




 下载:
下载:
-
中国是世界竹资源第一大国,竹子栽培和竹材利用历史悠久,在品种、面积、蓄积量、竹制品产量和出口额方面均居世界第一,素有“竹子王国”美誉。中国竹产业自20世纪90年代开始发展以来,其产品广泛应用于车辆、建筑、家具、装饰等各个领域[1-3]。中国竹家具的历史悠久,风格独特,造型美观,是传统家具的一个重要分支[4-5]。原竹家具是竹家具的重要组成部分,主要使用红竹Phyllostachys iridescins作为加工材料。在实际生产加工中,由于竹材含有较高的纤维素、半纤维素、淀粉、糖类及蛋白质等有机物,原竹家具在储存、加工和使用过程中容易产生虫蛀、霉变和开裂等问题,因此原竹家具用材需进行必要的改性处理[6-7]。竹材高温热处理是借鉴木材热处理技术而提出的改善竹材性能、提高产品质量的一种改性方法[8-11]。高温热处理有利于改善竹材材料的尺寸稳定性、耐腐性、耐候性等性能,实现竹材的高效利用,增加产品种类,提高产品质量[10]。目前,国内外对于竹材热处理的研究多集中于毛竹Phyllostachys edulis[3, 6-7, 12],对于红竹热处理研究较少,红竹分布于浙江、江苏、上海、四川和安徽等地,产量丰富,是具有代表性的小径级竹材,笔者对红竹竹材进行高温热处理试验,为原竹家具原料的改性处理和小径级竹的研究提供一定的技术支持。
-
试验材料为3年生红竹,采伐自安徽省宁国市。选取表面光滑,无明显缺陷的红竹,距地50 cm处采伐,往上1.5 m为试件原料,采伐12组,15根·组-1,共计180根。分别制作试件,参照GB/T 15780-1995《竹材物理力学性质试验方法》[13]和ISO 22157-1-2004 Specifies Test Methods for Evaluating the Following Characteristic Physical and Strength Properties for Bamboo[14]。制成10 mm × 10 mm × t mm(t为壁厚)竹片材试件,测基本密度、干缩率;20 mm × 20 mm × t mm竹片材试件,测顺纹抗压强度;160 mm × 10 mm × t mm竹片材试件,测抗弯强度;35 mm × 20 mm × t mm竹片材试件,测凸型顺纹抗剪强度;280 mm × 10 mm × t mm竹片材试件,测凹型顺纹抗拉强度;d mm(d为直径) × 40 mm的圆竹材试件,测顺纹抗压强度、顺纹抗剪前度、径向环刚度;d mm × 60 cm(测试点无竹节)圆竹材试件,测抗弯强度。
-
微机控制电子式木材万能试验机(DNS50),高低交变湿热试验箱(ER-10AGP),烘箱(SEG-021),电子分析天平(AB204-N),干燥碳化窑,游标卡尺,螺旋测微器等。
-
将试件放置于干燥箱中,初始温度为40 ℃,隔2 h温度升高5 ℃,直至103 ℃,竹材干燥至绝干;将绝干试件分别以温度为110,130,150,170 ℃,时间分别为1,2,3 h进行常压高温热处理。热处理过程中为防止竹材及圆竹材开裂,采用蒸汽高温热处理,试件放入干燥窑后,开始通蒸汽,进行高温热处理,试件共计12组。热处理后试件放入高低交变湿热试验箱中,以温度(20±2) ℃,湿度(65±5)%进行水分调节,待含水率稳定后,进行试验,竹材经过热处理后,其含水率处于稳定状态下为5.5%,因未处理竹材试验时含水率为12%,为了方便对比,将热处理竹材均计为含水率为12%时的强度。
-
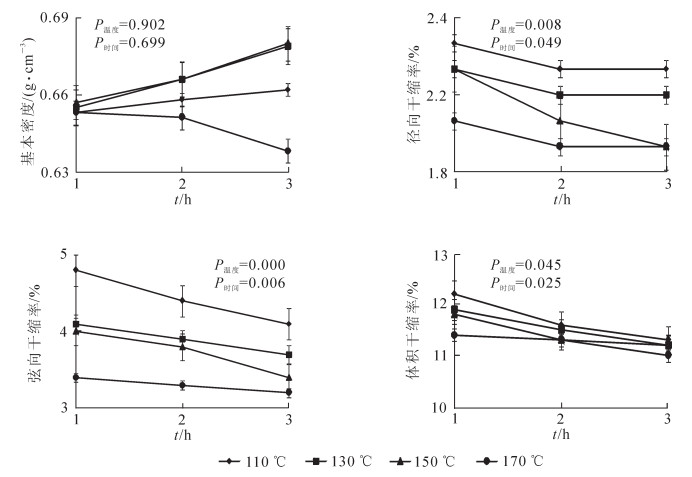
由图 1可见:热处理对竹材基本密度、径向干缩率、弦向干缩率、体积干缩率均有影响,根据各性能的方差分析,处理温度和时间对基本密度的影响不显著(P>0.05),对各项干缩性能的影响显著(P<0.05),其中温度为主影响因素;未处理竹材(对照)基本密度为0.60 g·cm-3,热处理竹材基本密度最小值为170 ℃,3 h时的0.64 g·cm-3,最大值为150 ℃,3 h时的0.68 g·cm-3,比对照增加了6.9%~13.9%;130和150 ℃竹材基本密度趋势差异较小,110~150 ℃呈现随温度升高、时间延长,基本密度增大趋势,170 ℃则呈相反趋势。径向干缩率、弦向干缩率、体积干缩率随热处理温度增加,数值下降;同一热处理温度下,时间延长,数值也有所下降;170 ℃时各项干缩率数值差异均较小;未处理竹材径向干缩率为2.2%,弦向干缩率为5.0%,体积干缩率为14.6%,热处理竹材径向干缩率最大为2.3%,数值最小为1.9%,比对未处理竹材变化率为4.55%~13.64%,弦向干缩率最大为4.8%,最小为3.2%,比对变化率为4.00%~36.00%,体积干缩率最大为12.2%,最小为11.0%,比对变化率16.44%~24.66%。

图 1 热处理竹材变化
Figure 1. Heat-treaded hamhoo's physical properties
-
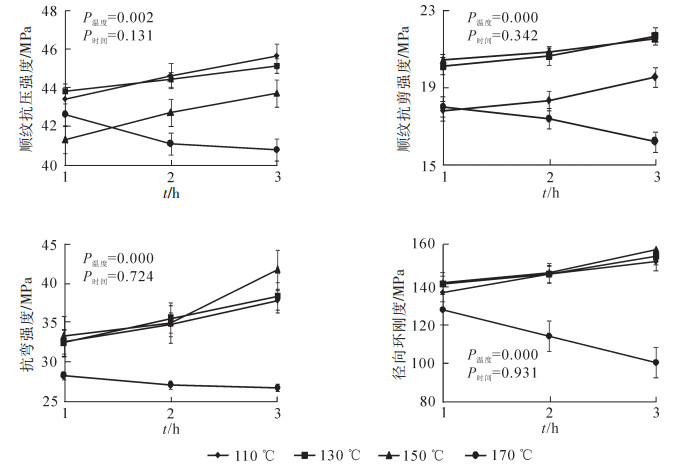
由图 2可见:热处理工艺对竹片材顺纹抗压强度、顺纹抗剪强度、抗弯强度、顺纹抗拉强度有明显的影响,竹片材各力学强度方差分析显示:处理温度为影响力学强度的极显著因素(P<0.01),处理时间影响不显著(P>0.05);力学强度随热处理温度升高整体呈现先上升后下降的趋势;未处理竹片材的抗剪强度、抗拉强度较170 ℃处理竹材高,这表明热处理温度过高,其力学性能反而有下降趋势;热处理温度为110,130,150 ℃时,随热处理时间的延长,力学强度呈增大趋势,而170 ℃则呈相反趋势。未处理竹片材顺纹抗压强度为51.6 MPa,热处理后最小值为69.3 MPa,最大值为72.4 MPa,比对变化率为32.17%~39.72%;未处理竹片材顺纹抗剪强度为19.6 MPa,热处理后最小值为17.5 MPa,最大值为24 MPa,比对变化率为-12.24%~20.91%;未处理竹片材的抗弯强度为142.7 MPa,热处理后最小值为162.3 MPa,最大值为196.6 MPa,比对变化率为13.52%~37.56%;未处理竹片材的顺纹抗拉强度为225.8 MPa,热处理后最小值为156.7 MPa,最大值为250.4 MPa,比对变化率为-30.73%~10.76%。

图 2 热处理工艺对竹片材力学性能影响
Figure 2. Heat-treaded bamboo sheet's mechanical properties
-
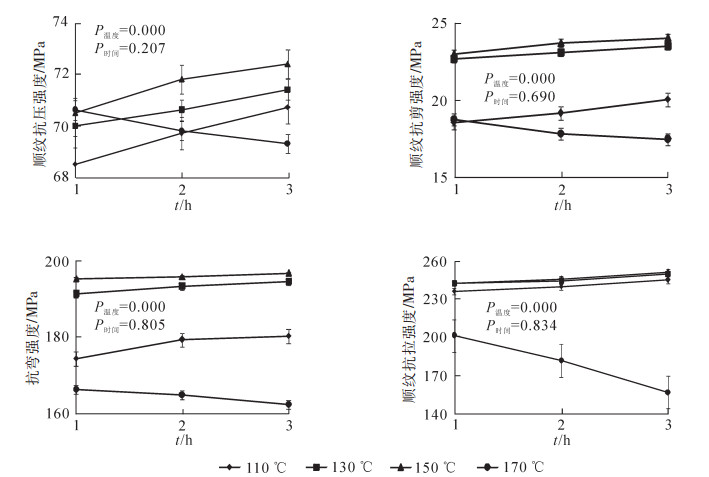
图 3反映了热处理工艺对圆竹材顺纹抗压强度、顺纹抗剪强度、抗弯强度、径向环刚度[15]的影响,圆竹材各力学性能方差分析可见,温度是影响各力学强度变化的主要因素(P<0.01),时间影响不显著(P>0.1);热处理后圆竹材的力学强度均大于未处理圆竹材;热处理温度为110,130,150 ℃时,圆竹材的力学强度随温度升高和时间延长呈上升趋势;110,130,150 ℃的温度条件时,圆竹顺纹抗压强度总体趋势从低到高依次为110,130,150 ℃,其中110 ℃与130,150 ℃差异较大;圆竹材顺纹抗剪强度增幅较缓,10,130,150 ℃温度之间抗剪强度差异较小;圆竹材抗弯强度3种温度条件差距不明显,时间因素影响较明显;圆竹材径向环刚度增幅较缓,110,130,150 ℃温度之间强度差异小;在170 ℃的温度条件下,圆竹材顺纹抗压强度、顺纹抗剪强度、抗弯强度及径向环刚度等4个力学性能均呈随时间延长,强度下降的趋势。未处理圆竹材顺纹抗压强度为29.8 MPa,热处理后最小值为40.8 MPa,最大值45.6 MPa,比对变化率为35.90%~52.01%;未处理圆竹材顺纹抗剪强度为11.1 MPa,热处理后最小值为16.2 MPa,最大值为21.6 MPa,比对变化率为43.24%~90.99%;未处理竹圆竹材抗弯强度为18.6 MPa,热处理后最小值为26.8 MPa,最大值为41.7 MPa,比对变化率为42.47%~122.58%;未处理圆竹材径向环刚度为98 kPa,热处理后最小值为100.4 kPa,最大值为159.8 kPa,比对变化率为2.14%~52.55%。可以看出,热处理后圆竹材的力学强度提升较大,这对于制作原竹家具是适合的改性方式。

图 3 热处理工艺对圆竹材力学性能的影响
Figure 3. Heat-treaded round bamboo's mechanical properties
-
根据木材学理论及木材碳化机制[16],竹材在热处理过程中纤维素,尤其是半纤维素发生分解,含量降低,同时竹材内部结构发生变化,导致氢键浓度降低,或氢键被非亲水性基团所取代,引起竹材干缩;在温度为110~150 ℃时,温度相对较低,竹材内部的纤维素、半纤维素分解缓慢,竹材内部自由水蒸发较多,体积干缩较大,竹材基本密度相对增加,力学性能增加;随着热处理温度的升高,竹材的胞壁物质、半纤维素、纤维素分解剧烈,竹材的胞壁物质减少,而体积基本不变,竹材基本密度相对降低,其力学性能降低。
由以上分析可知,温度是影响热处理后竹材物理力学性能的显著因素;110 ℃和170 ℃时的竹材力学性能较弱,而130 ℃与150 ℃的竹材其各项力学性能均相似,因此,首先排除110 ℃和170 ℃作为生产中热处理工艺温度;实际生产和热处理过程中,安全、环保、成本等均是需要综合考虑的因素,150 ℃热处理温度,其消耗燃料、有害气体排放均较多,温度较高,安全隐患较大。因此,最适宜生产的热处理工艺为温度130 ℃,时间2 h。
Physical and mechanical properties of Phyllostachys iridescins under normal pressure and heat temperature
-
摘要: 以红竹Phyllostachys iridescins为研究对象,研究了常压高温热处理温度(110,130,150,170℃)与处理时间(1,2,3 h)对红竹竹材物理力学性能的影响。结果表明:热处理后红竹材的物理力学性能优于未处理竹材,热处理温度是影响竹材性能的主要因素;在110,130,150,170℃热处理温度下,圆竹材顺纹抗压强度增长率为35.90%~52.01%,圆竹材顺纹抗剪强度增长率43.24%~90.99%,圆竹材抗弯强度增长率42.47%~122.58%,圆竹材径向环刚度增长率2.14%~52.55%;170℃热处理竹材的各项干缩性能较好;130℃和150℃热处理竹材的力学性能相近。综合各因素,适宜原竹家具用材红竹竹材的热处理工艺为温度130℃,时间2 h。Abstract: The effects of treatment temperature (110, 130, 150, 170℃) and time (1, 2, 3 h) on the physical and mechanical properties of Phyllostachys iridescins (including bamboo strips and round bamboo strips) were studied. The results showed that the physical and mechanical properties of Ph. iridescins after heat treatment were better than untreated bamboo, and the heat treatment temperature was the main factor affecting bamboo properties. Treated by 110, 130, 150, 170℃, compressive growth rate of round bamboo strips was 35.90%-52.01%, its shear growth rate was 43.24%-90.99%, its bending growth rate was 42.47%-122.58%, and its radial ring Stiffness growth rate was 2.14%-52.55%. The shrinkage of bamboo strip was better at 170℃. 130℃ and 150℃ heat treated bamboo's mechanical properties were similar. Considering all the factors, the heat treatment process of the bamboo material was 130℃ for 2 h.
-
[1] 李延军, 许斌, 张齐生, 等.我国竹材加工产业现状与对策分析[J].林业工程学报, 2016, 1(1):2-7. LI Yanjun, XU Bin, ZHANG Qisheng, et al. Present situation and the countermeasure analysis of bamboo timber processing industry in China[J]. J For Eng, 2016, 1(1):2-7. [2] 闫薇, 傅万四, 张彬, 等.基于规格竹片的胶合界面研究现状及建议[J].林业工程学报, 2016, 1(5):20-25. YAN Wei, FU Wansi, ZHANG Bin, et al. Research status and suggestion on specification bamboo bonding interface[J]. J For Eng, 2016, 1(5):20-25. [3] 黄梦雪, 张晓春, 余文军, 等.高温蒸汽软化竹材的力学性能及结构表征[J].林业工程学报, 2016, 1(4):64-68. HUANG Mengxue, ZHANG Xiaochun, YU Wenjun, et al. Mechanical properties and structure characterization of bamboo softened by high temperature steam[J]. J For Eng, 2016, 1(4):64-68. [4] 曾利.竹藤家具及其保养与修复技术[J].林产工业, 2010, 37(1):40-42. ZENG Li. The conservation and restoration of rattan furniture[J]. China For Prod Ind, 2010, 37(1):40-42. [5] 何晓琴.中国传统竹家具的文化特征[J].世界竹藤通讯, 2006, 4(2):42-45. HE Xiaoqin. Culture characteristics of traditional bamboo furniture in China[J]. World Bamboo Rat, 2006, 4(2):42-45. [6] 包永洁, 蒋身学, 程大莉, 等.热处理对竹材物理力学性能的影响[J].竹子研究汇刊, 2009, 28(4):50-53. BAO Yongjie, JAING Shenxue, CHENG Dali, et al. The effects of heat treatment on physical-mechanical properties of bamboo[J]. J Bamboo Res, 2009, 28(4):50-53. [7] 林勇, 沈珏程, 于利, 等.高温热处理竹材的物理力学性能研究[J].林业机械与木工设备, 2012, 40(8):22-24. LIN Yong, SHEN Yucheng, YU Li, et al. Study of physical-mechanical properties of bamboos through high temperature heat treatment[J]. For Mach Woodwork Equip, 2012, 40(8):22-24. [8] 李涛, 顾炼百. 185℃高温热处理对水曲柳木材力学性能的影响[J].林业科学, 2009, 45(2):92-97. LI Tao, GU Lianbai. Effects of high temperature heat treatment at 185℃ on mechanical properties of ash wood[J]. Sci Silv Sin, 2009, 45(2):92-97. [9] 李延军, 唐荣强, 鲍滨福, 等.高温热处理杉木力学性能与尺寸稳定性研究[J].北京林业大学学报, 2010, 32(4):232-235. LI Yanjun, TANG Rongqiang, BAO Binfu, et al. Mechanical properties and dimensional stability of heat-treated Chinese fir[J]. J Beijing For Univ, 2010, 32(4):232-235. [10] 李延军, 孙会, 鲍滨福, 等.国内外木材热处理技术研究进展及展望[J].浙江林业科技, 2008, 28(5):75-79. LI Yanjun, SUN Hui, BAO Binfu, et al. Advances and prospect of research on technology of wood heat-treatment[J]. J Zhejiang For Sci Technol, 2008, 28(5):75-79. [11] 李贤军, 傅峰, 蔡智勇, 等.高温热处理对木材吸湿性和尺寸稳定性的影响[J].中南林业科技大学学报, 2010, 30(6):92-96. LI Xianjun, FU Feng, CAI Zhiyong, et al. The effect of high temperature thermal treatment on moisture absorption and dimension stability of wood[J]. J Cent South Univ For Technol, 2010, 30(6):92-96. [12] 侯瑞光, 刘元, 李贤军, 等.高温热处理对重组竹物理力学性能的影响[J].中南林业科技大学学报, 2013, 33(2):101-104. HOU Ruiguang, LIU Yuan, LI Xianjun, et al. Effects of heat treatment on physical-mechanical properties of reconstituted bamboo lumber (RBL)[J]. J Cent South Univ For Technol, 2013, 33(2):101-104. [13] 国家技术监督局. GB/T 15780-1995竹材物理力学性质试验方法[S]. 北京: 中国标准出版社, 1996. [14] ISO/TC 165 Timber Structures. ISO 22157-1-2004 Specifies Test Methods for Evaluating the Following Characteristic Physical and Strength Properties for Bamboo[S]. https: //www. iso. org/standard/36150. html. [15] 张文福, 江泽慧, 王戈, 等.用环刚度法评价圆竹径向抗压力学性能[J].北京林业大学学报, 2013, 35(1):119-122. ZHANG Wenfu, JIANG Zehui, WANG Ge, et al. Radial compression mechanical properties of bamboo-culm by ring stiffness[J]. J Beijing For Univ, 2013, 35(1):119-122. [16] 吴帅, 于志明.木材炭化技术的发展趋势[J].中国人造板, 2008, 15(5):3-6. WU Shuai, YU Zhiming. Review on current situation and developing trend of wood carbonization technology[J]. China Wood-Based Pannels, 2008, 15(5):3-6. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2018.04.023
 点击查看大图
点击查看大图
图(3)
计量
- 文章访问数: 3827
- HTML全文浏览量: 1275
- PDF下载量: 573
- 被引次数: 0
