercianum和B.obtusa。其中B.dothidea为优势菌株,分离频率为71.42%,B.fabicercianum和B.obtusa分离频率分别为14.28%。经致病性测定,上述分离菌株均能引起健康山核桃Carya cathayensis枝条发病,但致病性存在差异。在山核桃上发现的B.obtusa和B.fabicercianum为首次报道。Abstract: Species of the Botryosphaeriaceae family are important pathogens causing cankers and die-back on many woody plants. In previous studies Botryosphaeria dothidea has commonly been reported as a pathogen of Chinese hickory(Carya cathayensis) canker; however, it was unclear whether other species of Botryosphaeriaceae were also related to hickory canker. In one study, several species of Botryosphaeriaceae were isolated from Chinese hickory, and then they were identified based on anamorphic morphology and rDNA Internal Transcribed Spacer(ITS) sequences. A phylogentic analysis indicated that these isolates formed three clades and were identified as B. dothidea, B. fabicercianum, and B. obtusa with isolation rates of B. dothidea (71.42%) predominant and B. fabicercianum and B. obtusa being much lower(14.28%). In pathogenicity testing, all three pathogens caused stem cankers on Chinese hickory but had different degrees of pathogenicity. B. obtusa and B. fabicercianum were first reported as a pathogen of Carya cathayensis canker.
-
Key words:
- forest protection /
- cash forestry /
- canker /
- Botryosphaeriaceae /
- Botryosphaeria dothidea /
- phylogeny /
- Carya cathayensis
-
图 1 茶薦子葡萄座腔菌
A:PDA上7 d菌落,B,C:载孢体(50 μm,20 μm);D:产孢细胞(10 μm);E:分生孢子(10 μm);F:发病症状。
Figure 1 Botryosphaeria dothidea (CXY1567)
图 2 Botryosphaeria fab icercianum(CXY1568)
A:PDA上7d菌落;B:载孢体横切面;C:萌发前的分生孢子(50μm);D:小分生孢子(10μm);E:大分生孢子(10μm);F:发病症状。
Figure 2 Botryosphaeria fab icercianum(CXY1568)
图 3 Botryosphaeria obtusa(CXY1570)
A:PDA上7 d菌落,B:载孢体(20μm); C:产孢细胞(20μm); D:有隔分生孢子(20μm); E:无隔分生孢子(20μm); F:发病症状。
Figure 3 Botryosphaeria obtusa(CXY1570)
图 4 基于rDNA-ITS序列构建的系统发育树
Figure 4 The dendrogram constructed based on rDNA-ITS sequences
表 1 供试菌株rDNA-ITS序列与GenBank相关菌株的相似率
Table 1. Similarity of rDNA-ITS sequences of the tested fungal strains with related strains blasted in GenBank
菌株编号 菌株接收号 GenBank中相似的种类 相似率最高菌株 最高相似率/% 相似菌株接收号 CRY1567 TC527826 Botryosphaeria dothidea CBS 121484 99 EU650670 CRY1566 TC527822 B. dothidea CBS116743 99 AY786322 CRY1565 TC527825 B. dothidea CMW800 99 AY236949 CRY1568 TC527823 B. fabicercianum CMW24703 100 HQ332195 CRY1569 TC527824 B. fabicercianum CMW24703 99 HQ332195 CRY1570 TC527827 B. obtusa CBS119049 99 DQ458889 CRY1571 TC527828 B. obtusa CBS119049 99 DQ458889  下载: 导出CSV
下载: 导出CSV
-
[1] 张传清,徐志宏,孙品雷,等. 新病害:山核桃果实黑斑病病原菌的鉴定[J]. 植物保护,2010,36(4):160-162. ZHANG Chuanqing, XU Zhihong, SUN Pinlei, et al. Identification of the pathogen causing a new disease nut black spot on Carya cathayensis[J]. Plant Prot, 2010, 36(4):160-162. [2] 郑宏兵,束庆龙,丁之恩. 山核桃溃疡病与营养元素的关系[J]. 经济林研究,2007,25(3):19-23. ZHENG Hongbing, SHU Qinglong, DING Zhien. Relationship between nutrient elements in bark and canker of Carya cathayensis[J]. Nonwood For Res, 2007, 25(3):19-23. [3] ZHANG Chuanqing, XU Bingchao. First report of canker on Chinese hickory (Carya cathayensis) caused by Botryosphaeria dothidea in China[J]. Plant Dis, 2011, 95(10):1319. [4] 程燕林,梁军,吕全,等. 葡萄座腔菌科研究进展——鉴定,系统发育学和分子生态学[J]. 生态学报, 2011,31(11):3197-3207. CHEN Yanlin, LIANG Jun, LÜ Quan, et al. Advances in Botryosphaeriaceae:identification, phylogeny and molecular ecology[J]. Acta Ecol Sin, 2011, 31(11):3197-3207. [5] DAMM U, CROUS P W, FOURI P H. Botryosphaeriaceae as potential pathogens of Prunus species in South Africa, with descriptions of Diplodia africana and Lasiodiplodia plurivora sp. nov.[J]. Mycologia, 2007, 99(5):664-680. [6] SLIPPERS B, WINHFIELD M J, SMIT W A, et al. Taxonomy, phylogeny and identification of Botryosphaeriaceae associated with pome and stone fruit trees in South Africa and other regions of the world[J]. Plant Pathol, 2007, 56(1):128-139. [7] 缪承杜,洪葵. 真菌分类技术的研究进展[J]. 安徽农业科学,2007,35(22):6695-6697, 6700. MIAO Chengdu, HONG Kui. Research progress on technology of fungi classification[J]. J Anhui Agric Sci, 2007, 35(22):6695-6697, 6700. [8] 赵嘉平,梁军,吕全,等. 葡萄座腔菌属Botryosphaeria系统分类评述[J]. 林业科学研究,2007,20(3):415-422. ZHAO Jiaping, LIANG Jun, LÜ Quan, et al. Review on the systemic taxonomy of genus Botryosphaeria[J]. For Res, 2007, 20(3):415-422. [9] CHEN Shuaifei, PAVLI D, ROUX J, et al. Characterization of Botryosphaeriaceae from plantation-grown Eucalyptus species in south China[J]. Plant Pathol, 2011, 60(4):739-751. [10] PHILIPS A J L, CROUS P W, ALVES A. Diplodia seriata, the anamorph of "Botryosphaeria" obtuse[J]. Fung Div, 2007, 25:141-155. [11] ADESEMOVE A O, ESKALEN A. First report of Spencermartinsia viticola, Neofusicoccum australe, and N. parvum causing branch canker of citrus in California[J]. Plant Dis, 2011, 95(6):770. [12] 吴志辉,明文平. 山核桃溃疡病的化学防治研究[J]. 安徽农业科学,2006,34(14):3409-3412. WU Zhihui, MING Wenping. Experiment in the chemical control of Dothiorella gregaria canker of Carya[J]. J Anhui Agric Sci, 2006, 34(14):3409-3412. [13] 杨淑贞,丁立忠,楼君芳,等. 山核桃干腐病发生发展规律及防治技术[J]. 浙江林学院学报,2009,26(2):228-232. YANG Shuzheng, DING Lizhong, LOU Junfang, et al. Occurrence regularity of Carya cathayensis canker disease and its control[J]. J Zhejiang For Coll, 2009, 26(2):228-232. [14] 彭丽. 山核桃溃疡病的发生与防治[J]. 现代农业科技,2010,3(6):176-177. PENG Li. Occurrence of Carya cathayensis canker disease and its control[J]. Mod Agric Sci Technol, 2010, 3(6):176-177. [15] 张传清,章祖平,孙品雷,等. 山核桃干腐病菌对7种杀菌剂的敏感性比较及其对苯醚甲环唑敏感基线的建立[J]. 农药学学报,2011,13(1):84-86. ZHANG Chuanqing, ZHANG Zuping, SUN Pinlei, et al. Comparison of sensitivity of Botryosphaeria dothidea to 7 fungicides and its baseline sensitivity to difenoconazole[J]. Chin J Pest Sci, 2011, 13(1):84-86. [16] 田甜,沈振明,徐秋芳,等. 土壤中山核桃干腐病抑制菌的筛选和鉴定[J]. 浙江农林大学学报,2012,29(1):58-64. TIAN Tian, SHEN Zhenming, XU Qiufang, et al. Screening and identification of inhibiting strains for Carya cathayensis canker disease[J]. J Zhejiang A & F Univ, 2012, 29(1):58-64. [17] SLIPPERS B, CROUS P W, DENMAN S, et al. Combined multiple gene genealogies and phenotypic characters differentiate several species previously identified as Botryosphaeria dothidea[J]. Mycologia, 2004, 96(1):83-101. [18] SMITH H, KEMP G H J, WINGFIELD U J. Canker and die-back of Eucalyptus in South Africa caused by Botryosphaeria dothidea[J]. Plant Pathol, 1994, 43(6):1031-1034. [19] SMITH H, WINGFIELD M J, CROUS P W, et al. Sphaeropsis sapinea and Botryosplzaeria dothidea endophytic in Pinus spp. and Eucalyptus spp. in South Africa[J]. Afr Bot, 1996, 62(2):86-88. [20] 罗卿权,徐颖,刘美. 红瑞木溃疡病病原菌的鉴定[J]. 植物保护,2012,38(4):115-117. LUO Qingquan, XU Ying, LIU Mei. Identification of the pathogen causing trunk canker of Cornus alba[J]. Plant Prot, 2012, 38(4):115-117. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2014.02.012
 点击查看大图
点击查看大图

计量
- 文章访问数: 4904
- HTML全文浏览量: 790
- PDF下载量: 779
- 被引次数: 0
