




 下载:
下载:
-
竹材是重要的速生可再生材料,具有强度高、硬度大、韧性好、纹理美观、生态功能强等优良特性,但也存在易发生虫蛀和霉腐等缺陷,严重限制了其在室内外的应用。热处理作为一种绿色环保的纯物理改性处理方法,能够显著改善材料的尺寸稳定性和耐候性,在板材预处理过程中优势显著[1-4]。竹材热处理是当前竹材高值化利用的重要方向,根据传热介质差异,通常有蒸气、空气和导热油热处理等3种方式。蒸气介质热处理法以饱和水蒸气为导热介质,多在炭化罐中进行,处理后竹材的密度及相应的总体力学性能下降,吸水厚度膨胀率降低,尺寸稳定性显著提升[5-10]。空气介质热处理法以空气为导热介质,多在高温热处理箱中进行,处理后竹材的密度、总体力学性能下降,尺寸稳定性大幅改善[11-14]。油介质热处理以各类导热油为介质,相较于前两者,处理后试材密度较高,抗弯强度降低,润湿性能减弱,耐久效果提升[15]。总的来说,水蒸气热处理过程对设备及操作的要求极高,热处理周期长、能耗大、设备投资高、劳动生产率较低,热处理后竹片的强度衰减较大;空气热处理法对处理温度精度的要求较高;油介质法使木材受热更均匀,控温更准确,但沥干过程所需时间较长,热油在使用寿命终结后亦存在收集和处理问题[16]。鉴于此,本研究选取4~6年生毛竹Phyllostachys edulis材为原料,利用热压机的高温和热压特性,研究不同热处理温度(225,250,275,300,325,350和375 ℃)下,毛竹材物理力学性能的变化。旨在研发一种接触式高温快速热压处理技术,在保障热处理性能的同时,缩短热处理时间,降低生产成本,为快速热处理工艺提供理论依据和基础数据。
-
4~6年生毛竹产自浙江省湖州市安吉县。原竹经横向截断、纵向裂解、四面粗刨、四面精刨、去青去黄等工艺,加工成800 mm(纵向)× 25 mm(弦向)× 6 mm尺寸试片,干燥至含水率为8%,用于后期热处理试验。
-
毛竹试片对半截开(400 mm × 25 mm × 6 mm),近青面朝上摆放,上下表面均覆盖锡箔纸,置于热压机中热处理。固定处理时间为45 s,固定压力为0.15 MPa,设置热处理温度为225(T1),250(T2),275(T3),300(T4),325(T5),350(T6)和375 ℃(T7)。处理结束后取出样件,自然冷却至室温;置于温度为(20±2)℃,相对湿度为(65±5)%的调温调湿箱至质量恒定。以相同尺寸未经热处理的竹片作为对照,对照(ck)也进行相同的调温调湿处理。重复3次·处理-1。
-
参照国家标准GB/T 15780-1995《竹材物理力学性质试验方法》测热处理前后竹材的物理力学性能。
-
试片尺寸为20 mm × 20 mm × 6 mm,每处理取9块。调温调湿,称量后置于(103±2)℃烘箱内烘至全干,干燥冷却后再次称量,经计算获得平衡含水率。平衡含水率W=(m1-m0)/m0×100%。其中:m1(g)为调温调湿后试片质量,m0(g)为试片全干时质量。
-
试片为饱和水分的湿材,尺寸为10 mm × 10 mm × 6 mm,取9块·处理-1。用百分表测量装置测量湿材径向和弦向的尺寸,计算体积;置于(20±2)℃、相对湿度(65±5)%的恒温恒湿箱中至气干,测量气干时径向、弦向的尺寸并计算体积;最后放入烘箱烘至全干,测量全干时径向、弦向尺寸并计算体积。经计算得到径向和弦向的全干缩率、气干干缩率及体积的全干缩率和体积的气干干缩率。径向或弦向全干缩率Bmax=(Lmax-L0)/Lmax×100%。其中:Lmax(mm)为湿材试片径向或弦向靠竹青、竹黄处尺寸的均值,L0(mm)为试片全干时径向或弦向靠竹青、竹黄处尺寸的均值。径向或弦向气干干缩率BW=(Lmax-Lw)/Lmax×100%。其中:Lw(mm)为试片气干时径向或弦向靠竹青、竹黄处尺寸的均值。体积全干缩率βVmax=(Vmax-V0)/Vmax×100%。其中:Vmax(mm3)为试片湿材时的体积,V0(mm3)为试片全干时的体积。体积气干干缩率βVw=(Vmax-Vw)/Vmax。其中:Vw(mm3)为试片气干时的体积。
-
试片尺寸为10 mm × 10 mm × 6 mm,调温调湿后测量试片尺寸并计算体积,称取质量,经计算获得气干密度。气干密度ρ=m1/V。其中:V(mm3)为试片调温调湿后体积。
-
试片尺寸为20 mm × 20 mm × 6 mm,每处理取9块。用百分表测量竹壁厚度和弦向宽度,万能力学试验机以均匀速度加荷,在(1.0 ± 0.5)min内破坏试样,记录破坏载荷;经计算得到顺纹抗压强度。顺纹抗压强度σ=Pmax/bt。其中:Pmax(N)为破坏载荷,b(mm)为试片宽度,t(mm)为试片厚度,即竹壁厚度。
-
试片尺寸为160 mm × 10 mm × 6 mm,每处理取6块。测量竹壁厚度和弦向高度,万能力学试验机试片弦向以均匀速度加荷,在(1 ± 0.5)min内破坏试片,记录破坏荷载、抗弯强度、抗弯弹性模量及挠度。
-
各组数据经整理后取平均值,在α=0.05水平进行显著性分析,而后绘制图表。
-
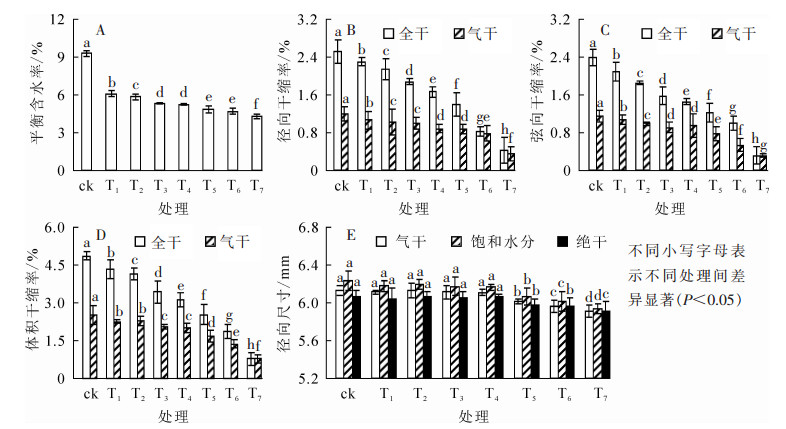
由图 1A可知:竹材的平衡含水率随温度的升高明显下降(P<0.05)。热处理温度为375 ℃时,平衡含水率达到最低值4.27%,相比对照下降了53.95%。热处理对平衡含水率影响显著。

图 1 热处理温度对平衡含水率、干缩率和径向尺寸变化的影响
Figure 1. Effect of heat treatment temperature on equilibrium moisture content, shrinkage rate and radial dimensional changes
竹材各项干缩性均随温度的上升而下降,温度对干缩率影响显著(P<0.05)。由图 1B可知:竹材径向全干干缩率和气干干缩率均随温度的上升而下降,当处理温度从225 ℃上升至375 ℃时,径向全干干缩率和径向气干干缩率分别下降了83.60%和70.59%。弦向全干干缩率从2.38%减少至0.31%,弦向气干干缩率从1.14%减少至0.30%(图 1C);体积全干干缩率从4.86%减少至0.78%,体积气干干缩率从2.51%减少至0.66%(图 1D)。相同热处理温度下,弦向干缩率的变化率>体积干缩率的变化率>径向干缩率的变化率,与左宋林等[17]研究结果一致。由图 1E可知:较对照而言,不同处理温度对径向尺寸影响不同。当热处理温度不高于300 ℃时,气干状态下径向尺寸变化不明显;当温度升至325 ℃后,由于含水率的降低,径向尺寸开始缩减,至375 ℃时,气干状态下处理组径向尺寸相比对照缩减了3.11%。
半纤维素和纤维素是竹材的主要成分。由于分子量低,高温下竹材中的半纤维素最先开始热解[18],而后是纤维素。半纤维素是一种无定性物质,由2种或多种糖基组成,其结构具有支化度,主链和侧链上含有亲水性基团,吸湿性大,是使竹木材产生吸湿膨胀、变形开裂的因素之一[19]。热处理导致半纤维素中的某些多糖裂解,羟基基团变得十分活跃,在氧气、水分和热量的作用下,裂解生成糖醛和某些糖类;游离羟基的减少[7]和能聚合生成不溶于水的聚合物的新物质的生成,降低了木质材料的吸湿性,使得竹材的平衡含水率大幅降低,也减少了竹材的干缩和湿胀[20-23],从而提升了尺寸稳定性。此外,在热处理过程中,竹材的水分逐渐失去,细胞壁非结晶区的纤维素分子链间距减少,游离羟基相互之间分子作用力增强,形成氢键总数增加,从而使非晶区纤维素分子的取向增强,这也是热处理材的尺寸稳定性得到改善的原因之一[24]。
-
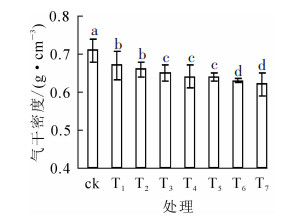
热处理温度对竹材气干密度影响显著(P<0.05)。由图 2可知:随着热处理温度升高,竹材气干密度逐渐降低,且温度越高,降低的幅度越大;当温度为375 ℃时,气干密度达到了最低值(0.62 g·cm-3),相比对照降低了13.04%,与黄成建[13]研究结果一致。包永洁[7]研究认为:当温度高于180 ℃时,半纤维素急剧分解,纤维素相对含量也减少,即竹材胞壁物质含量减少,密度也相应减小。

图 2 热处理温度对竹材气干密度的影响
Figure 2. Effect of heat treatment temperature on density
-
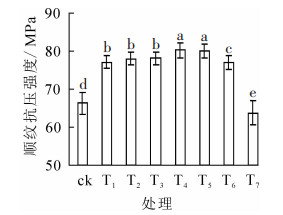
竹材顺纹抗压强度随温度的变化呈先上升后下降趋势(图 3)。当温度不高于300 ℃时,顺纹抗压强度随温度上升而增强,且增幅不断加大;300 ℃时,顺纹抗压强度达最大(80.19 MPa),相比对照增加了21.08%;当温度继续上升,顺纹抗压强度增幅减小,375 ℃时同对照相比降低了3.70%。

图 3 热处理温度对竹材顺纹抗压强度的影响
Figure 3. Effect of heat treatment temperature on compression strength
热处理使得竹片密度及含水率均降低,从而降低强度,提高竹材刚性[25];随着含水率从纤维饱和点下降至零点,除抗拉强度外,其他强度都显著增加[26-27]。在竹材的不同力学性能指标中,顺纹抗压强度受含水率的影响最大,而密度是决定竹材力学性能对含水率变化敏感程度的重要因素,密度越小,敏感性越高。本研究发现,处理温度不高于375 ℃时,竹材顺纹抗压强度表现为增长;一方面是由于低含水率提高了竹材强度,随着密度的降低,力学性能对含水率变化敏感程度逐渐增高,另一方面竹材表面在高温下形成了壳状层,抗弯弹性模量和抗压强度得到改善[5],从而弥补了因密度下降带来的强度的降低。温度达到375 ℃后,高温加速了裂解作用,低含水率带来的增强作用无法弥补密度降低引起的强度降低,因此顺纹抗压强度最终出现了下降。
包永洁[7]采用水蒸气介质热处理法对竹材进行热处理,发现处理温度对顺纹抗压强度影响显著,从20 ℃到210 ℃,顺纹抗压强度下降20.67%。采用空气介质热处理法对毛竹材进行热处理,处理温度为160~200 ℃时,毛竹顺纹抗压强度降低1.30%~19.00%[11-14]。相比之下,高温快速热压处理法热处理温度更高(375℃),顺纹抗压强度下降程度更小(平均降低3.70%),因此优势更明显。
-
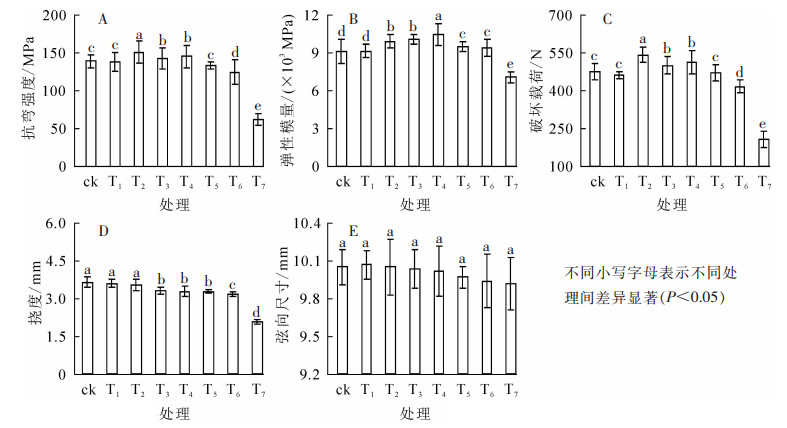
由图 4A可知:热处理温度为225 ℃时,抗弯强度(MOR)与对照相比,变化不大。当温度升至250 ℃时,处理组抗弯强度增幅最大,增加了8.37%,而后增幅逐渐减小。至325 ℃时,处理组抗弯强度开始降低,平均下降4.34%,而后随温度升高,迅速下降。375 ℃时,处理材抗弯强度达最低值,仅为61.85 MPa,相比对照降低了55.61%。

图 4 热处理温度对竹材抗弯强度、抗弯弹性模量、破坏载荷、挠度及弦向尺寸变化的影响
Figure 4. Effect of heat treatment temperature on MOR, MOE, failure load, deflection and tangential dimensional changes
毛竹片的抗弯弹性模量(MOE)的变化以300 ℃为转折点(图 4B),当热处理温度不高于300 ℃时,抗弯弹性模量随处理温度的升高而上升,至300 ℃时达最大值(10 487.44 MPa),较对照增长15.30%。处理温度高于300 ℃后,抗弯弹性模量仍然保持上升趋势,但增幅减小;375 ℃时达最低值(7 071.14 MPa),比对照降低了22.26%。
破坏载荷的变化趋势与抗弯强度基本一致(图 4C)。225 ℃下处理组破坏载荷变化不大,比对照仅降低了2.72%。250 ℃时处理组破坏载荷达到最大值543.81 N,相比对照上升了14.02%,而后逐渐减小,至325 ℃时低于对照组。后随温度升高迅速下降,至375 ℃时下降了56.56%,达最低值(207.17 N)。挠度随温度升高呈下降趋势(图 4D)。225 ℃时处理组挠度为3.61 mm,与对照相比下降了1.29%;至375 ℃达最低值(2.08 mm),相比对照组下降了43.18%。
温度对弦向尺寸变化的影响不显著(图 4E,P>0.05)。当热处理温度不高于300 ℃时,弦向尺寸受热压处理变化很小,与对照无显著差异;自325 ℃至375 ℃,随温度升高,竹材弦向收缩加剧,与对照相比,处理组弦向尺寸缩减了0.75%~1.25%。因此,弦向尺寸变化不是竹材抗弯强度及抗弯弹性模量变化的主要影响因素。
结合图 1A可知:随温度升高竹材平衡含水率逐渐降低,竹材所能承受的破坏载荷增大,抗弯强度和抗弯弹性模量增强。当热处理温度上升至325 ℃后,挠度相比300 ℃时未发生明显变化,但由于分解速率逐渐增大,竹材力学性受到负面影响,破坏载荷减小,抗弯强度和抗弯弹性模量出现降低。由此看出,225~300 ℃条件下高温快速热压处理法能增大竹材所能承受的破坏载荷,提高竹材抗弯强度,375 ℃之前,高温快速热压法能提升竹材抗弯弹性模量。相比于其他热处理法“抗弯强度随热处理温度升高、时间的延长呈明显下降趋势,热处理温度范围狭窄,降幅大”[5-15]的不足,高温快速热压处理法可达到更高温度,且处理后的竹材能承受更大的破坏载荷,具有更好的抗弯强度和抗弯弹性模量。
-
随着热处理温度的升高,竹材的平衡含水率和气干密度均呈下降趋势,最高分别下降了53.95%和13.04%。相同处理条件下,弦向干缩率的变化率>体积干缩率的变化率>径向干缩率的变化率,竹材的尺寸稳定性得到显著改善。
高温快速热压处理法处理的毛竹材力学性能得到增强。挠度随温度升高逐渐减小,顺纹抗压强度、破坏载荷、抗弯强度和抗弯弹性模量均随温度升高呈先上升后下降趋势。挠度至375 ℃达最低值(2.08 mm),相比对照下降了43.18%;顺纹抗压强度在300 ℃时最高,相比对照提升了21.08%;破坏载荷在250 ℃时达最大值543.81 N,比对照上升了14.02%;抗弯强度在250 ℃时增幅最大,比对照增加了8.37%;抗弯弹性模量在300 ℃时最高,达10 148.69 MPa,较对照增长了15.30%。
相比于其他热处理法,高温快速热压处理法处理后竹材顺纹抗压强度更强,抗弯强度和静曲模量更好,尺寸稳定性也更高。
Physical and mechanical properties of Phyllostachys edulis with fast hot-pressing and high temperature
-
摘要: 选取4~6年生的毛竹Phyllostachys edulis材为原料,采用热压机对毛竹材进行高温快速热压处理,研究不同热处理温度(225,250,275,300,325,350和375℃)下竹材物理力学性能的变化。结果表明:随着热处理温度的升高,竹材平衡含水率和气干密度明显下降(P < 0.05),与未处理材相比分别降低了34.39%~53.95%和7.89%~13.04%。相同热处理温度下,弦向干缩率的变化率>体积干缩率的变化率>径向干缩率的变化率;当温度达到375℃时,弦向全干干缩率下降了86.81%,径向全干干缩率下降了83.60%,体积全干干缩率下降了83.95%,达各向的最大值。热处理温度升高,竹材顺纹抗压强度、抗弯强度和抗弯弹性模量均先增加后减少,其中,顺纹抗压强度在375℃时达最小值(63.78 MPa);抗弯强度在250℃时达最大值(151.00 MPa),在375℃条件下达最小值(61.85 MPa);抗弯弹性模量在300℃时达最大值(10 487.44 MPa),在375℃时达最小值(7 071.14 MPa)。认为竹材接触式快速热处理工艺提升了竹材尺寸稳定性和力学性能。Abstract: To improve dimensional stability and weather resistance of a material, heat treatment, with fast hot-pressing and high temperature, was used. Pieces of 4-6 year-old Phyllostachys edulis were selected. Samples were treated with different temperatures (225, 250, 275, 300, 325, 350, and 375℃). Physical and mechanical properties were tested before and after treatments. Results showed that with an increase of temperature, there was an obvious decrease in equilibrium moisture content (34.39%-53.95%) and bamboo density (7.89%-13.04%) both P < 0.05. With the same conditions, the following occurred:rate of change for tangential shrinkage rate > rate of change for volume shrinkage rate > rate of change for radial shrinkage rate. When the temperature reached 375℃, tangential oven-dry shrinkage rate decreased 86.81%; whereas, radial oven-dry shrinkage rate dropped 83.60%, and volume oven-dry shrinkage rate decreased 83.95%, reaching a maximum in all directions. Compression strength parallel to the grain, modulus of rupture (MOR), and modulus of elasticity (MOE) of the bamboo increased with temperature first and then decreased. Compression strength reached a minimum of 63.78 MPa at 375℃. MOR reached a peak at 250℃ with 151.00 MPa, and decreased to 61.85 MPa at 375℃. MOE performed best at 300℃ with 10 487.44 MPa, and worst at 375℃ with 7 071.14 MPa. Thus, this experiment proved that fast hot-pressing with high temperature can improve the dimensional stability of the bamboo, the mechanical properties also generally improved after treatments.
-
图 1 热处理温度对平衡含水率、干缩率和径向尺寸变化的影响
Figure 1 Effect of heat treatment temperature on equilibrium moisture content, shrinkage rate and radial dimensional changes
图 2 热处理温度对竹材气干密度的影响
不同小写字母表示不同处理间差异显著(P<0.05)
Figure 2 Effect of heat treatment temperature on density
图 3 热处理温度对竹材顺纹抗压强度的影响
不同小写字母表示不同处理间差异显著(P<0.05)
Figure 3 Effect of heat treatment temperature on compression strength
-
[1] 夏雨, 牛帅红, 李延军, 等.常压高温热处理对红竹竹材物理力学性能的影响[J].浙江农林大学学报, 2018, 35(4):765-770. XIA Yu, NIU Shuaihong, LI Yanjun, et al. Physical and mechanical properties of Phyllostachys iridescins under normal pressure and heat temperature[J]. J Zhejiang A & F Univ, 2018, 35(4):765-770. [2] 汤颖, 李君彪, 沈钰程, 等.热处理工艺对竹材性能的影响[J].浙江农林大学学报, 2014, 31(2):167-171. TANG Ying, LI Junbiao, SHEN Yucheng, et al. Phyllostachys edulis with high temperature heat treatments[J]. J Zhejiang A & F Univ, 2014, 31(2):167-171. [3] 张亚梅, 于文吉.热处理对竹材性能及变色的影响[J].木材加工机械, 2008(4):40-43, 47. ZHANG Yamei, YU Wenji. Research about heat-treatment effect on bamboo's performance and discoloration[J]. Wood Proc Mach, 2008(4):40-43, 47. [4] 秦莉, 于文吉, 余养伦.重组竹材耐腐防霉性能的研究[J].木材工业, 2010, 24(4):9-11. QIN Li, YU Wenji, YU Yanglun. Mold and decay resistance of reconstituted bamboo lumber[J]. China Wood Ind, 24(4):9-11. [5] 邵卓平, 周学辉, 魏涛, 等.竹材在不同介质中加热处理后的强度变异[J].林产工业, 2003, 30(3):26-29. SHAO Zhuoping, ZHOU Xuehui, WEI Tao, et al. The strength variance of bamboo blanks heat-treated with different methods[J]. China For Prod Ind, 30(3):26-29. [6] 蒋身学, 程大莉, 张晓春, 等.高温热处理竹材重组材工艺及性能[J].林业科技开发, 2008, 22(6):80-82. JIANG Shenxue, CHENG Dali, ZHANG Xiaochun, et al. Pilot study on process and properties of high temperature heat-treated reconstituted bamboo lumber[J]. China For Sci Technol, 2008, 22(6):80-82. [7] 包永洁.高温热处理前后竹材主要化学成分及物理力学性能研究[D].南京: 南京林业大学, 2009. BAO Yongjie. Research on the Main Chemical Component, Physical and Mechanical Properties of Bamboo after Thermal Treatment[D]. Nanjing: Nanjing Forestry University, 2009. [8] 包永洁, 蒋身学, 程大莉, 等.热处理对竹材物理力学性能的影响[J].竹子研究汇刊, 2009, 28(4):50-53. BAO Yongjie, JIANG Shenxue, CHENG Dali, et al. The effects of heat treatment on physical-mechanical properties of bamboo[J]. J Bamboo Res, 2009, 28(4):50-53. [9] 秦莉.热处理对重组竹材物理力学及耐久性能影响的研究[D].北京: 中国林业科学研究院, 2010. QIN Li. Effect of Thermo-Treatment on Physical, Mechanical Properties and Durability of Reconstituted Bamboo Lumber[D]. Beijing: Chinese Academy of Forestry, 2010. [10] 宋路路, 任慧群, 王新洲, 等.高温饱和蒸汽处理对竹材材性的影响[J].林业工程学报, 2018, 3(2):23-28. SONG Lulu, REN Huiqun, WANG Xinzhou, et al. Effect of high temperature saturated steam treatment on bamboo properties[J]. J For Eng, 2018, 3(2):23-28. [11] 林勇, 沈钰程, 于利, 等.高温热处理竹材的物理力学性能研究[J].林业机械与木工设备, 2012, 40(8):22-24. LIN Yong, SHEN Yucheng, YU Li, et al. Study of physical-mechanical properties of bamboo through high temperature heat treatment[J]. For Mach Woodwork Equip, 2012, 40(8):22-24. [12] 程大莉, 蒋身学.热处理对慈竹重组材性能的影响[J].林业工程学报, 2012, 26(4):73-75. CHENG Dali, JIANG Shenxue. Effect of heat treatment on properties of Neosinocalamus affinis reconstituted bamboo lumber[J]. China For Sci Technol, 2012, 26(4):73-75. [13] 黄成建.热处理毛竹材细胞壁结构及力学性能研究[D].杭州: 浙江农林大学, 2015. HUANG Chengjian. Microstructural and Micromechanical Properties of Thermo-treated Bamboo Cell Wall[D]. Hangzhou: Zhejiang A&F University, 2015. [14] 刘炀, 曹琳, 李金朋.中低温热处理对竹材材性的影响[J].河北林业科技, 2016(3):14-17. LIU Yang, CAO Lin, LI Jinpeng. Effect of heat treatment in middle and low temperature on the quarantine of bamboo[J]. J Hebei For Sci Technol, 2016(3):14-17. [15] 赵鹤, 卢康平, 林金国.油介质热处理对毛竹材性能的影响[J].林业机械与木工设备, 2015, 43(12):14-16. ZHAO He, LU Kangping, LIN Jinguo. Effect on properties of Phyllostachys heterocycla cv. Pubescens by heat treatment with oil medium[J]. For Mach Woodwork Equip, 2015, 43(12):14-16. [16] 丁涛, 蔡家斌, 耿君.欧洲木材热处理技术的研究及应用[J].木材工业, 2015, 29(5):29-33. DING Tao, CAI Jiabin, GENG Jun. Research and application of wood thermal modification technologies in Europe[J]. China Wood Ind, 2015, 29(5):29-33. [17] 左宋林, 高尚愚, 阮锡根, 等.炭化过程中的竹材收缩率[J].南京林业大学学报(自然科学版), 2003, 27(3):15-20. ZUO Songlin, GAO Shangyu, RUAN Xigen, et al. A study on shrinkages during the carbonization of bamboo[J]. J Nanjing For Univ Nat Sci Ed, 2003, 27(3):15-20. [18] FENGEL D, WEGENER G. Wood:Chemistry, Ultrastructure, Reactions[M]. Verlag Kessel, Germany:Remagen-Oberwinter, 2003. [19] 张亚梅.热处理对竹材颜色及物理力学性能影响的研究[D].北京: 中国林业科学研究院, 2010. ZHANG Yamei. Study on the Effect of Color and Physical-Mechanical Properties for Heat-Treated Bamboo[D]. Beijing: Chinese Academy of Forestry, 2010. [20] NUOPPONEN M, VUORINEN T, JÄMSÄ S, et al. The effects of a heat treatment on the behaviour of extractives in softwood studied by FTIR spectroscopic methods[J]. Wood Sci Technol, 2003, 37(2):109-115. [21] 吴帅, 于志明.木材炭化技术的发展趋势[J].中国人造板, 2008, 15(5):3-6. WU Shuai, YU Zhiming. Review on current situation and developing trend of wood carbonization technology[J]. China Woodbased Panels, 2008, 15(5):3-6. [22] SANTOS J A. Mechanical behaviour of Eucalyptus wood modified by heat[J]. Wood Sci Technol, 2000, 34(1):39-43. [23] 孙润鹤, 刘元, 李贤军, 等.高温热处理对竹束颜色和平衡含水率的影响[J].中南林业科技大学学报, 2012, 32(9):138-141. SUN Runhe, LIU Yuan, LI Xianjun, et al. Effects of heat treatment on color and equilibrium moisture content of mao-bamboo bundles[J]. J Cent South Univ For Technol, 2012, 32(9):138-141. [24] 高振华, 邸明伟.生物质材料及应用[M].北京:化学工业出版社, 2008. [25] PONCSÁK S, KOCAEFE D, BOUAZARA M, et al. Effect of high temperature treatment on the mechanical properties of birch (Betula papyrifera)[J]. Wood Sci Technol, 2006, 40(8):647-663. [26] MATAN N, KYOKONG B. Effect of moisture content on some physical and mechanical properties of juvenile rubberwood (Hevea brasiliensis Muell. Arg.)[J]. Songklanakarin J Sci Technol, 2003, 25(3):327-340. [27] 张雨, 王浚浩, 马中青, 等.温度对竹材烘焙过程中气固液三相产物组成及特性的影响[J].农业工程学报, 2018, 34(18):242-251. ZHANG Yu, WANG Junhao, MA Zhongqing, et al. Effects of torrefaction temperature on composition and characteristics of gas-solid-liquid three-phase products in bamboo torrefaction process[J]. Trans Chin Soc Agric Eng, 2018, 34(18):242-251. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2019.05.017
 点击查看大图
点击查看大图
计量
- 文章访问数: 3604
- HTML全文浏览量: 881
- PDF下载量: 42
- 被引次数: 0
