





-
露天矿在开采过程中形成的排土场,由松散的土石混合剥离物构成,一般占矿山用地的一半以上[1]。中国的露天矿大多位于干旱少雨区域[2],在大风天气易产生大量沙(粉)尘[3],是当地大气污染的主要来源之一[4],危害人体健康和周边环境,因此修复排土场迫在眉睫[5-6]。研究表明:植物可以有效地减少大气中颗粒物的含量[7]。与非生物材料相比,植物独特的形态结构具备吸着、黏附颗粒物的良好条件,并可随生长更新叶片,提高滞尘能力[8]。植物在漫长的进化和发展过程中,与环境相互作用,逐渐形成了许多内在生理和外在形态方面的适应策略,这些适应策略的表现即为植物功能性状[9]。一方面,植物对环境变化产生响应(响应性状);另一方面,植物对生态系统功能产生影响(影响性状)[10]。基于此,在选择植物种时,既要考虑植物的适生性,也要使形成的植物群落发挥重要的生态功能,比如涵养水源、保持土壤、滞沙滞尘等[11]。当前,国内外对植物滞尘能力的研究主要集中在城市绿化的乔灌木[12-13],而对于干旱荒漠区特别是露天矿开采区的草本植物滞尘功能研究较少。矿区排土场土壤肥力较差,氮、磷、有机质缺乏,制约植物的生长发育[14]。根据植物群落演替规律,复垦初期群落植被以1~2年生草本为主[15]。草本植物根系较密,在固着松散土壤颗粒[16],减少扬尘方面效果较好。关于露天矿排土场的植被恢复,许多学者进行了研究。实际工作中主要分为2类:一是参照历史[17],二是进行对比试验[18]。目前,国际上提出了一种新思路,即理论研究与植被恢复工作实际相结合,通过模型分析给出恢复策略[19]。基于生境过滤[20]、竞争理论[21]、质量比理论[22]等生态学理论,选择适宜的植物种及其配比,预测性相对较好,适合对环境条件发生显著变化的区域进行植被恢复。
本研究以内蒙古自治区乌海市新星煤矿排土场及其周边为研究区域,调查研究区地带性适生植被及沙(粉)尘特征,采用植物功能性状统计模型(Community Assembly by Trait Selection, CATS模型)进行分析[19, 23],结合生态学理论与植被修复实际工作,筛选出矿区抗逆适生的植物种,建立应对特定环境条件的优势滞尘植物功能性状配比,以发挥较好的滞尘效果,改善排土场及其周边的生态环境,以期为乌海市矿区环境治理提供科学依据,为今后理论研究运用于实践提供参考。
-
乌海市(38°35′44″~40°27′01″N, 105°57′15″~107°48′21″E)位于内蒙古自治区西部,地处黄河上游,东、北与鄂尔多斯市相接,西邻阿拉善盟,南部与宁夏回族自治区石嘴山市毗连。全市地势东西高、中间低,南高北低,处在贺兰山脉北部余端,是南北走向的桌子山、甘德尔山、五虎山之间形成的狭长平坦谷地。乌海市处于干旱荒漠区,属于典型的温带大陆性气候,夏季高温少雨,冬季寒冷少雪,春秋干旱多风,昼夜温差大。年平均气温为9.8 ℃,年平均降水量为157.9 mm,年平均蒸发量为3249.0 mm[24],多年平均湿度为42%,年平均风速为2.7 m·s−1。该地区的植被类型以荒漠植被为主,其中草本主要有针茅Stipa capillata、猪毛菜Salsola collina、骆驼蓬Peganum harmala、火煤草Olgaea leucophylla、沙蒿Artemisia desertorum、雾冰藜Bassia dasyphylla等。
选择乌海市新星煤矿4号排土场作为本研究试验场地。4号排土场位于进矿道路东侧,南北走向,坡度38°,共6个台阶。由下往上第1~4台阶坡高为25 m,第5台阶坡高为28 m,第6台阶正在排土。第1台阶和第2台阶采用干砌石护坡,第3台阶和第4台阶主要采用人工覆土、撒播草籽、草帘覆盖和菱形沙柳Salix cheilophila沙障等边坡防护措施,覆土深度不足50 cm,并配置喷灌系统;第5台阶和第6台阶未进行治理[24]。已治理坡面主要植物有柠条锦鸡儿Caragana korshinskii、紫苜蓿Medicago sativa、黑沙蒿Artemisia ordosica、细枝山竹子Hedysarum scoparium等,未治理坡面分布的植物主要是沙蓬Agriophyllum squarrosum、雾冰藜、白茎盐生草Halogeton arachnoideus等。
-
为深入了解研究区植物的种类和功能性状,于2020年8月,在排土场周围根据不同植物群落组成共选择14个5 m×5 m植物样地进行调查。为减少降雨对植物叶片冲刷,影响叶片滞尘量的测定,调查均在降雨8 d后进行。每个样地随机选取3个1 m×1 m的草本样方,总计42个草本样方。记录每个样地的坐标位置,统计每个样方中的植物种及其株数,同时获取植物叶片和完整根系。
-
同一样地的每种植物随机选取3株生长良好、无病害的健康植株作为样本,用剪刀小心剪下健康完整的叶片若干,装入干净的自封袋中并牢固封口。每种植物重复3次,带回实验室4 ℃保存。采集过程均佩戴聚乙烯手套,减少抖动,避免叶片上尘土掉落。
-
采集被选的3株健康植株的叶片后,用铁锹小心挖取植株的完整根系,轻轻抖落浮土,装入标本夹,带回实验室4 ℃保存,待测。
-
为减少降雨对土壤水分测定的影响,均在降雨8 d后测定土壤含水量。在排土场未治理坡面设置1个样地,随机选取3个1 m×1 m排土场样方。对42个草本样方和3个排土场样方按五点梅花法用土钻钻取0~30 cm的土壤样品,带回室内用烘干法进行土壤水分测定。
-
2020年8月,采用流动监测法,使用Dustmate便携式粉尘监测仪测定14个草本样地(C草本)和1个排土场样地(C排土场)的总悬浮颗粒物浓度。每个监测点每天早晚各监测1次,1 min采集1次数据,共采集6次,取每日6次平均值作为该监测点的总悬浮颗粒物浓度背景值。
-
植物单位叶面积在单位时间内滞留的粉尘量可用于评价其滞尘能力[25]。本研究采用水洗过滤差重法[26]测定所调查的植物种的单位叶面积滞尘量(D)。将采集的叶片放入盛有蒸馏水的烧杯中浸泡,并用软毛刷小心清洗叶片正、反面的粉尘,用蒸馏水冲洗叶片后晾干备用。将所有清洗液用已知质量的0.15 μm孔径的滤膜(上海新亚混合纤维微孔滤膜)过滤。过滤后的滤膜放入60 ℃烘箱(Binder FED53)烘干至恒量,再用万分之一天平称量,求得滤膜过滤前后质量之差即为植物叶片滞尘量(W)。清洗晾干后的叶片经扫描仪(Epson 600)扫描后,用Photoshop 7.0软件计算叶面积(S),求得植物单位叶面积滞尘量(D):
$D=W/S。 $ -
将采集的根系用蒸馏水小心洗去泥沙,置于根系盘并使之充分舒展,用扫描仪(Epson V850 Pro)扫描得到图像,经配置的Scanner.cal校准文件处理后,用WinRHIZO-Pro根系分析系统分析根系体积。
-
由于植物分布区域不同,大气粉尘浓度背景值影响植物滞尘过程,因此,需要对实测单位叶面积滞尘量(D实测)进行校正,通过计算模拟同一环境条件下各物种的单位叶面积滞尘量,即标准单位叶面积滞尘量(D标准)。校正公式为:
$D_{标准}=D_{实测} \times (C_{排土场}/C_{草本})。 $ -
通过环境过滤理论可得到用于植被恢复的物种组合,1个功能性状目标值可以得到1种群落组合[19]。为了确定未来植被恢复群落的功能性状目标,需要根据环境条件对群落平均性状(CWM,表示群落内所测物种某一性状值与其相对生物量积的总和[27])进行回归,从而根据待恢复区域的环境条件(如土壤水分)获得最优性状值(功能性状目标值
$ {{\bar T}_{k}} $ )。$\displaystyle {C_{{\rm{WM}}}} = \sum\limits_{i = 1}^S {{p_i} {t_i}} 。$ 其中:S为物种数,pi为第i物种的相对多度,ti为第i物种的平均性状值。 -
采用广义线性模型对植物样地的根体积与其相对生物量积的总和(CWM)和土壤水分进行线性回归和相关性分析
-
CATS模型是由最大熵(Maxent)模型转化而来。该模型简化了种群动态变化的复杂性,可从物种库中筛选出适合的植物种,使得在给定环境中以某种相对多度和某些植物功能性状决定群落组成[28]。该模型基于物种和群落水平的功能性状来预测区域中每个物种的相对多度分布。当得到待恢复区域的功能目标值时,CATS模型就可以合理地预测物种的相对多度[19]。该模型的数学公式如下:
$\displaystyle \sum\limits_{i = 1}^S {{t_{ik}}{p_i} = {{\bar T}_k}}$ ;$\displaystyle \sum\limits_{i = 1}^S {{p_i} = 1}$ ;${p}_{i} > 0 $ 。其中:tik为第i个物种的第k个性状;pi为物种i相对多度;$ {\bar{T}_{k}} $ 为功能性状目标值;S为物种数。通常情况下,性状数少于物种数,方程组可能出现多解,即欠定线性方程组。采用R包limSolve求解约束线性方程,“xsample”函数采用马尔可夫链蒙特卡洛(MCMC,迭代次数iter设置为2 000)算法对解空间进行采样。用线性代数方法,将CATS模型改写成Ax=b矩阵,其中A为系数矩阵(即物种平均性状),x为未知物种相对多度矩阵,b为常数(即性状约束值)。若约束值一致(1个解或者无穷多个解存在),可从x得到统一的样本,即得到满足功能性状约束条件的物种相对多度的范围。
-
根据植物样方调查的植物种,选择适合生长的乡土植物,如表(1)所示。
-
选择合理的功能性状目标值(
$ {{\bar T}_{k}} $ ),这些目标值应落在物种库中功能性状值的范围内。对于植物根体积,根据排土场土壤水分含量,结合根体积的CWM与土壤水分的线性回归关系,模拟得到未来植物群落的根体积性状目标值($ {{\bar T}_{k1}} $ )。对于植物单位叶面积滞尘量,从各植物种的标准单位叶面积滞尘量中选择较高的数值作为未来植物群落的单位叶面积滞尘量性状目标值($ {{\bar T}_{k2}} $ )。 -
通过对14个植物样地的调查,选取了22种乡土植物种,并且对每种植物的单位叶面积滞尘量和根体积进行测定。由表1可知:根体积与单位叶面积滞尘量呈反比关系。其中猪毛菜、燥原芥Ptilotrichum cancscens、阿尔泰狗娃花Aster altaicus均表现为单位叶面积滞尘量较高、根体积较小;与上述3种植物相反,防风Saposhnikovia divaricata、骆驼蓬、戈壁天门冬Asparagus gobicus表现出单位叶面积滞尘量较少、根体积较大;驴欺口Echinops latifolius、冷蒿Artemisia frigida、沙葱Allium mongolicum的单位叶面积滞尘量和根体积分别为2.00 g·m−2和0.30 cm3,表现出单位叶面积滞尘量和根体积均较小;其余物种的单位叶面积滞尘量和根体积分别为5.00~35.00 g·m−2和0.50~5.80 cm3。
表 1 植物种功能性状值
Table 1. Functional trait values of plant species
编号 植物种名 平均根体积/cm3 平均单位叶面积滞尘量/
(g·m−2)编号 植物种名 平均根体积/cm3 平均单位叶面积滞尘量/
(g·m−2)1 猪毛菜 0.65 35.36 12 沙葱 0.57 2.34 2 燥原芥 0.21 32.17 13 苦豆子 0.80 1.99 3 阿尔泰狗娃花 0.97 32.17 14 荒漠黄耆 0.56 1.81 4 雾冰藜 1.10 19.91 15 防风 5.92 1.52 5 蝎虎驼蹄瓣 0.36 15.78 16 戈壁天门冬 5.46 1.43 6 刺沙蓬 0.59 11.98 17 驴欺口 0.19 1.20 7 隐子草 2.90 8.10 18 火煤草 1.35 1.20 8 砂珍芨豆 1.35 6.30 19 骆驼蒿 0.54 1.04 9 骆驼蓬 5.85 6.14 20 沙鞭 1.88 0.77 10 细枝岩黄耆 0.22 5.01 21 针茅 3.11 0.60 11 冷蒿 0.14 2.98 22 沙蒿 2.74 0.35 说明:蝎虎驼蹄瓣Zygophyllum mucronatum、刺沙蓬Salsola ruthenica、隐子草Cleistogenes serotina、砂珍芨豆Oxytropis racemosa、 细枝岩黄耆Hedysarum scoparium、苦豆子Sophora alopecuroides、荒漠黄耆Astragalus grubovii、骆驼蒿Peganum nigellastrum、沙鞭Psammochloa villosa -
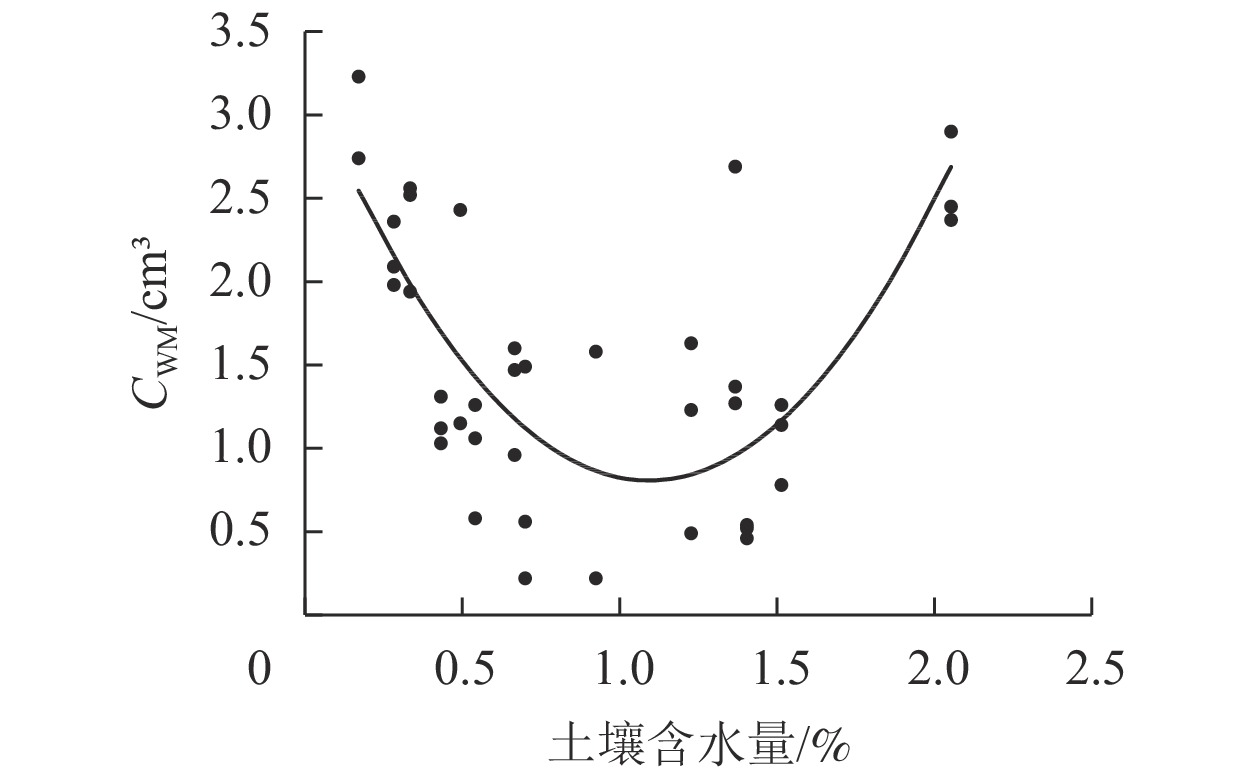
将42个草本样方的CWM与土壤含水量进行曲线估算,发现两者呈现“U”型曲线分布(图1),以土壤含水量1.1%为分界点,CWM随土壤含水量增加先减小后增大。由CWM(y)与土壤含水量(x)拟合得到二次函数关系式:y=20423x2−446.49x+3.2461 (P<0.05),将排土场土壤含水量代入方程计算,模拟得到未来植物群落的根体积性状目标值(
$ {{\bar T}_{k1}} $ ) =2.85 cm3。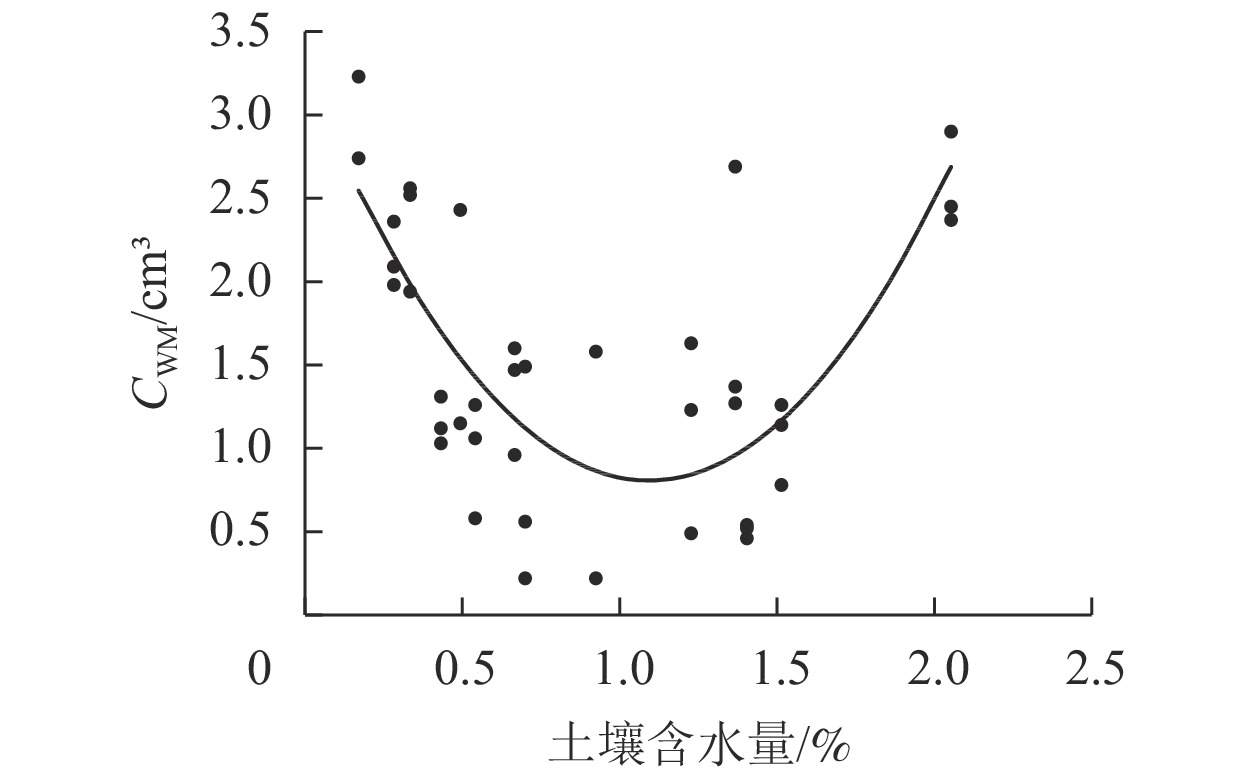
图 1 CWM与土壤含水量的关系
Figure 1. Relationship between CWM values of root volume and soil moisture content
-
根据模拟得到的未来植物群落的根体积性状目标值(
$ {{\bar T}_{k1}} $ )和设定的单位叶面积滞尘量性状目标值($ {{\bar T}_{k2}} $ ),以及22种植物的根体积和单位叶面积滞尘量,使用CATS模型得到的各植物种相对多度(图2)。由图2可见:猪毛菜和骆驼蓬的平均相对多度较高,中位数分别为0.41和0.32,阿尔泰狗娃花、防风、燥原芥、戈壁天门冬的平均相对多度分别为0.05、0.04、0.01和0.01。其他物种相对多度为0。这表明:在研究区的环境条件下,猪毛菜适合作为群落的优势物种,骆驼蓬适合作为群落的亚优势物种,其他物种不适合作为群落的优势物种。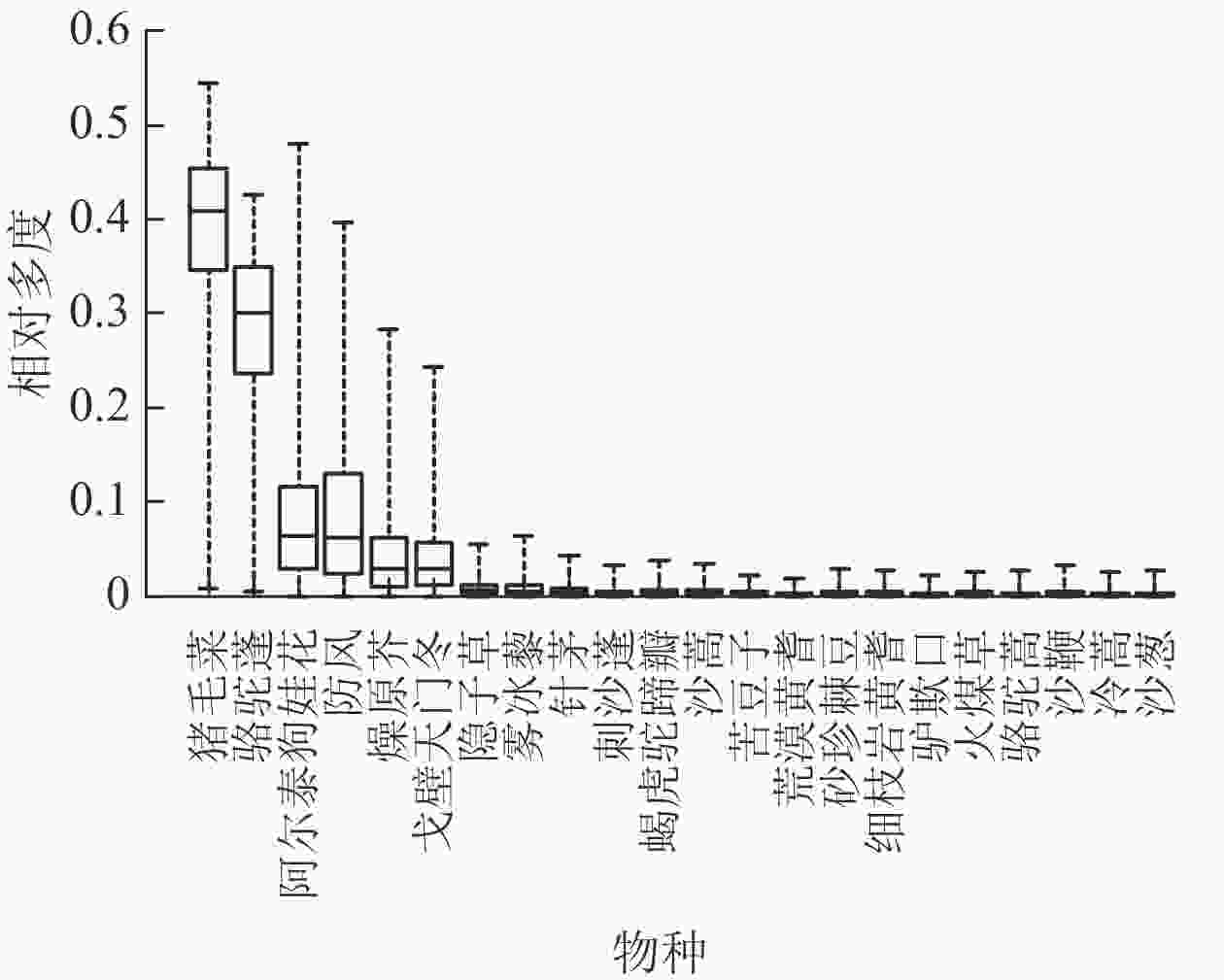
图 2 植物种相对多度分布
Figure 2. Relatively abundant distribution of plant species
-
植物种的选择是植被恢复过程中的重要环节[29]。在干旱荒漠区露天煤矿排土场进行植被重建能否成功很大程度上取决于植物种类的选择,排土场首要的环境限制因子是土壤水分。植物形成了多种生理生态特征以应对干旱胁迫,特别是增大根茎比。植物根系中许多功能性状与土壤含水量有关,极端干旱环境下白梭梭Haloxylon persicum的根系生物量和根长与土壤含水量呈极显著正相关[30];白刺Nitraria tangutorum中根系生物量和细根生物量与土壤水分呈显著相关[31];本研究对根体积CWM与土壤含水量进行曲线估算,两者成“U”型曲线分布。当土壤含水量少于1.1%时,土壤含水量与根体积CWM是负相关关系,这可能是植物为适应干旱环境的一种响应。韩锦峰等[32]研究发现:在烤烟Nicotiana tabacum伸根期,轻度土壤干旱条件下根系体积增长迅速,干物质积累量大。席璐璐等[33]研究发现:1年生草本植物在轻度干旱胁迫时,会增加根系活力,保证根系从土壤中吸收更多水分,以缓解干旱对生长的抑制。而在重度干旱胁迫时,茎长明显下降。植物通过延伸主根系长度、增加根长来适应严重干旱。本研究中,当土壤含水量大于1.1%时,土壤含水量与根体积CWM是正相关关系。这可能是因为当土壤含水量增加,植物根系通过增大根体积以便于更好地吸收水分。要更加全面地研究干旱区植物的适应策略,还需要对植物诸多性状进行监测。
排土场植被恢复的另一目标是滞尘。本研究排土场处于干旱荒漠区,其所在的乌海市,形成了以工矿业为主的单一化产业结构,容易产生大量粉尘[34],对环境和人体产生强烈的负面影响。本研究测定了22种草本植物单位叶面积的滞尘量,高值与低值之间差异明显,分析其原因可能有2个方面:①不同植物距离地面的高度不同,大气颗粒物浓度随高度分布存在差异。②不同植物叶表面微结构差异较大,对不同大气颗粒物的滞留能力存在差异。另外,大气颗粒物之间的质量存在差异。植物滞留空气颗粒物受到多方面因素的影响,而植物种类和叶表面形态对于大气颗粒物的滞留具有重要的影响作用。今后研究不同植物种的滞尘功能,还需观测植物种的其他性状,特别是植物叶面微结构特征与其滞尘能力之间的关系,有利于科学合理选择优势滞尘植物种。
植被恢复常用2类方法,其一是从历史视角,参照过去某一环境条件下植物群落的组成结构确定配置方式[35],该方法的风险在于过去与未来的环境条件可能差异巨大,得到的植被恢复策略不能保证适用于未来的环境条件。其二是根据立地条件、生态功能或用途进行恢复,对不同植物种进行对比试验[36],选择最适合当下环境和需求的植物种进行群落配置,筛选出适宜植物种,模拟群落的时空结构,作为待恢复区域的植被恢复策略。杨修等[35]对德兴铜矿1号尾矿库的立地条件进行调查,发现植被恢复的主要限制因子是土壤条件,又经过立地试验对15种草种进行优劣比较,发现最优的适生先锋草种为水蜡烛Dysophylla yatabeana。也有研究者利用集成地统计学、点格局分析和空间比较方法[37],对乌海市的灌木与土壤养分的空间格局关系进行量化,以筛选得到适生植物种,但该方法适合用于灌木和乔木研究,对于密生草本难以得到空间格局关系。CATS模型联系理论和实践,基于植物功能性状,通过植物对环境的响应与影响,预测未来环境条件下的群落丰富度,得到在排土场适合种植猪毛菜和骆驼蓬。猪毛菜是1年生草本,叶片具表皮毛可降低蒸腾作用,具有发达的薄壁储水组织提高水分蓄存能力,增加根冠比适应干旱环境[38]。骆驼蓬为蒺藜科Zygophyllaceae骆驼蓬属Peganum多年生草本,主要分布在中国西北部的干旱半干旱地区,是荒漠植被的重要组成成分之一,具有发达的根系和很强的耐盐耐旱抗寒特性[39]。有研究表明:骆驼蓬通过降低比叶面积[40],产生较厚的蜡质角质层、发达的栅栏组织和排列紧密的海绵组织,减弱蒸腾应对干旱胁迫,因此,猪毛菜和骆驼蓬能够适应排土场干旱少雨的环境。但这仅仅是理论研究结果,还没有经过实践检验。如果引入更多的植物功能性状,在实践中不断修正CATS模型参数,那么研究结果更具有统计学意义,更有利于指导植被修复工作。
本研究通过调查内蒙古自治区乌海市新星煤矿排土场及其周边的适生性草本植物,将植物的滞尘功能和适生性用于筛选优势滞尘植物,经CATS模型模拟得到配比,将植物功能性状的滞尘作用从物种水平推至群落尺度。本研究建议以猪毛菜和骆驼蓬的相对多度比6∶4进行播种,作为研究区的植被恢复策略。
Ratio of dominant dust retaining plants in waste dump of desert open-pit coal mine based on CATS model
-
摘要:
目的 干旱荒漠区露天矿排土场易产生沙(粉)尘,造成大气污染。基于植物功能性状模型(Community Assembly by Trait Selection,CATS模型)模拟,得到研究区排土场优势滞尘植物及物种在恢复群落中的配比,提出植被恢复策略。 方法 于2020年8月在内蒙古自治区乌海市新星煤矿排土场周围进行样方调查,测定物种水平和群落水平的性状值,并计算功能性状目标值,将植物滞尘功能和根体积作为模型的2个因子代入模型计算。 结果 植物种的根体积与单位叶面积滞尘量呈反比;群落根体积与土壤含水量呈现“U”型曲线分布;模型模拟结果表明:各物种相对多度最高的是猪毛菜Salsola collina和骆驼蓬Peganum harmala,相对多度的中位数分别为0.41和0.32。 结论 在研究区排土场以猪毛菜和骆驼蓬的相对多度比6∶4的比例播种,可发挥相当的滞尘效益,能够很好地解释植被修复过程中的一些机制性问题,为其他研究提供借鉴。图2表1参40 Abstract:Objective The dump of open-pit mine in arid desert regionis easy to produces and (powder) dust and cause air pollution. The purpose of this study is to obtain the ratio of dominant dust retaining plants and species in the restoration community based on the simulation of Community Assembly by Trait Selection (CATS) model, and put forward the vegetation restoration strategy. Method A quadrat survey was conducted around the waste dump of Wuhai Xinxing Coal Mine in Inner Mongolia Autonomous Regionin August 2020 to determine the trait values at species and community levels, calculate the target values of functional traits, and substitute the dust retention function and root volume of plants into the model as two factors. Result The root volume of plant species was inversely proportional to the dust retention per unit leaf area. The root volume and soil water content was U-shaped. The simulation results showed that the highest relative abundance of each species was Salsola collina and Peganum harmala, with amedian of 0.41 and 0.32, respectively. Conclusion To plant S. collina and P. harmala in the dumping site with the relative abundance ratio of 6 to 4 is beneficial to dust detention, which can also explain some mechanical problems in vegetation restoration and provide reference for other studies. [Ch, 2 fig. 1 tab. 40 ref.] -
Key words:
- CATS model /
- waste dump of mining area /
- plant functional traits /
- ratio of plant
-
图 1 CWM与土壤含水量的关系
Figure 1 Relationship between CWM values of root volume and soil moisture content
表 1 植物种功能性状值
Table 1. Functional trait values of plant species
编号 植物种名 平均根体积/cm3 平均单位叶面积滞尘量/
(g·m−2)编号 植物种名 平均根体积/cm3 平均单位叶面积滞尘量/
(g·m−2)1 猪毛菜 0.65 35.36 12 沙葱 0.57 2.34 2 燥原芥 0.21 32.17 13 苦豆子 0.80 1.99 3 阿尔泰狗娃花 0.97 32.17 14 荒漠黄耆 0.56 1.81 4 雾冰藜 1.10 19.91 15 防风 5.92 1.52 5 蝎虎驼蹄瓣 0.36 15.78 16 戈壁天门冬 5.46 1.43 6 刺沙蓬 0.59 11.98 17 驴欺口 0.19 1.20 7 隐子草 2.90 8.10 18 火煤草 1.35 1.20 8 砂珍芨豆 1.35 6.30 19 骆驼蒿 0.54 1.04 9 骆驼蓬 5.85 6.14 20 沙鞭 1.88 0.77 10 细枝岩黄耆 0.22 5.01 21 针茅 3.11 0.60 11 冷蒿 0.14 2.98 22 沙蒿 2.74 0.35 说明:蝎虎驼蹄瓣Zygophyllum mucronatum、刺沙蓬Salsola ruthenica、隐子草Cleistogenes serotina、砂珍芨豆Oxytropis racemosa、 细枝岩黄耆Hedysarum scoparium、苦豆子Sophora alopecuroides、荒漠黄耆Astragalus grubovii、骆驼蒿Peganum nigellastrum、沙鞭Psammochloa villosa  下载: 导出CSV
下载: 导出CSV
-
[1] 郭义强, 罗明, 王军. 中德典型露天煤矿排土场土地复垦技术对比研究[J]. 中国矿业, 2016, 25(2): 63 − 68. GUO Yiqiang, LUO Ming, WANG Jun. Study on comparison of land reclamation technology in typical surface mine dump between China and Germany [J]. China Min Mag, 2016, 25(2): 63 − 68. [2] 吕凯, 李雪飞, 智颖飙. 露天煤矿排土场生物修复与生态重建技术[J]. 内蒙古师范大学学报(自然科学汉文版), 2019, 48(5): 458 − 464. LÜ Kai, LI Xuefei, ZHI Yingbiao. Mechanism of phytoremediation and co-reconstruction for waste dump in open-cast collieries [J]. J Inner Mongolia Norm Univ Nat Sci Ed, 2019, 48(5): 458 − 464. [3] 刘韵, 王若水, 张艳, 等. 春季黄河附近乌海市露天煤矿大气不同粒径粉尘质量浓度分布规律[J]. 中国水土保持科学, 2020, 18(3): 1 − 11. LIU Yun, WANG Ruoshui, ZHANG Yan, et al. Dust concentration distribution patterns of different particulate matter in atmosphere in a surface coal mine of Wuhai City near the Yellow River during spring [J]. Sci Soil Water Conserv, 2020, 18(3): 1 − 11. [4] 陈晓艳, 白洁冰. 乌海市区大气污染特征及防治对策研究[J]. 环境与发展, 2014, 26(3): 31 − 33. CHEN Xiaoyan, BAI Jiebing. Study on the air pollution characteristics of Wuhai urban dis trict and its coutermeasures for control [J]. Environ Dev, 2014, 26(3): 31 − 33. [5] 关军洪, 郝培尧, 董丽, 等. 矿山废弃地生态修复研究进展[J]. 生态科学, 2017, 36(2): 193 − 200. GUAN Junhong, HAO Peiyao, DONG Li, et al. Review on ecological restoration of mine wasteland [J]. Ecol Sci, 2017, 36(2): 193 − 200. [6] 韩涛, 梁旭燕. 煤矿矿区大气污染现状及对策[J]. 能源环境保护, 1994, 8(4): 24 − 26. HAN Tao, LIANG Xuyan. Present situation and countermeasure of air pollution in coal mine area [J]. Energy Environ Prot, 1994, 8(4): 24 − 26. [7] BECKETT K P, FREER-SMITH P H, TAYLOR G. Effective tree species for local air quality management [J]. J Arboric, 2000, 26(1): 12 − 19. [8] 柴一新, 祝宁, 韩焕金. 城市绿化树种的滞尘效应: 以哈尔滨市为例[J]. 应用生态学报, 2002, 13(9): 1121 − 1126. CHAI Yixin, ZHU Ning, HAN Huanjin. Dust removal effect of urban tree species in Harbin [J]. Chin J Appl Ecol, 2002, 13(9): 1121 − 1126. [9] 刘晓娟, 马克平. 植物功能性状研究进展[J]. 中国科学: 生命科学, 2015, 45(4): 325 − 339. LIU Xiaojuan, MA Keping. Plant functional traits: concepts, applications and future directions [J]. Sci Sin Vitae, 2015, 45(4): 325 − 339. [10] KLUMPP K, SOUSSANA J F. Using functional traits to predict grassland ecosystem change: a mathematical test of the response-and-effect trait approach [J]. Global Change Biol, 2010, 15(12): 2921 − 2934. [11] 彭少麟. 恢复生态学与退化生态系统的恢复[J]. 中国科学院院刊, 2000(3): 188 − 192. PENG Shaolin. Restoration ecology and restoration of degraded ecosystems [J]. Bull Chin Acad Sci, 2000(3): 188 − 192. [12] 廖莉团, 苏欣, 李小龙, 等. 城市绿化植物滞尘效益及滞尘影响因素研究概述[J]. 森林工程, 2014, 30(2): 21 − 24. LIAO Lituan, SU Xin, LI Xiaolong, et al. Review on the purification effects of urban landscape plants and factors affecting detaining dust [J]. For Eng, 2014, 30(2): 21 − 24. [13] JANHÄLL S. Review on urban vegetation and particle air pollution: deposition and dispersion [J]. Atmos Environ, 2015, 105: 130 − 137. [14] 王丽丽, 甄庆, 王颖, 等. 晋陕蒙矿区排土场不同改良模式下土壤养分效应研究[J]. 土壤学报, 2018, 55(6): 1525 − 1533. WANG Lili, ZHEN Qing, WANG Ying, et al. Effect of soil amelioration on soil nutrients at mining dumps in the Shanxi-Shaanxi-Inner Mongolia Region [J]. Acta Pedol Sin, 2018, 55(6): 1525 − 1533. [15] NOVÁK J, PRACH K. Vegetation succession in basalt quarries: pattern on a landscape scale [J]. Appl Veg Sci, 2003, 6(2): 111 − 116. [16] 任蔓莉, 魏晨辉, 裴忠雪, 等. 松嫩平原沙土区不同植被类型对土壤相关指标的影响[J]. 植物研究, 2015, 35(5): 765 − 771. REN Manli, WEI Chenhui, PEI Zhongxue, et al. Influences of different vegetation types on soil parameters in degraded sandy lands of Songnen Plain [J]. Bull Bot Res, 2015, 35(5): 765 − 771. [17] 原野, 赵中秋, 白中科, 等. 露天煤矿复垦生物多样性恢复技术体系与方法: 以平朔矿排土场为例[J]. 中国矿业, 2017, 26(8): 93 − 98. YUAN Ye, ZHAO Zhongqiu, BAI Zhongke, et al. Technology system and method of biodiversity restoration for the reclamation of opencast coal mine: a case study from the dumps in Pingshuo mine [J]. China Min Mag, 2017, 26(8): 93 − 98. [18] 李晋川, 王翔, 岳建英, 等. 安太堡露天矿植被恢复过程中土壤生态肥力评价[J]. 水土保持研究, 2015, 22(1): 66 − 71, 79. LI Jinchuan, WANG Xiang, YUE Jianying, et al. Evaluation on soil ecologic fertility during vegetation succession in Antaibao Open Pit [J]. Res Soil Water Conserv, 2015, 22(1): 66 − 71, 79. [19] LAUGHLIN D C. Applying trait-based models to achieve functional targets for theory-driven ecological restoration [J]. Ecol Lett, 2014, 17(7): 771 − 784. [20] JENTSCH A. The challenge to restore processes in face of nonlinear dynamics: on the crucial role of disturbance regimes [J]. Restoration Ecol, 2007, 15(2): 334 − 339. [21] KEDDY P A, SHIPLEY B. Competitive hierarchies in herbaceous plant communities [J]. Oikos, 1989, 54(2): 234 − 241. [22] GRIME J P. Benefits of plant diversity to ecosystems: immediate, filter and founder effects [J]. J Ecol, 1998, 86(6): 902 − 910. [23] LAUGHLIN D C, LAUGHLIN D E. Advances in modeling trait-based plant community assembly [J]. Trends Plant Sci, 2013, 18(10): 584 − 593. [24] 郭光. 内蒙古乌海市新星矿区露天煤矿生态修复治理规划研究[D]. 北京: 北京林业大学, 2020. GUO Guang. Study on Open-pit Coal Mine Ecological Restoration and Planning in Xinxing Mining Area, Wuhai City, Inner Mongolia[D]. Beijing: Beijing Forestry University, 2020. [25] 张灵艺, 秦华. 城市园林绿地滞尘研究进展及发展方向[J]. 中国园林, 2015, 31(1): 64 − 68. ZHANG Lingyi, QIN Hua. Research on progress of dust-retention for urban green space [J]. Chin Landscape Archit, 2015, 31(1): 64 − 68. [26] 岳晨, 李广德, 席本野, 等. 叶片大气颗粒物滞纳能力评估方法的定量对比[J]. 环境科学, 2021, 42(1): 114 − 126. YUE Chen, LI Guangde, XI Benye, et al. Quantitative comparison of methods to assess the airborne particulate matter retention capacity of leaves [J]. Environ Sci, 2021, 42(1): 114 − 126. [27] ACKERLY D D, CORNWELL W K. A trait-based approach to community assembly: partitioning of species trait values into within-and among-community components [J]. Ecol Lett, 2007, 10(2): 135 − 145. [28] SHIPLEY B, VILE D, GARNIER E. From plant traits to plant communities: a statistical mechanistic approach to biodiversity [J]. Science, 2006, 314(5800): 812 − 814. [29] 王金满, 杨睿璇, 白中科. 草原区露天煤矿排土场复垦土壤质量演替规律与模型[J]. 农业工程学报, 2012, 28(14): 229 − 235. WANG Jinman, YANG Ruixuan, BAI Zhongke. Succession law and model of reclaimed soil quality of opencast coal mine dump in grassland [J]. Trans Chin Soc Agric Eng, 2012, 28(14): 229 − 235. [30] 田起隆, 刘彤. 极端干旱环境下白梭梭细根分布与土壤水分关系[J]. 石河子大学学报, 2020, 38(1): 75 − 82. TIAN Qilong, LIU Tong. Relationship between the distribution characteristics of fine roots of Haloxylon persicum and soil moisture under extreme drought conditions [J]. J Shihezi Univ, 2020, 38(1): 75 − 82. [31] 李鸿儒, 王继和, 蒋志荣, 等. 白刺沙包发育过程的土壤水分与根系生物量的关系[J]. 甘肃农业大学学报, 2010, 45(6): 133 − 138. LI Hongru, WANG Jihe, JIANG Zhirong, et al. Relationship between soil moisture and root biomass during developmental process of Nitraria tangutorum nebkha [J]. J Gansu Agric Univ, 2010, 45(6): 133 − 138. [32] 韩锦峰, 汪耀富, 张新堂. 土壤水分对烤烟根系发育和根系活力的影响[J]. 中国烟草, 1992(3): 14 − 17. HAN Jinfeng, WANG Yaofu, ZHANG Xintang. Effects of soil moisture on root development and root activity of flue-cured tobacco [J]. China Tob, 1992(3): 14 − 17. [33] 席璐璐, 缑倩倩, 王国华, 等. 荒漠绿洲过渡带典型1年生草本植物对干旱胁迫的响应[J]. 生态学报, 2021, 41(13): 5425 − 5434. XI Lulu, GOU Qianqian, WANG Guohua, et al. The responses of typical annual herbaceous plants to drought stress in a desert-oasis ecotone [J]. Acta Ecol Sin, 2021, 41(13): 5425 − 5434. [34] 邓雅元, 赵廷宁, 张艳, 等. 基于SEM-EDS的荒漠草原区煤炭基地及其周边春季大气降尘特征及来源解析[J]. 中国环境科学, 2021, 41(12): 5512 − 5521. DENG Yayuan, ZHAO Tingning, ZHANG Yan, et al. Characterization and source appointment of atmospheric dust fall in Coal Base and surrounding areas in desert steppe based on SEM-EDS [J]. China Environ Sci, 2021, 41(12): 5512 − 5521. [35] 杨修, 高林. 德兴铜矿矿山废弃地植被恢复与重建研究[J]. 生态学报, 2001, 21(11): 1932 − 1940. YANG Xiu, GAO Lin. A study on re-vegetation in mining wasteland of Dexing Copper Mine, China [J]. Acta Ecol Sin, 2001, 21(11): 1932 − 1940. [36] 于泳, 杨伟, 李璐, 等. 弃渣场植被恢复与重建研究进展综述[J]. 亚热带水土保持, 2015, 27(3): 2 − 5, 28. YU Yong, YANG Wei, LI Lu, et al. Review on the research progress of vegetation restoration and reconstruction in slag disposal sites [J]. Subtrop Soil Water Conserv, 2015, 27(3): 2 − 5, 28. [37] HOU Jian, YANG Jianying, TAN Jin. A new method for revealing spatial relationships between shrubs and soil resources in arid regions[J/OL]. Catena, 2019, 183[2021-08-10]. doi. org/10.1016/j. catena. 2019.104187. [38] 彭磊, 张力, 周小龙, 等. 水分胁迫对新疆准东地区钠猪毛菜的生活史对策的影响[J]. 草业学报, 2021, 30(5): 65 − 74. PENG Lei, ZHANG Li, ZHOU Xiaolong, et al. Effects of water stress on life history strategy of Salsola nitrariain Zhundong, Xinjiang [J]. Acta Pratac Sin, 2021, 30(5): 65 − 74. [39] 任尚福. 南疆不同生境骆驼蓬叶片解剖结构的生态适应性[J]. 黑龙江农业科学, 2018(8): 19 − 23. REN Shangfu. Ecological adaptability of leaf anatomical structure of Peganum harmalain different habitats of Southern Xinjiang [J]. Heilongjiang Agric Sci, 2018(8): 19 − 23. [40] 郭新新, 左小安, 岳平, 等. 内蒙古荒漠草原沙生针茅、碱韭(Allium polyrhizum)和骆驼蓬叶形态性状对土壤水氮耦合的响应[J]. 中国沙漠, 2021, 41(1): 137 − 144. GUO Xinxin, ZUO Xiaoan, YUE Ping, et al. Responses of leaf morphological traits of three dominant plants to water and nitrogen in desert steppe of Inner Mongolia [J]. J Desert Res, 2021, 41(1): 137 − 144. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210604
 点击查看大图
点击查看大图
图(2) / 表(1)
计量
- 文章访问数: 1610
- HTML全文浏览量: 317
- PDF下载量: 31
- 被引次数: 0















