





-
随着城市化进程的加速,作为城市生态系统的重要组成部分,城市森林在改善小气候、净化大气、减弱噪音、改善人居环境等方面发挥着越来越重要的作用[1-3]。乔木作为城市森林群落的主体,决定了森林群落的结构、功能和演替方向。公园风景游憩林作为城市森林的重要组成部分,为人们提供了观赏和游憩的空间。胸径、树高和冠幅是乔木最重要的生长指标,反映了最基本群落林分结构,是用于预测群落林分结构发展的重要标志[4-7]。本研究对佛山市千灯湖公园、文华公园和平洲公园的风景游憩林经级、高度级和冠面积等级分布格局进行了研究,分析不同类型公园风景游憩林的优势树种和结构特征,揭示风景游憩林构建的规律和存在的问题,为佛山市城市公园风景游憩林群落结构调整和配置优化提供理论依据。
-
佛山市地处广东省中南部(22°38′~23°34′N,112°22′~113°23′E),珠江三角洲腹地,总面积为3 797.72 km2,是“粤港澳大湾区”“珠江·西江经济带”的重要组成部分。佛山市属南亚热带季风气候,雨热充沛,干湿分明,年均气温为22.5 ℃,年均降水量为1 681.2 mm。
城市森林样地分别位于南海区、禅城区和三山新城,为人工营建的公园风景游憩林。其中,千灯湖公园风景游憩林样地位于佛山市城市核心区;文华公园风景游憩林样地处于佛山城市次核心区;平洲公园风景游憩林位于佛山城郊,与广州毗邻。由于城市公园为人为营建的公园,3个公园从整体的规划设计风格上具有差异,千灯湖公园、文化公园、平洲公园的风格依次表现出由规则式设计向自然式设计过渡。
-
采用森林群落生态学研究方法,每个样地中样方数量的选取方法参照周红敏等[8]的研究成果,在考虑调查时间和成本情况下,设置样方大小为30 m×30 m,抽取样方4个·hm−2,以准确表达森林群落的空间分布格局。千灯湖公园研究区面积约4.95 万m2,文化公园研究区面积约10.12 万m2,平洲公园研究区面积约4.88 万m2。因此,本研究以900 m2样方为单元,分别在千灯湖公园风景游憩林群落样地设置调查样方20个,文华公园风景游憩林群落样地设置调查样方40个,平洲公园风景游憩林群落样地设置调查样方20个。所有样方在满足随机选取的前提下,应做到互不相邻,同时水面、建筑、纯草地面积占比1/3以上的样方无效。样方的植被调查包括每木检尺记录胸径DBH≥5.0 cm的树种名称、胸径、树高、冠幅、活枝下高、健康状况等指标。
-
计算不同类型公园风景游憩林的优势树种相对多度(RA)、相对频度(RF)、相对显著度(RD)和乔木重要值(IV)。RA=(某个种的株数/全部种的总株数)×100%;RF=(某个种的频度/所有种的频度总和)×100%;RD=(某个种的断面积/全部种的总断面积)
$ \times $ 100%;IV=(RA+RF+RD)/300%。 -
根据分布位置不同,将风景游憩林样方划分为核心林地和边缘林地。其中,核心林地分为核心有林地和核心疏林地(以下简称有林地和疏林地),边缘林地分为亲水林地和建筑旁林地。核心有林地为样方最远一边远离水体或建筑50 m以上,郁闭度0.20以上的林地;核心疏林地为样方最远一边远离水体或建筑50 m以上,郁闭度在0.10~0.19的林地,亲水林地为样方近水边距离水体小于50 m的林地,建筑旁林地为样方近建筑边距离建筑物小于50 m的林地。
-
根据统计学原理,样方乔木的径级(DBH)、高度级(H)和冠面积等级(CD)均采用上限排外法划分[7]。径级的划分方法:DBH处于5~10 cm,为第1径级,每5 cm为1个递增单位,当DBH处于10~40 cm时,每10 cm为1个递增单位;当DBH处于40~100 cm时,每20 cm为1个递增单位;DBH在100 cm以上时,为1个径级。高度级的划分方法:H处于3~5 m,为第1高度级,H处于5~7 m为第2高度级,每2 m为1个递增单位,当H处于11~20 m时,每3 m为1个递增单位;当H在20 m以上时,每4 m为1个递增单位。冠面积等级的划分方法:CD处于0~4 m2为第1等级,CD处于4~16 m2为第2等级,CD处于16~36 m2为第3等级,CD处于36~64 m2为第4等级,CD处于64~100 m2为第5等级,CD在100 m2以上为第6等级。
-
根据公园风景游憩林类型划分原则,将各公园风景游憩林类型进行划分。千灯湖公园核心有林地包含11个样方;核心疏林地包含2个样方;亲水林地包含5个样方;建筑旁林地包含2个样方。文华公园核心有林地包含9个样方;核心疏林地包含17个样方;亲水林地包含9个样方;建筑旁林地包含5个样方。平洲公园核心有林地包含7个样方;核心疏林地包含2个样方;亲水林地包含7个样方;建筑旁林地包含4个样方。各类型公园风景游憩林样方优势树种见表1。
表 1 不同类型风景游憩林优势树种分布
Table 1. Dominant species distribution of the different landscape-recreational forests
公园 类型 优势树种(重要值排名前5的树种) 千灯湖 有林地 非洲楝Khaya senegalensis(0.43)、樟树Cinnamomum camphora(0.19)、羊蹄甲Bauhinia purpurea(0.07)、木棉Bombax ceiba(0.07)、海南蒲桃Syzygium hainanense(0.04) 疏林地 小叶榕Ficus microcarpa(0.36)、非洲楝(0.17)、菩提榕Ficus religiosa(0.14)、黄花风铃木Handroanthus chrysanthus(0.12)、杧果Mangifera indica(0.12) 亲水林地 小叶榕(0.29)、樟树(0.15)、垂叶榕Ficus benjamina(0.085)、麻楝Chukrasia tabularis(0.085)、蒲葵Livistona chinensis(0.085) 建筑旁林地 非洲楝(0.38)、杧果(0.25)、糖胶树Alstonia scholaris(0.13)、樟树(0.10)、美丽针葵Phoenix canariensis(0.05) 文华 有林地 小叶榕(0.18)、长芒杜英Elaeocarpus apiculatus(0.07)、白兰Michelia×alba(0.07)、小叶榄仁Terminalia neotaliala(0.06)、秋枫Bischofia javanica(0.05) 疏林地 小叶榕(0.33)、大花紫薇Lagerstroemia speciosa(0.09)、木棉(0.09)、凤凰木Delonix regia(0.05)、无忧树Saraca indica(0.05) 亲水林地 菩提榕(0.33)、大花紫薇(0.08)、木棉(0.06)、羊蹄甲(0.06)、腊肠树Cassia fistula(0.04) 建筑旁林地 菩提榕(0.33)、狐尾椰子Wodyetia bifurcata(0.14)、小叶榄仁(0.10)、南洋杉Araucaria cunninghamii(0.10)、铁刀木Senna siamea(0.05) 平洲 有林地 高山榕Ficus altissima(0.26)、南洋楹Falcataria moluccana(0.08)、糖胶树Alstonia scholaris(0.05)、枫香Liquidambar formosana(0.05)、水石榕Elaeocarpus hainanensis(0.04) 疏林地 小叶榕(0.44)、白兰(0.36)、海枣Phoenix dactylifera(0.07)、三角梅Bougainvillea spectabilis(0.06)、四季桂Osmanthus fragrans(0.06) 亲水林地 菩提榕(0.31)、苏铁Cycas revoluta(0.08)、四季桂(0.06)、糖胶树(0.05)、大花紫薇(0.04) 建筑旁林地 高山榕(0.33)、白千层Melaleuca cajuputi(0.16)、木棉(0.12)、非洲楝(0.07)、小叶榕(0.05) 说明:括号内数值为该树种的重要值 -
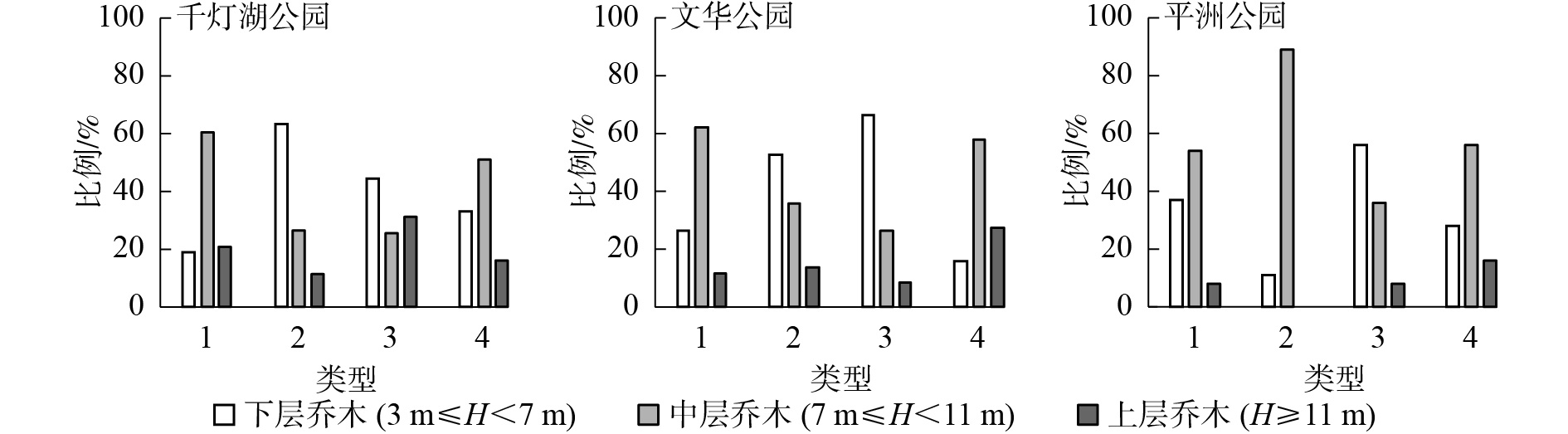
由图1可知:3个样地的风景游憩林基本以中、小径级比例占优势。千灯湖公园有林地群落构成多为中径级乔木,数量占比为72.4%。疏林地群落构成多为中、大径级乔木,中径级乔木数量占比为44.4%,大径级乔木数量占比为37.0%。亲水林地群落构成多为小、中径级乔木,小径级乔木数量占比为36.4%,中径级乔木数量占比为45.5%。建筑旁林地群落构成多为中径级乔木,数量占比为58.1%。文华公园有林地群落构成多为小、中径级乔木,小径级乔木数量占比为41.8%,中径级乔木数量占比为46.9%。疏林地群落构成多为小径级乔木,数量占比为49.5%。亲水林地群落构成多为小径级乔木,数量占比为56.4%。建筑旁林地群落构成多为中径级乔木,数量占比为63.6%。平洲公园有林地群落构成多为小径级乔木,数量占比为48.1%。疏林地群落构成多为中径级乔木,数量占比为55.6%。亲水林地群落构成多为小、中径级乔木,小径级乔木数量占比为39.3%,中径级乔木数量占比为37.2%。建筑旁林地群落构成多为中径级乔木,数量占比为45.4%。

图 1 不同类型风景游憩林径级分布
Figure 1. Diameter class distribution of the different landscape-recreational forests
从表2可知:3个样地风景游憩林的树种密度从大到小依次为有林地、建筑旁林地、亲水林地、疏林地。千灯湖公园4种风景游憩林的大径级树种主要为小叶榕、非洲楝,较为单一;文华公园和平洲公园的大径级树种较为丰富;不同径级之间的树种并无明显差异。
表 2 风景游憩林不同径级的树种
Table 2. Tree species of the different diameter class of landscape-recreational forests
公园 类型 树种密度/
(株·hm−2)小径级树种
(5 cm≤DBH<20 cm)中径级树种
(20 cm≤DBH<40 cm)大径级树种
(DBH≥40 cm)千灯湖 有林地 292.9 樟树(23.1) 非洲楝(25.2)、樟树(20.5) 非洲楝(51.7) 疏林地 150.0 黄花风铃木(100.0) 小叶榕(75.0)、杧果(25.0) 小叶榕(37.5)、非洲楝(25.0) 亲水林地 195.6 樟树(40.6)、黄花风铃木(25.0) 樟树(20.0)、麻楝(20.0) 小叶榕(25.0) 建筑旁林地 238.9 美丽针葵(42.9)、
大花紫薇(28.6)杧果(44.0)、糖胶树(24.0)、
樟树(24.0)非洲楝(63.6)、杧果(27.3) 文华 有林地 313.3 白兰(24.2) 长芒杜英(12.6) 秋枫(24.1)、樟树(20.7) 疏林地 225.7 大花紫薇(22.7) 木棉(26.6) 木棉(22.8) 亲水林地 249.4 大花紫薇(20.3) 木棉(27.6) 黄葛榕Ficus virens(20.0) 建筑旁林地 311.1 南洋杉(56.8) 小叶榄仁(23.6) 狐尾椰子(36.0) 平洲 有林地 252.4 枫香(17.1) 高山榕(13.7)、木棉(13.7)、
蒲葵(13.7)高山榕(39.7) 疏林地 50.0 白兰(100.0) 白兰(100.0) 白兰(66.7)、小叶榕(22.2) 亲水林地 230.2 四季桂(29.8) 蒲葵(16.7) 糖胶树(23.5)、银海枣Phoenix sylvestris(20.6) 建筑旁林地 238.9 木棉(22.6) 白千层(35.9) 非洲楝(25.0) 说明:括号内数值为各径级优势树种所占比例(%) -
从图2可知:3个样地的风景游憩林基本以中、下层乔木比例占优势。千灯湖公园有林地群落构成以中层乔木为主,数量占比为60.3%。疏林地群落构成以下层乔木树种为主,数量占比为63.0%。亲水林地群落构成以下层乔木树种为主,数量占比为44.3%。建筑旁林地群落构成以中层乔木树种为主,数量占比为51.2%。文华公园有林地群落构成以中层乔木树种为主,数量占比为62.1%。疏林地群落构成以下层乔木树种为主,数量占比为52.0%。亲水林地群落构成以下层乔木树种为主,数量占比为66.3%。建筑旁林地群落构成以中层乔木树种为主,数量占比为57.1%。平洲公园有林地群落构成以中层乔木树种为主,中层乔木数量占比为54.4%。疏林地群落构成以中层乔木树种为主,无上层乔木,中层乔木数量占比为88.9%。亲水林地群落构成以下层乔木树种为主,下层乔木数量占比为56.4%。建筑旁林地群落构成以中层乔木树种为主,中层乔木数量占比为55.8%。
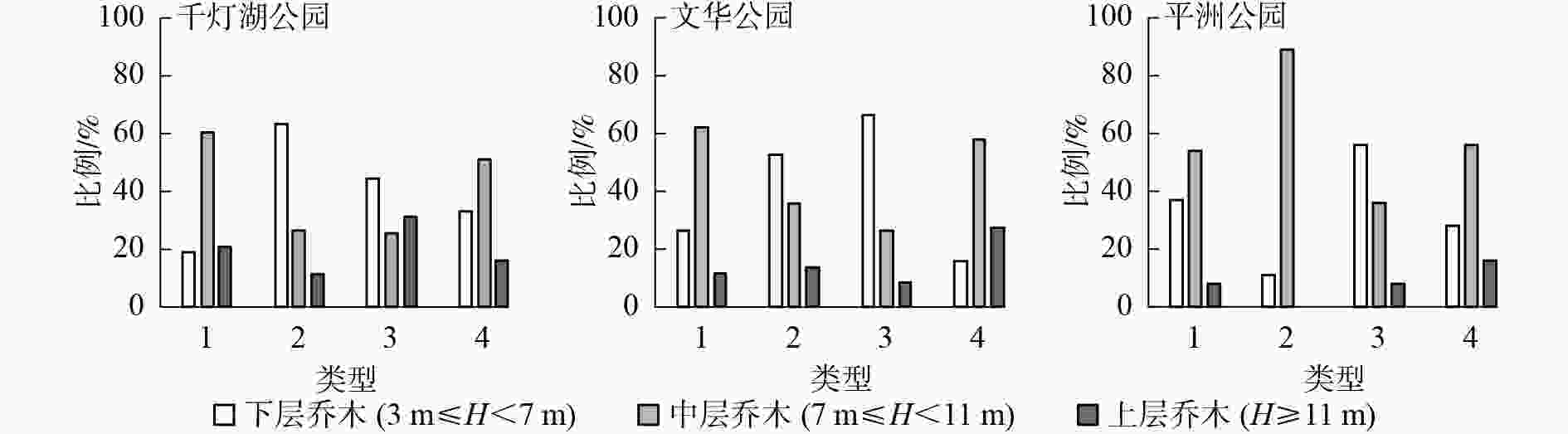
图 2 不同类型风景游憩林高度级分布
Figure 2. Height class distribution of the different landscape-recreational forests
由表3可知:千灯湖公园4种风景游憩林的上层乔木树种主要为非洲楝,较为单一;文华公园和平洲公园的上层乔木树种较为丰富;随着由高度级的降低,不同风景游憩林类型的树种差异逐渐增大,树种色彩丰富度也逐渐增强;千灯湖公园的风景游憩林高度级差异最大,文化公园中疏林地和亲水林地的高度级分布一致,平洲公园中有林地和疏林地(即核心林地)的高度级分布一致,亲水林地和建筑旁林地(即边缘林地)高度级分布一致。
表 3 风景游憩林不同高度级树种
Table 3. Tree species of the different height class of landscape-recreational forests
公园 类型 下层乔木树种(3 m≤H<7 m) 中层乔木树种(7 m≤H<11 m) 上层乔木树种(H≥11 m) 千灯湖 有林地 木棉(22.2) 非洲楝(17.1) 非洲楝(59.0) 疏林地 小叶榕(64.7)、黄花风铃木(29.4) 杧果(100.0) 非洲楝(66.7)、菩提榕(33.3) 亲水林地 蒲葵(25.6)、黄花风铃木(20.5) 樟树(31.8) 樟树(29.6)、非洲楝(25.9) 建筑旁林地 杧果(57.1)、美丽针葵(21.4) 糖胶树(36.4)、杧果(27.3)、樟树(27.3) 非洲楝(100.0) 文华 有林地 白兰(39.2) 长芒杜英(16.7) 凤凰木(25.6) 疏林地 大花紫薇(25.1) 木棉(25.7) 小叶榄仁(31.0)、木棉(28.6) 亲水林地 大花紫薇(19.0) 木棉(28.3)、樟树(22.6) 南洋楹(35.3)、菩提榕(23.5) 建筑旁林地 垂叶榕(29.2) 狐尾椰子(40.0)、南洋杉(22.5) 小叶榄仁(50.0)、秋枫(23.7) 平洲 有林地 竹柏(11.7)、美丽针葵(11.7) 高山榕(23.4) 南洋楹(34.6) 疏林地 银海枣(100.0) 白兰(75.0)、小叶榕(25.0) 亲水林地 四季桂(20.5) 蒲葵(24.3) 大王椰子(21.4) 建筑旁林地 木棉(24.5) 白千层(47.4) 白千层(57.1)、非洲楝(35.7) 说明:括号内数值为各高度级优势树种所占比例(%) -
从图3可知:3个样地的风景游憩林基本以中冠面积等级的乔木比例占优势。千灯湖公园有林地群落构成以中树冠面积的乔木为主,中树冠面积乔木数量占比为70.8%。疏林地群落构成以中树冠面积的乔木为主,中树冠面积乔木数量占比为63.0%。亲水林地群落构成以小、中树冠面积乔木为主,小树冠面积乔木数量占比为39.4%,中树冠面积乔木数量占比为45.8%。建筑旁林地群落构成以中树冠面积的乔木为主,中树冠面积乔木数量占比为67.4%。文华公园有林地群落构成以中冠面积的乔木为主,中冠面积乔木数量占比为60.6%。疏林地群落构成以中冠面积的乔木为主,中冠面积乔木数量占比为63.4%。亲水林地群落构成以中冠面积的乔木为主,中冠面积乔木数量占比为53.5%。建筑旁林地群落构成以中面积的乔木为主,中冠面积乔木数量占比为70.0%。平洲公园有林地群落构成以中树冠面积的乔木为主,中树冠面积乔木数量占比为61.4%。疏林地群落构成以中树冠面积的乔木为主,中树冠面积乔木数量占比为77.8%,无小树冠面积乔木。亲水林地群落构成以中树冠面积的乔木为主,中树冠面积乔木数量占比为59.7%。建筑旁林地群落构成以中树冠面积的乔木为主,中树冠面积乔木数量占比为68.6%。
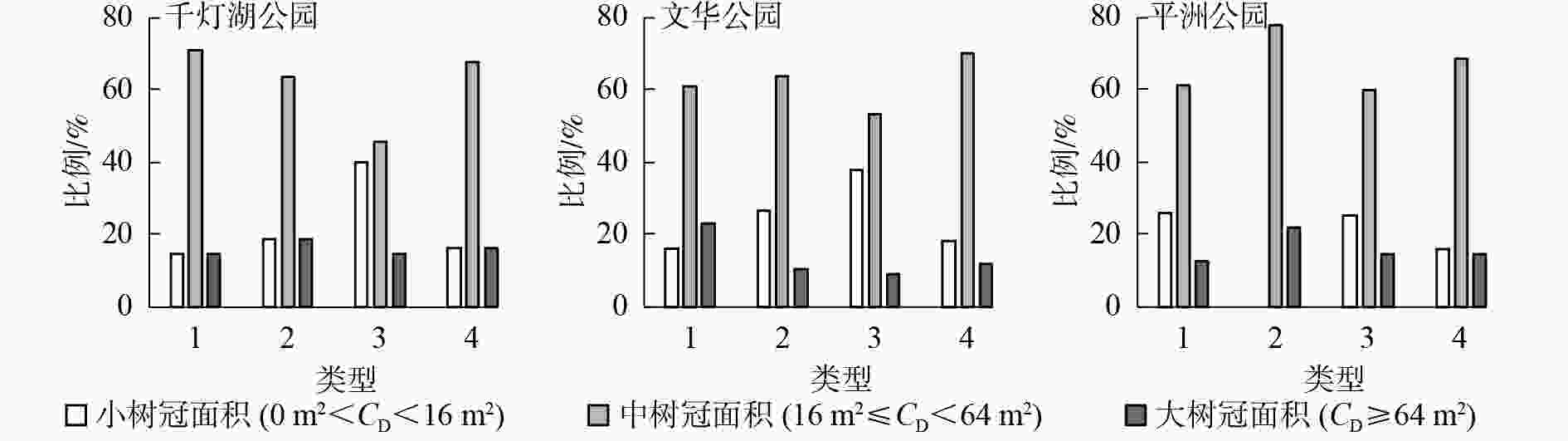
图 3 不同类型风景游憩林冠面积等级分布
Figure 3. Canopy class distribution of the different landscape-recreational forests
由表4可知:千灯湖公园4种风景游憩林的大冠面积等级树种主要为非洲楝,较为单一;文华公园和平洲公园的大冠面积等级树种较为丰富;随着由冠面积等级的降低,不同风景游憩林类型的树种差异逐渐增大,树种色彩丰富度也逐渐增强;千灯湖公园的有林地和疏林地(核心林地)、文华公园的有林地和疏林地(核心林地)以及亲水林地和建筑旁林地(边缘林地)、平洲公园的亲水林地和建筑旁林地(边缘林地)内部冠面积等级分布均表现较为一致。
表 4 风景游憩林不同冠面积等级的树种
Table 4. Tree species of the different canopy class of landscape-recreational forests
公园 类型 小冠面积树种
(0 m2≤CD<16 m2)中冠面积树种
(16 m2≤CD<64 m2)大冠面积树种
(CD≥64 m2)千灯湖 有林地 非洲楝(25.0)、樟树(20.5) 羊蹄甲(26.2)、樟树(24.3) 非洲楝(26.7) 疏林地 黄花风铃木(100.0) 小叶榕(72.2)、杧果(22.2) 非洲楝(40.0) 亲水林地 黄花风铃木(21.6) 樟树(34.1) 非洲楝(28.6)、小叶榕(21.4) 建筑旁林地 美丽针葵(42.9)、大花紫薇(28.6) 杧果(48.3)、糖胶树(27.6)、
樟树(20.7)非洲楝(100.0) 文华 有林地 白兰(63.0) 长芒杜英(17.9) 小叶榄仁(23.1) 疏林地 非洲楝(25.0)、樟树(20.5) 大花紫薇(18.8) 木棉(27.3) 亲水林地 散尾葵Dypsis lutescens(13.8) 大花紫薇(20.2) 南洋楹(27.8)、菩提榕(27.8) 建筑旁林地 南洋杉(24.2)、散尾葵(24.2) 狐尾椰子(32.7) 小叶榄仁(47.1)、铁刀木(23.5) 平洲 有林地 美丽针葵(14.3)、龙柏Juniperus chinensis(14.3) 枫香(12.4) 南洋楹(35.0)、高山榕(25.0) 疏林地 白兰(85.7) 小叶榕(100.0) 亲水林地 四季桂(44.4) 大花紫薇(13.5)、糖胶树13.5) 黄葛榕(18.2)、小叶榕(18.2) 建筑旁林地 荔枝Litchi chinensis(30.8) 白千层(30.5)、木棉(22.0) 非洲楝(46.2)、高山榕(23.1) 说明:括号内数值为各冠面积等级优势树种所占比例(%) -
森林群落的径级、高度级和冠面积等级是最基本的群落林分结构指标,能综合反映植物生长与环境的关系,是用于预测群落林分结构发展的重要指标[4, 9-12]。谢春平等[13]对南京仙林大学城公共绿地的研究表明:仙林公园绿地乔木以中、小径级占多数;随着径级的增大,数量呈递减趋势,径级大于18 cm的树木零星分布。由于大学城的开发时间较短,因此大径级乔木偏少,这与本研究的结果相似。除了位于城市核心区的千灯湖公园建筑旁林地外,本研究的其他样地乔木径级多分布于中、小径级。中、小径级乔木在生态功能和景观效果上不如大径级的乔木,但更新能力强,由于处于快速成长期,中、小径级乔木的更新能力将决定群落的未来发展[14-15]。
本研究样地的疏林地和亲水林地高度级基本呈“前大后小”分布,中、下层乔木所占比例较大,千灯湖公园亲水林地的下层乔木和上层乔木分布比例多,中层乔木分布比例少;而有林地和建筑旁林地的中层乔木较多,其次为下层乔木。刘秀萍[16]对北京城区的城市森林分析表明:各环路乔木高度平均值均由城中心到城外围逐渐降低,这是由建设时间和绿化时间引起的,城中心的绿化建设时间比较早,这与本研究的结果较为一致。千灯湖公园的平均树高(8.39 m)最高,其次为文华公园,平均树高为7.53 m,平洲公园的平均树高(7.20 m)最小,表明随着风景游憩林位置由城市核心区、次核心区、城郊的过渡,其树高平均值逐渐降低。
李晓婷[17]对位于北京各环路高校乔木的平均冠面积研究表明:乔木平均冠面积随着环路向外扩张,平均冠面积由城中心到城外围逐渐减小。本研究与李晓婷[17]的研究结果有所不同,位于城市核心区的千灯湖公园平均冠面积(41.23 m2)是最大的,其次为城郊的平洲公园(平均冠面积37.08 m2),平均冠面积最小的为城市次核心区文华公园(34.40 m2)。导致这种现象的原因很可能是城市森林中乔木的各个生长指标均受到较强的人为干扰,而树冠作为受人为干扰影响最强的生长指标,城市绿化管理部门会周期性的进行树冠修剪工作,如仅仅采用静态的冠面积数据来反映不同生态位森林群落的冠面积分布趋势是有局限性的。因此,须开展长期的动态冠面积监测才能反映真实的冠面积分布趋势。
城市森林与郊野森林有显著的区别,即城市森林与城市相互融合,人为干扰强烈,群落结构较为单一,景观异质性强,富含人文景观价值[18-20],而郊野森林人为干扰小,群落结构较为复杂,景观异质性弱。城市森林和郊野森林的群落特征不同,会导致研究过程中所借助的研究方法和手段有所差异。郊野森林在满足最小样方面积和数量的基础上,一般选择随机取样的方式。本研究的取样方法是建立在周红敏等[8]对郊野森林随机取样方法研究成果的基础上,根据群落异质性的不同,针对城市森林的特殊性进行调整,即满足每1 万m2抽取4个样方的条件下,样方的选取应避开水面、建筑以及纯草地,并应做到互不相邻,这种样方设置方法更适合城市森林的研究。
-
本研究表明:3个样地森林群落均具备热带或亚热带季风性气候植物区系特征。有林地的树种数量多,以常绿树种为主,搭配少量观花树种和棕榈科Palmae植物;而疏林地的树种数量相对较少,主要为常绿树种;亲水林地的树种主要为常绿树种,搭配少量观花树种和棕榈科植物,小乔木数量较多;建筑旁林地主要为常绿树种,搭配棕榈科植物,有少量高大常绿乔木。从风景游憩林径级分布上分析,核心林地和边缘林地内部径级分布均表现出差异,随着城市核心区、次核心区、城郊的过渡,风景游憩林高度级分布差异逐渐变小;从风景游憩林冠面积分布上分析,核心林地、边缘林地内部冠面积分布表现出一定的相似性。
Community structure characteristics for landscape-recreational forests in the urban parks of Foshan City
-
摘要:
目的 明确不同生态位风景游憩林的优势树种和结构特征,可揭示风景游憩林构建的规律和存在的问题。 方法 采用群落生态学的方法,以900 m2样方为单元,分别在佛山千灯湖公园(20个样方)、文华公园(40个样方)、平洲公园(20个样方)设置风景游憩林群落样地,结合高分遥感影像,进行每木调查。先对径级、高度级和冠面积划分等级,再进行不同类型风景游憩林的差异分析。 结果 ①千灯湖公园的大径级、大高度级和大冠面积等级的主要树种为非洲楝Khaya senegalensis,均比文华公园和平洲公园单一;②在3个样地的有林地和建筑旁林地中,中径级乔木占相对优势明显,大径级乔木占比最少,疏林地和亲水林地小径级乔木占相对优势明显,大径级乔木占比最少;③在3个样地的有林地和建筑旁林地中,中层乔木占比最多,上层乔木占比最少,疏林地和亲水林地下层乔木相对优势明显,上层乔木占比最少;④在3个样地的风景游憩林中,中冠面积等级乔木占比最多,大冠幅等级占比最少。 结论 3个样地森林群落均具备热带或亚热带季风性气候植物区系特征。3个样地的有林地和疏林地(核心林地)、亲水林地和建筑旁林地(边缘林地)内部径级分布均表现出差异。风景游憩林所在位置由城市核心区、次核心区、城郊的过渡,其高度级分布差异逐渐变小。由于树冠是受人为干扰影响最强的生长指标,须开展长期的动态冠面积监测才能反映真实的分布趋势。图3表4参20 Abstract:Objective With an attempt at the clarification of the dominant species and structural characteristics of landscape-recreational forests in different ecological niches, the current paper is aimed to conclude the rules and research the existing problems of the construction of landscape-recreational forests. Method With the method of community ecology employed and 20 sample plots in Qiandeng Lake Park, 40 sample plots in Wenhua Park, and 20 sample plots in the Pingzhou Park selected as researching areas, the research was conducted with every tree surveyed using high-resolution remote sensing images. After the diameter classes, height classes and crown areas were clarified, an analysis was made of the differences among different types of landscapes. Result (1) Compared with Wenhua Park and Pingzhou Park, Qiandeng Lake Park enjoys a higher level of singleness in tress species with Khaya senegalensis as the dominant tree of large diameter, height, and canopy class; (2) Landscape-recreational forests on closed forest land and besides the buildings are mainly composed of medium-diameter trees while those on in the open forest land and besides the water are mainly composed of small diameter trees with larger diameter trees taking up a small percentage; (3) Landscape-recreational forests in the closed forest land and besides the buildings are mainly composed of the medium-height trees while, those in the open forest land and besides the water are mainly composed of small height trees, with large height trees taking up a small percentage; (4) Landscape-recreational forests are mainly composed of the medium canopy class trees with, the large canopy trees taking up only a small percentage. Conclusion To sum up, floristic characteristics of tropical or subtropical monsoon are all shown in the landscape-recreational forests of the three parks. There are differences in the distribution of diameter classes among the trees in the closed forest land and open forest land (the center forest land) as well as those besides the water and buildings (fringe forest land). The differences in the distribution of height class are gradually decreased with the location of landscape-recreational forests shifted from the downtown to subsidiary center and finally to the suburbs. It is necessary to conduct long-term dynamic canopy monitoring to reflect the trend of canopy distribution. [Ch, 3 fig. 4 tab. 20 ref.] -
Key words:
- forest ecology /
- dominant species /
- landscape-recreational forests /
- diameter class /
- height class /
- canopy class /
- urban forest
-
图 1 不同类型风景游憩林径级分布
1. 有林地;2. 疏林地;3. 亲水林地;4. 建筑旁林地
Figure 1 Diameter class distribution of the different landscape-recreational forests
图 2 不同类型风景游憩林高度级分布
1. 有林地;2. 疏林地;3. 亲水林地;4. 建筑旁林地
Figure 2 Height class distribution of the different landscape-recreational forests
图 3 不同类型风景游憩林冠面积等级分布
1. 有林地;2. 疏林地;3. 亲水林地;4. 建筑旁林地
Figure 3 Canopy class distribution of the different landscape-recreational forests
表 1 不同类型风景游憩林优势树种分布
Table 1. Dominant species distribution of the different landscape-recreational forests
公园 类型 优势树种(重要值排名前5的树种) 千灯湖 有林地 非洲楝Khaya senegalensis(0.43)、樟树Cinnamomum camphora(0.19)、羊蹄甲Bauhinia purpurea(0.07)、木棉Bombax ceiba(0.07)、海南蒲桃Syzygium hainanense(0.04) 疏林地 小叶榕Ficus microcarpa(0.36)、非洲楝(0.17)、菩提榕Ficus religiosa(0.14)、黄花风铃木Handroanthus chrysanthus(0.12)、杧果Mangifera indica(0.12) 亲水林地 小叶榕(0.29)、樟树(0.15)、垂叶榕Ficus benjamina(0.085)、麻楝Chukrasia tabularis(0.085)、蒲葵Livistona chinensis(0.085) 建筑旁林地 非洲楝(0.38)、杧果(0.25)、糖胶树Alstonia scholaris(0.13)、樟树(0.10)、美丽针葵Phoenix canariensis(0.05) 文华 有林地 小叶榕(0.18)、长芒杜英Elaeocarpus apiculatus(0.07)、白兰Michelia×alba(0.07)、小叶榄仁Terminalia neotaliala(0.06)、秋枫Bischofia javanica(0.05) 疏林地 小叶榕(0.33)、大花紫薇Lagerstroemia speciosa(0.09)、木棉(0.09)、凤凰木Delonix regia(0.05)、无忧树Saraca indica(0.05) 亲水林地 菩提榕(0.33)、大花紫薇(0.08)、木棉(0.06)、羊蹄甲(0.06)、腊肠树Cassia fistula(0.04) 建筑旁林地 菩提榕(0.33)、狐尾椰子Wodyetia bifurcata(0.14)、小叶榄仁(0.10)、南洋杉Araucaria cunninghamii(0.10)、铁刀木Senna siamea(0.05) 平洲 有林地 高山榕Ficus altissima(0.26)、南洋楹Falcataria moluccana(0.08)、糖胶树Alstonia scholaris(0.05)、枫香Liquidambar formosana(0.05)、水石榕Elaeocarpus hainanensis(0.04) 疏林地 小叶榕(0.44)、白兰(0.36)、海枣Phoenix dactylifera(0.07)、三角梅Bougainvillea spectabilis(0.06)、四季桂Osmanthus fragrans(0.06) 亲水林地 菩提榕(0.31)、苏铁Cycas revoluta(0.08)、四季桂(0.06)、糖胶树(0.05)、大花紫薇(0.04) 建筑旁林地 高山榕(0.33)、白千层Melaleuca cajuputi(0.16)、木棉(0.12)、非洲楝(0.07)、小叶榕(0.05) 说明:括号内数值为该树种的重要值  下载: 导出CSV
下载: 导出CSV
表 2 风景游憩林不同径级的树种
Table 2. Tree species of the different diameter class of landscape-recreational forests
公园 类型 树种密度/
(株·hm−2)小径级树种
(5 cm≤DBH<20 cm)中径级树种
(20 cm≤DBH<40 cm)大径级树种
(DBH≥40 cm)千灯湖 有林地 292.9 樟树(23.1) 非洲楝(25.2)、樟树(20.5) 非洲楝(51.7) 疏林地 150.0 黄花风铃木(100.0) 小叶榕(75.0)、杧果(25.0) 小叶榕(37.5)、非洲楝(25.0) 亲水林地 195.6 樟树(40.6)、黄花风铃木(25.0) 樟树(20.0)、麻楝(20.0) 小叶榕(25.0) 建筑旁林地 238.9 美丽针葵(42.9)、
大花紫薇(28.6)杧果(44.0)、糖胶树(24.0)、
樟树(24.0)非洲楝(63.6)、杧果(27.3) 文华 有林地 313.3 白兰(24.2) 长芒杜英(12.6) 秋枫(24.1)、樟树(20.7) 疏林地 225.7 大花紫薇(22.7) 木棉(26.6) 木棉(22.8) 亲水林地 249.4 大花紫薇(20.3) 木棉(27.6) 黄葛榕Ficus virens(20.0) 建筑旁林地 311.1 南洋杉(56.8) 小叶榄仁(23.6) 狐尾椰子(36.0) 平洲 有林地 252.4 枫香(17.1) 高山榕(13.7)、木棉(13.7)、
蒲葵(13.7)高山榕(39.7) 疏林地 50.0 白兰(100.0) 白兰(100.0) 白兰(66.7)、小叶榕(22.2) 亲水林地 230.2 四季桂(29.8) 蒲葵(16.7) 糖胶树(23.5)、银海枣Phoenix sylvestris(20.6) 建筑旁林地 238.9 木棉(22.6) 白千层(35.9) 非洲楝(25.0) 说明:括号内数值为各径级优势树种所占比例(%)
下载: 导出CSV
表 3 风景游憩林不同高度级树种
Table 3. Tree species of the different height class of landscape-recreational forests
公园 类型 下层乔木树种(3 m≤H<7 m) 中层乔木树种(7 m≤H<11 m) 上层乔木树种(H≥11 m) 千灯湖 有林地 木棉(22.2) 非洲楝(17.1) 非洲楝(59.0) 疏林地 小叶榕(64.7)、黄花风铃木(29.4) 杧果(100.0) 非洲楝(66.7)、菩提榕(33.3) 亲水林地 蒲葵(25.6)、黄花风铃木(20.5) 樟树(31.8) 樟树(29.6)、非洲楝(25.9) 建筑旁林地 杧果(57.1)、美丽针葵(21.4) 糖胶树(36.4)、杧果(27.3)、樟树(27.3) 非洲楝(100.0) 文华 有林地 白兰(39.2) 长芒杜英(16.7) 凤凰木(25.6) 疏林地 大花紫薇(25.1) 木棉(25.7) 小叶榄仁(31.0)、木棉(28.6) 亲水林地 大花紫薇(19.0) 木棉(28.3)、樟树(22.6) 南洋楹(35.3)、菩提榕(23.5) 建筑旁林地 垂叶榕(29.2) 狐尾椰子(40.0)、南洋杉(22.5) 小叶榄仁(50.0)、秋枫(23.7) 平洲 有林地 竹柏(11.7)、美丽针葵(11.7) 高山榕(23.4) 南洋楹(34.6) 疏林地 银海枣(100.0) 白兰(75.0)、小叶榕(25.0) 亲水林地 四季桂(20.5) 蒲葵(24.3) 大王椰子(21.4) 建筑旁林地 木棉(24.5) 白千层(47.4) 白千层(57.1)、非洲楝(35.7) 说明:括号内数值为各高度级优势树种所占比例(%)
下载: 导出CSV
表 4 风景游憩林不同冠面积等级的树种
Table 4. Tree species of the different canopy class of landscape-recreational forests
公园 类型 小冠面积树种
(0 m2≤CD<16 m2)中冠面积树种
(16 m2≤CD<64 m2)大冠面积树种
(CD≥64 m2)千灯湖 有林地 非洲楝(25.0)、樟树(20.5) 羊蹄甲(26.2)、樟树(24.3) 非洲楝(26.7) 疏林地 黄花风铃木(100.0) 小叶榕(72.2)、杧果(22.2) 非洲楝(40.0) 亲水林地 黄花风铃木(21.6) 樟树(34.1) 非洲楝(28.6)、小叶榕(21.4) 建筑旁林地 美丽针葵(42.9)、大花紫薇(28.6) 杧果(48.3)、糖胶树(27.6)、
樟树(20.7)非洲楝(100.0) 文华 有林地 白兰(63.0) 长芒杜英(17.9) 小叶榄仁(23.1) 疏林地 非洲楝(25.0)、樟树(20.5) 大花紫薇(18.8) 木棉(27.3) 亲水林地 散尾葵Dypsis lutescens(13.8) 大花紫薇(20.2) 南洋楹(27.8)、菩提榕(27.8) 建筑旁林地 南洋杉(24.2)、散尾葵(24.2) 狐尾椰子(32.7) 小叶榄仁(47.1)、铁刀木(23.5) 平洲 有林地 美丽针葵(14.3)、龙柏Juniperus chinensis(14.3) 枫香(12.4) 南洋楹(35.0)、高山榕(25.0) 疏林地 白兰(85.7) 小叶榕(100.0) 亲水林地 四季桂(44.4) 大花紫薇(13.5)、糖胶树13.5) 黄葛榕(18.2)、小叶榕(18.2) 建筑旁林地 荔枝Litchi chinensis(30.8) 白千层(30.5)、木棉(22.0) 非洲楝(46.2)、高山榕(23.1) 说明:括号内数值为各冠面积等级优势树种所占比例(%)
下载: 导出CSV
-
[1] 汪永英,孔令伟,李雯,等. 哈尔滨城市森林小气候状况及对人体舒适度的影响[J]. 东北林业大学学报, 2012, 40(7): 90 − 93. WANG Yongying, KONG Lingwei, LI Wen, et al. Status of microclimate in urban forests in harbin and its effects on human comfort degree [J]. J Northeast For Univ, 2012, 40(7): 90 − 93. [2] 徐文铎,何兴元,陈玮,等. 沈阳城市森林小气候特征的研究[J]. 应用生态学报, 2005, 16(9): 1650 − 1654. XU Wenduo, HE Xingyuan, CHEN Wei, et al. Microclimate characters of urban forest in Shenyang City [J]. Chin J Appl Ecol, 2005, 16(9): 1650 − 1654. [3] 吴泽民,吴文友,高健,等. 合肥市区城市森林景观格局分析[J]. 应用生态学报, 2003, 14(12): 2117 − 2122. WU Zenmin, WU Wenyou, GAO Jian, et al. Analysis of forest landscape pattern in Hefei [J]. Chin J Appl Ecol, 2003, 14(12): 2117 − 2122. [4] 韩文娟,张文辉,何景峰,等. 不同坡向油松人工林建群种种群结构及群落特征分析[J]. 西北农林科技大学学报(自然科学版), 2012, 40(3): 47 − 55. HAN Wenjuan, ZHANG Wenhui, HE Jingfeng, et al. Analysis of population structure and plantation community characteristics of Pinus tabulaeformis plantations in Loess Plateau in different slope [J]. J Northwest A&F Univ Nat Sci Ed, 2012, 40(3): 47 − 55. [5] 郝建锋,王德艺,唐永彬,等. 人为干扰对江油地区马尾松人工林群落结构和物种多样性的影响[J]. 生态环境学报, 2014, 23(5): 729 − 735. HAO Jianfeng, WANG Deyi, TANG Yongbin, et al. Effects of human disturbance on species diversity of Pinus massoniana plantation in Jiangyou district, Sichuan Province [J]. Ecol Environ Sci, 2014, 23(5): 729 − 735. [6] 余义亮,丁彦芬,朱贵珍,等. 扬州地区乡村绿化植物群落的结构特征[J]. 浙江农林大学学报, 2019, 36(3): 541 − 549. YU Yiliang, DING Yanfen, ZHU Guizhen, et al. Structural characteristics of rural greening plant communities in Yangzhou [J]. J Zhejiang A&F Univ, 2019, 36(3): 541 − 549. [7] 芦建国,景蕾. 南京市公园绿地苔藓植物多样性及特点[J]. 浙江农林大学学报, 2019, 36(3): 486 − 493. LU Jianguo, JING Lei. Biodiversity and characteristics of ground bryophytes in 11 parks of Nanjing [J]. J Zhejiang A&F Univ, 2019, 36(3): 486 − 493. [8] 周红敏,惠刚盈,赵中华,等. 森林结构调查中最适样方面积和数量的研究[J]. 林业科学研究, 2009, 22(4): 482 − 495. ZHOU Hongmin, HUI Gangyin, ZHAO Zhonghua, et al. Studies on the area and the number of the sample for forest structure [J]. For Res, 2009, 22(4): 482 − 495. [9] 邵晓莉,程毅康,王茜茜,等. 海南岛热带云雾林地上生物量分布规律[J]. 生态学杂志, 2018, 37(9): 2566 − 2572. SHAO Xiaoli, CHENG Yikang, WANG Xixi, et al. Distribtution patterns of aboveground biomass of tropical cloud forests in Hainan Island [J]. Chin J Ecol, 2018, 37(9): 2566 − 2572. [10] 汪殿蓓,暨淑仪,陈飞鹏,等. 深圳南山区天然森林群落多样性及演替现状[J]. 生态学报, 2003, 23(7): 1415 − 1422. WANG Dianpei, JI Shuyi, CHEN Feipeng, et al. A study on the species diversity and succession situation of natural forest communities in Nanshan District, Shenzhen City [J]. Acta Ecol Sin, 2003, 23(7): 1415 − 1422. [11] 王帅,高尔刚,罗小菊,等. 施秉典型常绿阔叶林森林群落特征[J]. 分子植物育种, 2019, 17(10): 3432 − 3441. WANG Shuai, GAO Ergang, LUO Xiaoju, et al. Characteristics of forest communities of typical evergreen broad-leaved forest in Shibing [J]. Mol Plant Breed, 2019, 17(10): 3432 − 3441. [12] 于军,白冠章,梁继业,等. 塔里木河上、中、下游胡杨种群高度结构特征[J]. 干旱区资源与环境, 2012, 26(7): 103 − 109. YU Jun, BAI Guangzhang, LIANG Jiye, et al. The height structure characteristics of Populus euphraticu population along Tarim River [J]. J Arid Land Resour Environ, 2012, 26(7): 103 − 109. [13] 谢春平,田甜,伊贤贵,等. 南京仙林大学城公共绿地木本植物多样性分析[J]. 云南农业大学学报(自然科学), 2018, 33(6): 993 − 1002. XIE Chunping, TIAN Tian, YI Xiangui, et al. Analysis of woody species diversity of public green space in Xianlin College Town, Nanjing City [J]. J Yunnan Agric Univ Nat Sci, 2018, 33(6): 993 − 1002. [14] 翟石磊,陈步峰,林娜,等. 广州市典型森林植被的结构及多样性研究[J]. 生态环境学报, 2015, 24(10): 1625 − 1633. ZHAI Shilei, CHEN Bufeng, LIN Na, et al. Structure and species diversity of typical forests in Cuangzhou City [J]. Ecol Environ Sci, 2015, 24(10): 1625 − 1633. [15] 万媛媛,李洪远,贺梦璇,等. 基于不同铺装条件下的生境片断化对城市绿地中臭椿幼苗自然更新的影响[J]. 应用与环境生物学报, 2017, 23(6): 1110 − 1116. WAN Yuanyuan, LI Hongyuan, HE Mengxuan, et al. Influences of habitat fragmentation on natural regeneration of Ailanthus altissima in urban greenland based on different pavement conditions [J]. Chin J Appl Environ Biol, 2017, 23(6): 1110 − 1116. [16] 刘秀萍. 北京城区居住区和机关单位城市森林结构调查与树冠覆盖动态分析[D]. 北京: 中国林业科学研究院, 2017. LIU Xiuping. A Study on Urban Forest Structure and Urban Tree Canopy of Residential Areas and Public Institutions in Beijing Urban Area[D]. Beijing: Chinese Academy of Forestry, 2017. [17] 李晓婷. 北京城区校园与医院林木树冠覆盖与城市森林结构研究[D]. 北京: 中国林业科学研究院, 2018. LI Xiaoting. A Study on Urban Tree Canopy and Urban Forest Structure of Schools and Hospitals in Beijing Urban Area[D]. Beijing: Chinese Academy of Forestry, 2018. [18] 杨学军,许东新,金为民,等. 上海城市森林生态网络系统工程体系建设初探[J]. 上海农学院学报, 2000, 18(2): 132 − 137. YANG Xuejun, XU Dongxin, JIN Weimin, et al. Probe for the construction of the urban forest eco-network system engineering in Shanghai [J]. J Shanghai Agric Coll, 2000, 18(2): 132 − 137. [19] 刘常富,李海梅,何兴元,等. 城市森林概念探析[J]. 生态学杂志, 2003, 22(5): 146 − 149. LIU Changfu, LI Haimei, HE Xingyuan, et al. Concept discussion and analysis of urban forest [J]. Chin J Ecol, 2003, 22(5): 146 − 149. [20] 何荣晓,杨帆,崔明. 海口市城市森林结构及植物多样性指标相关性分析[J]. 浙江农林大学学报, 2019, 36(6): 1142 − 1150. HE Rongxiao, YANG Fan, CUI Ming. Urban forest community structure and correlation analysis on the plant diversity index in Haikou City, China [J]. J Zhejiang A&F Univ, 2019, 36(6): 1142 − 1150. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20190396
 点击查看大图
点击查看大图
计量
- 文章访问数: 2720
- HTML全文浏览量: 845
- PDF下载量: 90
- 被引次数: 0

