





-
在全球气候变暖背景下,由水分胁迫引起的树木死亡和作物减产等事件频发,水分胁迫带来的损失甚至超过其他环境胁迫所造成的损失总和[1]。研究表明:水力失效是导致植物死亡的主要原因[2]。研究植物的水力功能(水力效率和安全性)是量化植物抗旱能力的关键,而木质部结构是水力功能的基础,因此,结构和功能关系的定量研究是树木干旱胁迫研究的热点之一[3-4]。
导水率(Kh)和比导率(Ks)是衡量植物水力效率的重要参数[5],是植物为适应特定环境条件及物种竞争而形成的不同水力功能特征[6]。Ks越大,说明单位有效面积输送水分的能力越强,输水效率越高。此外,一般用导水率损失50%时的木质部水势(P50)来表征植物栓塞抗性,P50是确定木本植物抗旱极限的一个重要性状[7],不同类群植物的栓塞抗性差异很大[8]。有研究指出:水力效率与栓塞抗性之间可能存在一种权衡关系,即水力效率越大栓塞抗性就越小[9]。虽然水力效率-安全权衡存在与否仍有争议,但现有证据表明:在全球尺度上,效率和安全之间存在微弱的负相关[10-11]。
不同树种的材性、解剖等结构性状会影响树木的水分运输效率和水分利用策略[11-12]。按照次生木质部横截面上的导管管孔特征,双子叶植物可大致分为散孔材和环孔材2种功能类群[13-14]。在面对干旱诱导形成的栓塞产生的环境胁迫时,2种材性树种在水力结构上会进化不同的适应性策略。环孔材树种往往导管直径较大,具有较高的水分运输效率,但在水分胁迫条件下更容易遭受空穴化和栓塞胁迫[12]。而没有明显的早材和晚材区别的散孔材树种,它们较小的导管抵抗水分胁迫产生空穴化和栓塞的能力较强[5, 14]。除导管直径外,导管壁厚度、木材基本密度等对散孔材和环孔材树种水力效率和栓塞抗性都具有一定的影响[15-16]。COCHARD等[17]研究指出:导管壁厚度与栓塞抗性呈正相关,且厚度跨度比与栓塞抗性的相关性显著。此外,木质部在结构上形成了一个相互连通的网络管道结构,导管在木质部网络的空间排列影响了木质部的功能特性[18]。与散孔材树种相比,环孔材树种存在导管密度小、导管连接度低的特性,使其通过相邻导管或纹孔运输水分的概率显著小于散孔材树种,故而环孔材树种导管较散孔材更易发生栓塞[19]。因此,研究木质部结构与水力效率、栓塞抗性的关系,可揭示树木干旱致死机制。
目前,多年干旱和降水降低趋势可能是未来气候的开端[20],在这种气候背景下,物种的抗旱能力是生态系统稳定性与可持续发展的关键。河南省作为气候变化敏感区,自1961年以来,干旱发生频率和发生范围不断扩大[21],这对鸡公山国家级自然保护区内具有生态、经济和药用价值的园林树种的生长和生存产生潜在影响。基于此,本研究选取河南省鸡公山国家级自然保护区内园林树种中的3个散孔材和3个环孔材树种,对2种功能树种的枝条木质部解剖结构和水力功能性状差异进行分析,从植物水力结构的角度探讨植物的水分运输和抗旱性,以期为研究区园林植物的保护和开发提供指导意见。
-
研究样地位于河南省信阳市南部的鸡公山国家级自然保护区(31°46′~31°52′N,114°01′~114°06′E),该区地处北亚热带边缘,具有北亚热带向暖温带过渡的季风气候和山地气候特征。四季分明,光、热、水同期,年均气温为15.2 ℃,年均降水量为1 118.7 mm,植被以亚热带常绿阔叶林以及暖温带落叶阔叶林为主。保护区内森林茂密、生物资源丰富,为河南省物种最为丰富的区域之一。
-
选取鸡公山国家自然保护区2种材性的6个园林树种作为研究对象(表1),包括3个散孔材树种(枫香Liquidambar formosana、旱柳Salix matsudana、深山含笑Michelia maudiae)和3个环孔材树种(槲树Quercus dentata、楝Melia azedarach、野核桃Juglans cathayensis)。2019年6—9月,随机选取生长状况良好,株高、胸径、冠幅相似的植株,选择向阳生长的当年生枝条,所截枝条基部直径为6~8 mm,长为30~50 cm,每个树种取3~5株树,每株树4~6个重复,每个树种20个重复。
表 1 散孔材和环孔材样树的基本特性
Table 1. Basic characteristics of diffuse- and ring-porous sampled trees
材性 树种 树高/m 胸径/cm 树龄/a 冠幅/m 散孔材 枫香 17.45±0.35 16.26±0.12 15~20 4.10±0.22 旱柳 12.75±0.25 12.18±0.24 10~15 2.14±0.07 深山含笑 12.41±0.43 13.65±0.54 15~20 3.45±0.14 环孔材 槲树 13.32±0.25 13.34±0.27 15~20 5.86±0.08 楝 8.78±0.11 9.15±0.42 10~15 2.54±0.23 野核桃 11.33±0.17 13.42±0.15 15~20 4.02±0.21 说明:数值为平均值±标准误;样本数为20 株·种−1 -
将符合实验要求的枝条样品取下后,立刻浸入水中并用不透光容器保存带回。实验开始之前,将样品置于水中进行60~120 min的暗适应,之后在水下修剪样品,并在样品中间部位剪取一段约16~17 cm的茎段。将茎段基部表皮剥离,随后连接到木质部导水率与栓塞测量系统XYL’EM-Plus(Bronkhorst, Montigny-les-Cormeilles, 法国),用于测量导水率。首先用冲洗液(20 mmol·L−1氯化钾+1 mmol·L−1氯化钙)在低压下测得茎段的原位导水率(K0, kg·m·s−1·MPa−1),用0.15 MPa的压力冲刷枝条,直至枝条管胞内无气泡溢出为止,然后测量最大导水率(Kmax,kg·m·s−1·MPa−1)[22]。最大导水率测量结束后,将茎段放入压力腔中并保持茎段两端露出压力腔外,将压力表连在压力腔上[11]。在一定压力梯度下持续加压一定时间以诱导茎段发生栓塞,这个过程以0.2 或0.3 MPa的增量重复进行(取决于植物),直至导水率损失达90%以上,同时记录相应压力下的导水率(Kh, kg·m·s−1·MPa−1)。导水率损失百分比(PLC)计算如下:PLC=(1−Kh/Kmax)×100%。将所得压力值和PLC值在Origin软件中用sweibull分布函数进行拟合,获得脆弱曲线和导水率损失50%时对应的茎段木质部水势(P50)。比导率(Ks, kg·s−1·m−1·MPa−1)通过Kmax除以无髓、无树皮的基部边材横截面积得到。
-
从用于测定脆弱曲线的枝条上截取4段0.5 cm长的茎段。样品经固定、软化后采用番红固绿对染法制作石蜡切片。使用Leica DM 3000光学显微镜在50和400倍镜下摄像(图1)。用Image-J软件分析所摄照片,测量参数包括导管直径(μm)、导管壁厚度(μm)、导管密度(个·mm−2)以及导管壁厚度跨度比。
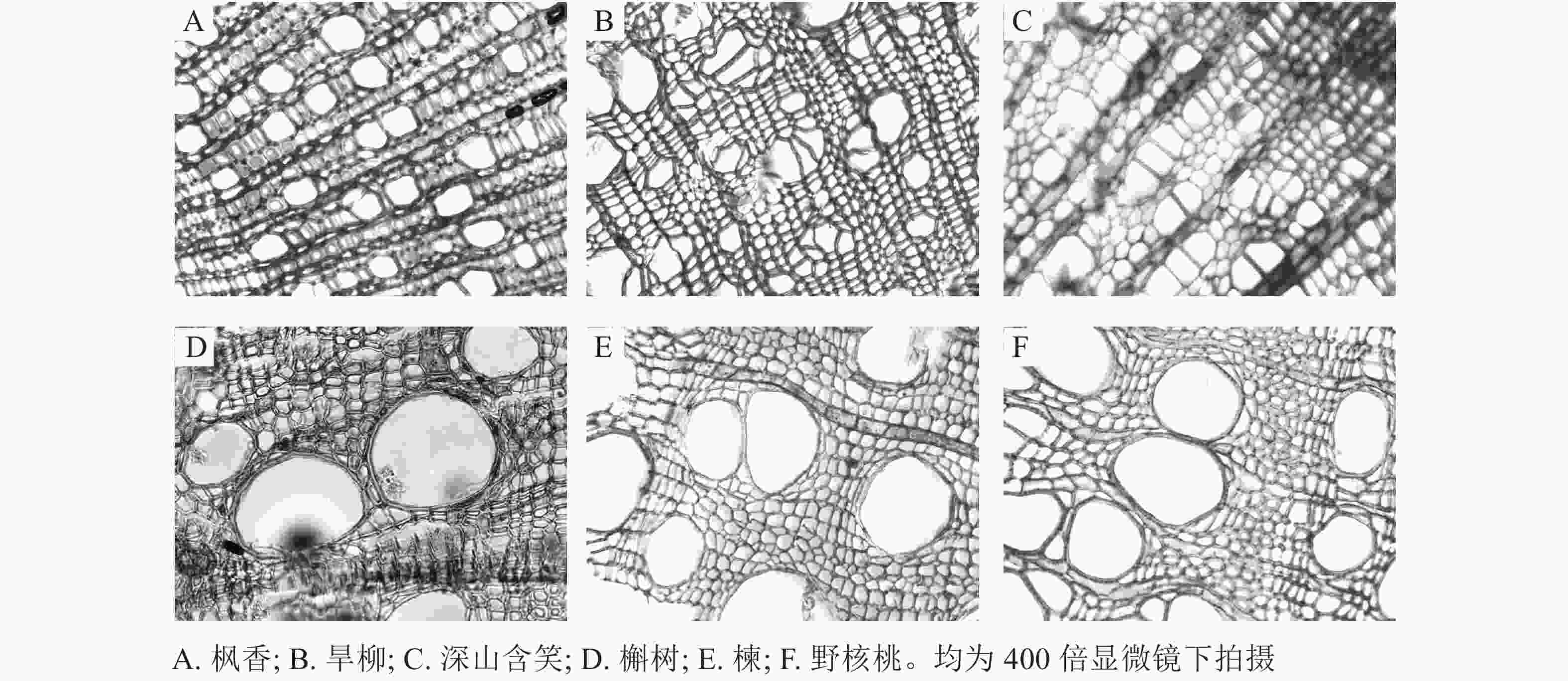
图 1 6个树种枝条木质部横切面照片
Figure 1. Photographs of cross-sections of xylem of 6 tree species
-
根据HACKE等[23]的方法测量样本的木材基本密度。将木材样品树皮用剃刀除去后浸没在蒸馏水中。采用排水法测定木材新鲜样品的体积(V,cm3)。然后将样品放置在75 ℃的烘箱烘48 h,测量干质量(W,g)。木材基本密度(WBD)计算公式为:WBD(g·cm−3)=W/V。
-
采用Minitab 18.1软件对数据进行单因素方差分析(one-way ANOVA), 显著性水平为0.05。采用Pearson相关分析检验各功能性状间的相关性。利用Origin软件作图。
-
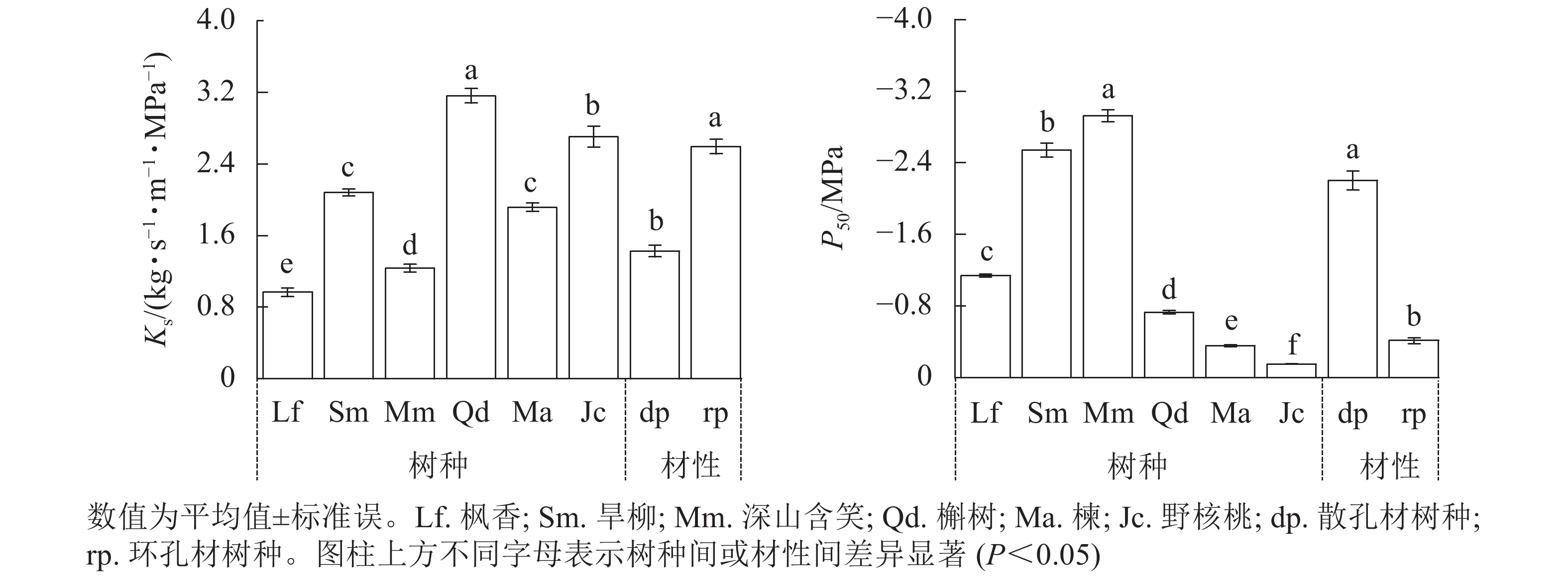
由图2可见:2种材性树种枝条比导率(Ks)和栓塞抗性(P50)差异均显著(P<0.05)。散孔材树种Ks为(1.43±0.07) kg·s−1·m−1·MPa−1,显著小于环孔材树种(P<0.05);散孔材树种P50为(−2.20±0.10) MPa,显著小于环孔材树种(P<0.05)。3个散孔材树种Ks均显著小于环孔材树种槲树和野核桃(P<0.05);3个散孔材树种P50均显著大于3个环孔材树种(P<0.05)。
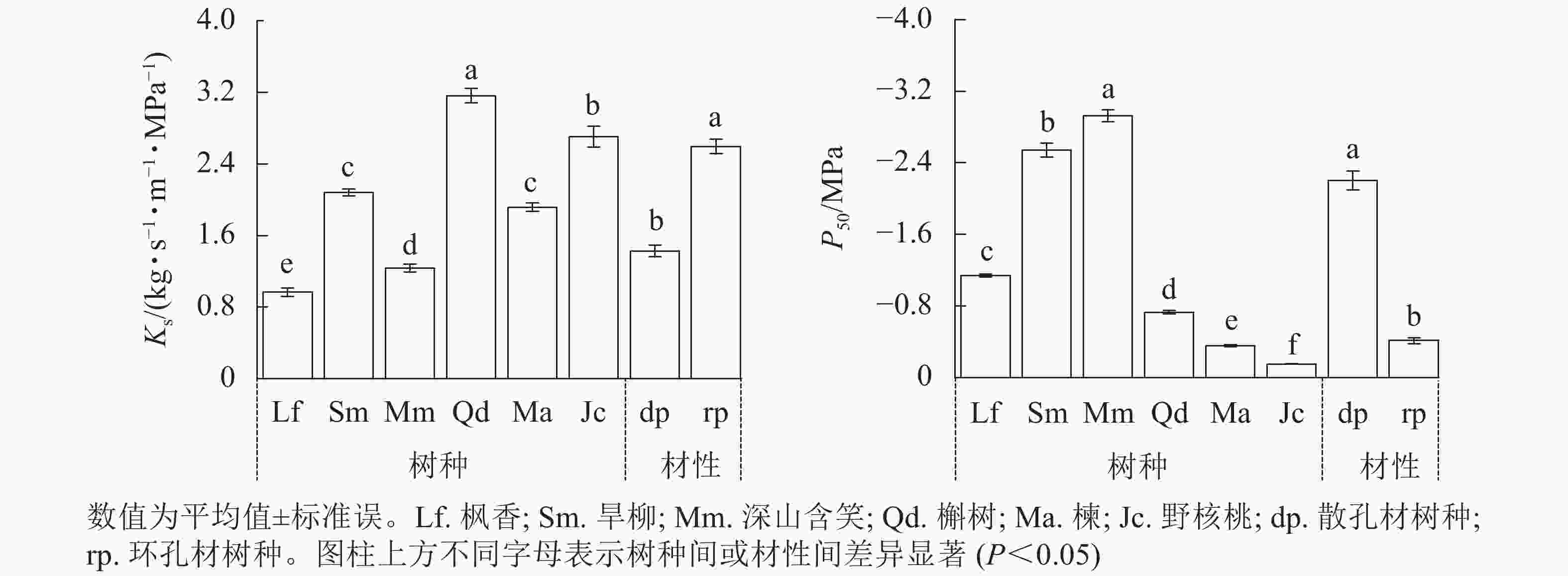
图 2 2种材性树种的比导率(Ks)与栓塞抗性(P50)
Figure 2. Comparison of specific sapwood conductivity (Ks) and cavitation resistance (P50) of tree species with two wood properties
-
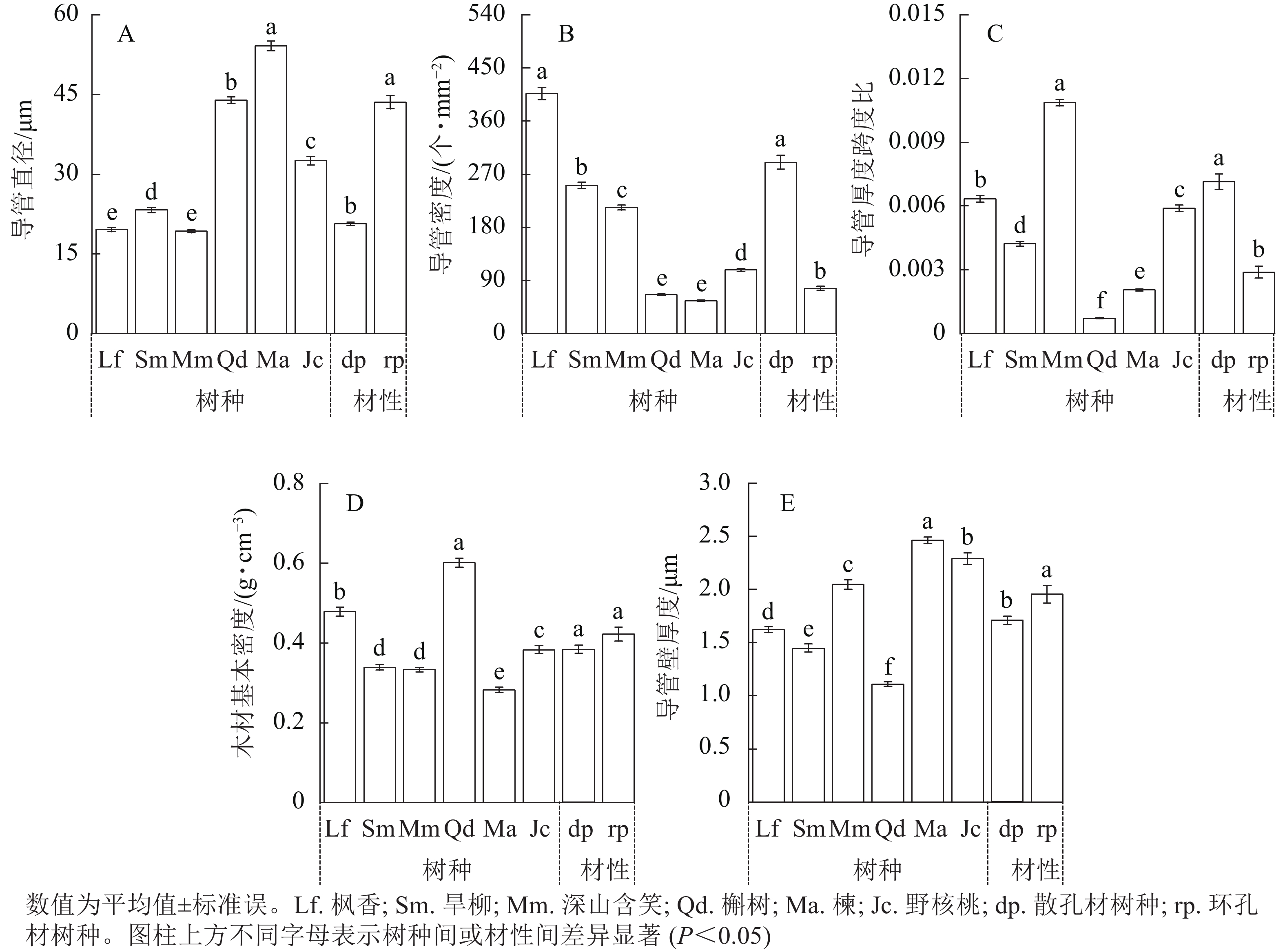
2种材性树种枝条木质部结构导管直径、导管密度、导管壁厚度跨度比和导管壁厚度均存在显著差异(图3,P<0.05)。散孔材树种平均导管直径显著小于环孔材树种(图3A,P<0.05),平均导管密度显著大于环孔材树种(图3B,P<0.05),平均导管壁厚度跨度比显著大于环孔材树种(图3C,P<0.05),平均木材基本密度无显著差异(图3D,P>0.05)。散孔材树种平均导管壁厚度显著小于环孔材树种(图3E,P<0.05),其中环孔材树种楝和野核桃的导管壁厚度显著大于其他4个树种(P<0.05)。
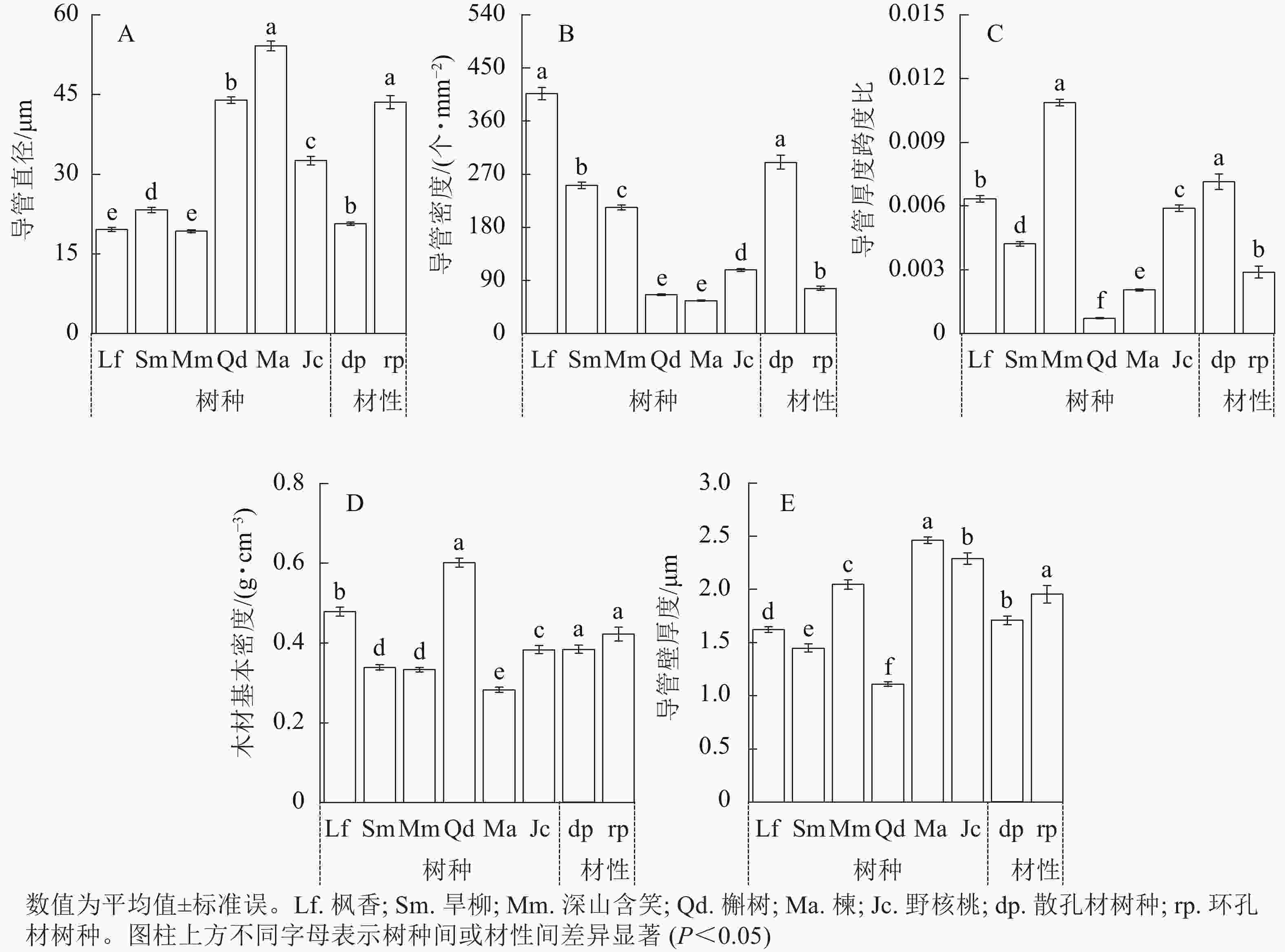
图 3 2种材性树种解剖结构性状
Figure 3. Comparison of the structural characteristics of two wood-based tree species
-
散孔材树种枫香(图4A)和旱柳(图4B)的比导率与栓塞抗性不相关,深山含笑的比导率与栓塞抗性呈显著负相关(P<0.05,图4C);环孔材树种楝的比导率与栓塞抗性呈显著正相关(P<0.05,图4D),槲树(图4E)和野核桃(图4F)的比导率与栓塞抗性不相关。6个树种中,仅散孔材树种深山含笑的比导率与栓塞抗性之间存在权衡,其他5个树种的比导率与栓塞抗性之间没有类似的权衡关系。

图 4 6个树种比导率(Ks)与栓塞抗性(P50)的关系
Figure 4. Relationship between specific conductance (Ks) and embolic resistance (P50) of six tree species
-
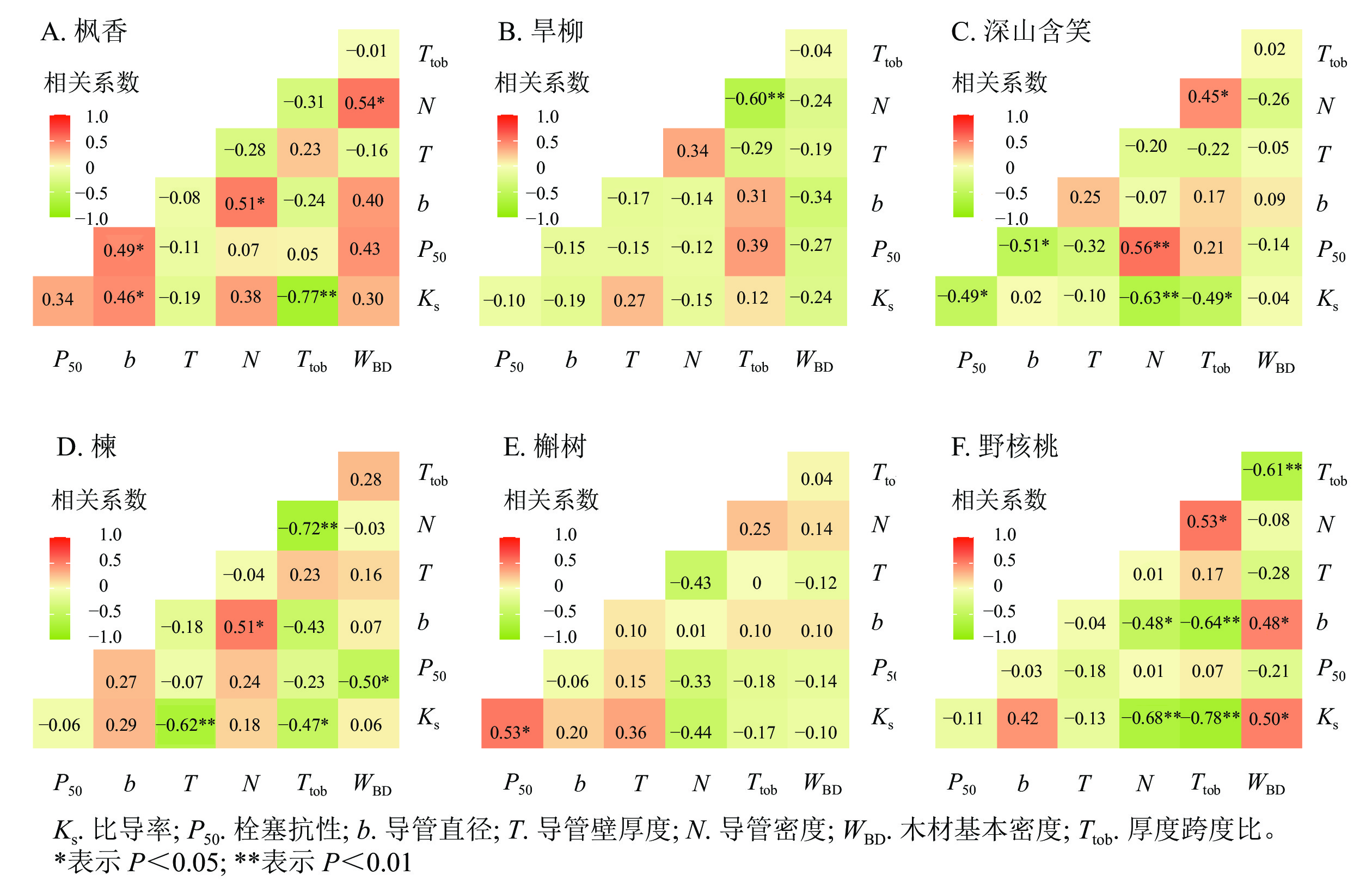
散孔材树种中,枫香的比导率与导管直径呈显著正相关(P<0.05),与导管壁厚度跨度比呈极显著负相关(P<0.01);枫香的栓塞抗性与导管直径呈显著正相关(P<0.05,图5A)。旱柳木质部结构与水力功能之间不存在显著相关(图5B)。深山含笑的比导率与导管壁厚度跨度比呈显著负相关(P<0.05),与导管密度呈极显著负相关(P<0.01);深山含笑的栓塞抗性与导管直径呈显著负相关(P<0.05),与导管密度呈极显著正相关(P<0.01,图5C)。
环孔材树种中,槲树的比导率与导管壁厚度呈极显著负相关(P<0.01),与导管壁厚度跨度比呈显著负相关(P<0.05);槲树的栓塞抗性与木材基本密度呈显著负相关(P<0.05,图5D)。楝的木质部结构与水力功能之间不存在显著相关(图5E)。野核桃的比导率与导管密度、导管壁厚度跨度比呈极显著负相关(P<0.01),与木材基本密度呈显著正相关(P<0.05,图5F)。
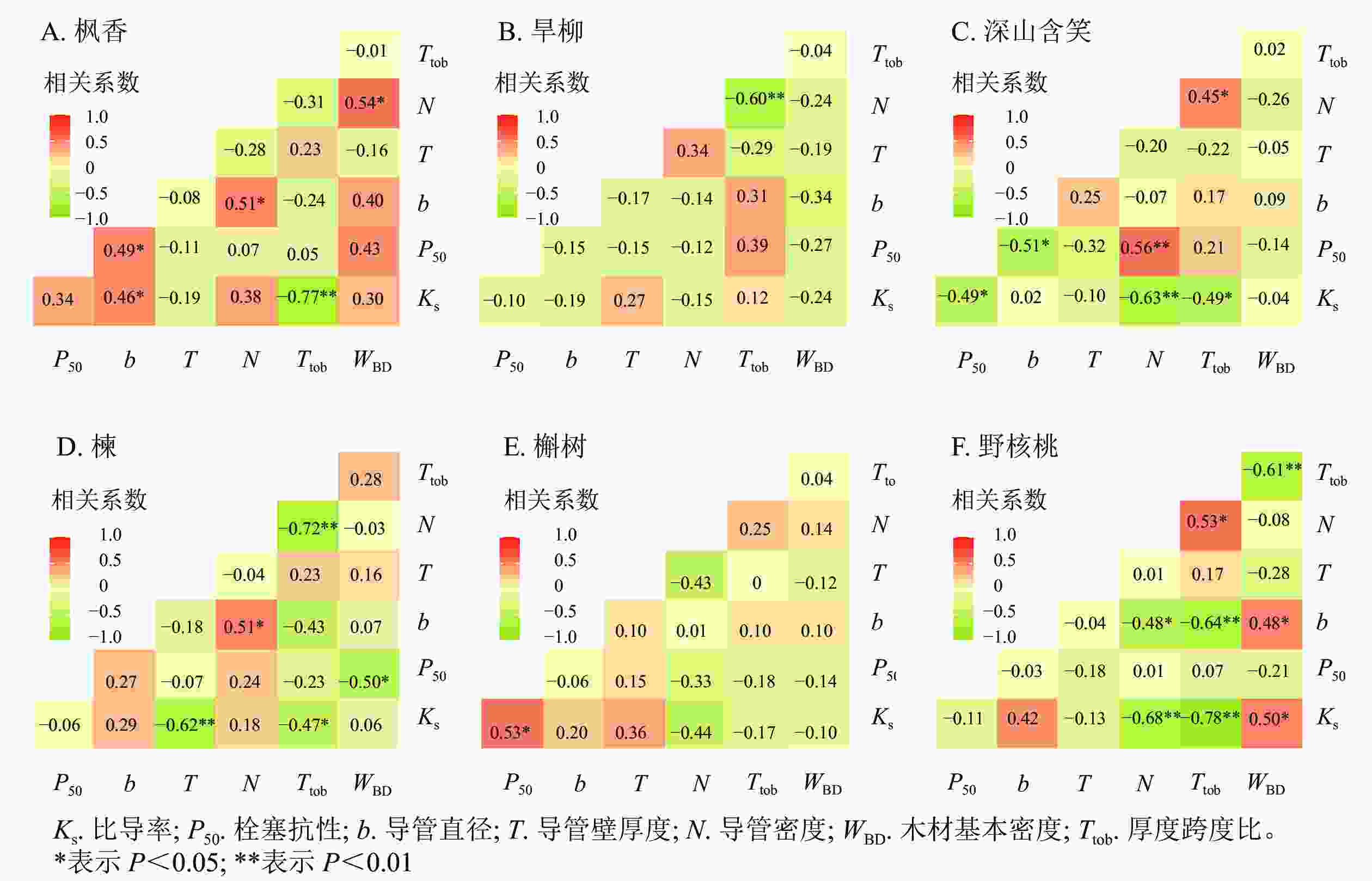
图 5 6个树种枝条木质部结构与水力功能的相关性
Figure 5. Correlation between xylem structure and hydraulic function of six tree species
-
本研究表明:散孔材树种的水分运输效率低于环孔材树种,但栓塞抗性比环孔材树种更强,这与SPERRY等[11]的研究结果相符。这2种材性植物的导管结构特征导致水分生理调节存在显著差异[24-25]。本研究的3个环孔材树种的平均木质部导管直径显著大于3个散孔材树种。大的导管直径虽然提升了水力效率,但是也会导致较小的栓塞抗性[26]。在木质部边材面积一定的情况下,单个导管的横截面积越大,边材所能容纳的导管数量也会相应减少[27]。本研究的3个散孔材树种的平均导管密度显著大于3个环孔材树种。虽然环孔材树种较大的导管直径使其获得较高的水分运输能力,但是其较大的导管直径在水分运输上的优势会被较低的导管密度抵消[28]。导管数量的增加对于水分运输能力的提升是有限的,对水分运输能力的影响远远没有导管直径增加带来的影响大[27]。本研究的3个散孔材树种的平均导管壁厚度跨度比显著大于3个环孔材树种,根据Pearson相关分析:导管壁厚度跨度比与散孔材和环孔材树种的Ks、P50相关显著。虽然木质部栓塞抗性与导管直径、导管壁厚度均存在相关关系[29],但COCHARD等[17]研究发现:导管壁厚度跨度比与栓塞抗性的相关性更紧密,导管壁厚度跨度比越大,树种栓塞抗性越强。本研究表明:不同材性树种水分利用策略不同,水力功能与解剖结构相协调;与其他结构指标相比,导管壁厚度跨度比能更好地指示不同材性树种栓塞抗性的强弱。
“安全”的木质部意味着对导管的充分保护,主要是防止空穴化和栓塞的发生,“效率”意味着一定导管组织投资下,导管水力阻力较低。在“安全”和“效率”之间存在权衡关系,即水分运输效率高的物种易发生栓塞,导致其安全性低;而水分运输效率低的物种则栓塞抗性高[30]。本研究中,除散孔材深山含笑外,其他树种比导率与栓塞抗性均呈正相关或无明显相关性,表明这些物种不存在水力效率-安全权衡。研究表明:树种中的Ks和P50之间只存在微弱的权衡或没有权衡[31]。GLEASON等[32]研究发现:栖息地、环境条件和植物生理结构的差异可能会造成一部分植物不存在水力效率与安全性之间的权衡。综上所述,木质部效率-安全权衡仍存在争议,不同生境或不同分类单元的植物可能具有物种特异性。
散孔材与环孔材树种枝输水效率、栓塞抗性间显著差异,且Ks和P50的高低均与枝木质部解剖结构特性相关。输水效率低的树种需要构建一个安全的木质部以降低栓塞风险。散孔材树种的导管直径小,导管密度和导管壁厚度跨度比大,因此,与环孔材树种相比,散孔材树种的栓塞抗性更强,在遭受水分胁迫时能更好地保证树体的存活。此外,2种材性树种中5个树种水力效率与水力安全均不存在权衡关系,这可能与植物生长所需的环境条件和植物生理结构上的差异有关。但水力功能性状与结构性状对生态功能的预测能力,以及对环境变化的响应等问题都是因地而异,仍需在不同尺度上开展大量的研究。
Relationship between xylem structure and function of diffuse-porous and ring-porous wood species in Jigongshan Nature Reserve
-
摘要:
目的 木质部解剖结构是植物水力功能和水分利用策略的基础。散孔材和环孔材的导管分布和形态存在显著差异,对2种材性树种木质部解剖结构和水力功能的关系进行比较,有助于理解不同材性树种的水分适应机制。 方法 选取河南省信阳市鸡公山自然保护区中的3个散孔材树种(枫香Liquidambar formosana、旱柳Salix matsudana、深山含笑Michelia maudiae)和3个环孔材树种(槲树Quercus dentata、楝Melia azedarach、野核桃Juglans cathayensis),研究2种材性树种的枝水力功能(比导率和栓塞抗性)和解剖结构的关系。 结果 3个环孔材树种比导率更大,3个散孔材树种栓塞抗性更强;在物种水平上,只有散孔材树种深山含笑比导率和栓塞抗性呈显著负相关(P<0.05),存在水力效率-安全权衡。3种散孔材树种相较于3种环孔材树种具有更大的导管密度、导管壁厚度跨度比和更小的导管直径、导管壁厚度。 结论 环孔材树种倾向于通过增大导管直径显著提高其输水效率,从而避免水势下降、降低潜在栓塞风险。散孔材树种木质部结构中的导管壁厚度和导管壁厚度跨度比对植物水分传输功能具有重要影响。图5表1参32 Abstract:Objective The anatomical structure of xylem is the basis of plant hydraulic function and water use strategy. There are significant differences in vessel distribution and morphology between diffuse-porous and ring-porous wood species. The purpose of this study is to compare anatomical structure and hydraulic function of xylem of the two wood species, as well as the quantitative relationship between structure and function, so as to understand the water adaptation mechanism of different wood species. Method Three diffuse-porous species (Liquidambar formosana, Salix matsudana, Michelia maudiae) and three ring-porous species (Quercus dentata, Melia azedarach, Juglans cathayensis) in Jigongshan Nature Reserve, Henan Province, were selected to study the branch hydraulic function (specific conductivity, Ks and embolism resistance, P50) and anatomical structure, and the relationship between hydraulic function and anatomical structure was quantitatively analyzed. Result Ks of the three ring-porous species was higher and P50 of the three diffuse-porous species was stronger. At the species level, Ks of M. maudiae was negatively correlated with P50 (P<0.05), and there was a tradeoff between hydraulic efficiency and safety. Compared with the three ring-porous wood species, the three diffuse-porous species had higher vessel density, vessel thickness span ratio, smaller vessel diameter and vessel wall thickness. Conclusion The ring-porous species tend to significantly improve their water delivery efficiency by increasing vessel diameter to avoid water potential decline and reduce the risk of embolism. The ratio of vessel wall thickness and vessel thickness span in the xylem structure of the diffuse-porous species has important effects on plant water transfer function. [Ch, 5 fig. 1 tab. 32 ref.] -
图 2 2种材性树种的比导率(Ks)与栓塞抗性(P50)
Figure 2 Comparison of specific sapwood conductivity (Ks) and cavitation resistance (P50) of tree species with two wood properties
图 3 2种材性树种解剖结构性状
Figure 3 Comparison of the structural characteristics of two wood-based tree species
图 4 6个树种比导率(Ks)与栓塞抗性(P50)的关系
Figure 4 Relationship between specific conductance (Ks) and embolic resistance (P50) of six tree species
图 5 6个树种枝条木质部结构与水力功能的相关性
Figure 5 Correlation between xylem structure and hydraulic function of six tree species
表 1 散孔材和环孔材样树的基本特性
Table 1. Basic characteristics of diffuse- and ring-porous sampled trees
材性 树种 树高/m 胸径/cm 树龄/a 冠幅/m 散孔材 枫香 17.45±0.35 16.26±0.12 15~20 4.10±0.22 旱柳 12.75±0.25 12.18±0.24 10~15 2.14±0.07 深山含笑 12.41±0.43 13.65±0.54 15~20 3.45±0.14 环孔材 槲树 13.32±0.25 13.34±0.27 15~20 5.86±0.08 楝 8.78±0.11 9.15±0.42 10~15 2.54±0.23 野核桃 11.33±0.17 13.42±0.15 15~20 4.02±0.21 说明:数值为平均值±标准误;样本数为20 株·种−1  下载: 导出CSV
下载: 导出CSV
-
[1] KRAMER P J. Water Relation of Plants[M]. New York: Academic Press, 1983: 342 − 489. [2] ADAMS H D, ZEPPEL M J B, ANDEREGG W R L, et al. A multi-species synthesis of physiological mechanisms in drought-induced tree mortality [J]. Nat Ecol Evol, 2017, 1: 1285 − 1291. [3] 陆世通, 陈森, 李彦, 等. 罗汉松科3种植物茎和根木质部水分运输、解剖结构与机械强度之间的关系[J]. 植物生态学报, 2021, 45(6): 659 − 669. LU Shitong, CHEN Sen, LI Yan, et al. Relationships among xylem transport, anatomical structure and mechanical strength in stems and roots of three Podocarpaceae species [J]. J Plant Ecol, 2021, 45(6): 659 − 669. [4] 叶琳峰, 李彦, 王忠媛, 等. 湿润地区3种松属植物枝和根导水系统的效率-安全关系[J]. 林业科学, 2021, 57(7): 194 − 204. YE Linfeng, LI Yan, WANG Zhongyuan, et al. Efficiency-safety relationships of hydraulic conducting system for branch and root of three Pinus species growing in humid area [J]. Sci Silv Sin, 2021, 57(7): 194 − 204. [5] TYREE M T, EWERS F W. The hydraulic architecture of trees and other woody plants [J]. New Phytol, 1991, 119(3): 345 − 360. [6] 李吉跃, 翟洪波. 木本植物水力结构与抗旱性[J]. 应用生态学报, 2000, 11(2): 301 − 305. LI Jiyue, ZHAI Hongbo. Hydraulic architecture and drought resistance of woody plants [J]. J Appl Ecol, 2000, 11(2): 301 − 305. [7] CHOAT B, JANSEN S, BRODRIBB T J, et al. Global convergence in the vulnerability of forests to drought [J]. Nature, 2012, 491: 752 − 755. [8] WILLIGEN C V, SHERWIN H W, PAMMENTER N W. Xylem hydraulic characteristics of subtropical trees from contrasting habitats grown under identical environmental conditions [J]. New Phytol, 2000, 145: 51 − 59. [9] HACKE U G, SPICER R, SCHREIBER S G, et al. An ecophysiological and developmental perspective on variation in vessel diameter [J]. Plant Cell Environ, 2017, 40(6): 831 − 845. [10] COCHARD H. Vulnerability of several conifers to air embolism [J]. Tree Physiol, 1992, 11(1): 73 − 83. [11] SPERRY J S, NICHOLS K L, SULLIVAN J E M, et al. Xylem embolism in ring-porous, diffuse-porous, and coniferous trees of northern Utah and interior Alaska [J]. Ecology, 1994, 75(6): 1736 − 1752. [12] NIU C Y, MEINZER F C, HA G Y. Divergence in strategies for coping with winter embolism among co-occurring temperate tree species: the role of positive xylem pressure, wood type and tree stature [J]. Funct Ecol, 2017, 31(8): 1550 − 1560. [13] BUSH S E, PATAKI D E, HULTINE K R, et al. Wood anatomy constrains stomatal responses to atmospheric vapor pressure deficit in irrigated, urban trees [J]. Oecologia, 2008, 156(1): 13 − 20. [14] 左力翔, 李俊辉, 李秧秧, 等. 散孔材与环孔材树种枝干、叶水力学特性的比较研究[J]. 生态学报, 2012, 32(16): 5087 − 5094. ZUO Lixiang, LI Junhui, LI Yangyang, et al. Comparison of hydraulic traits in branches and leaves of diffuse-and ring-porous species [J]. Acta Ecol Sin, 2012, 32(16): 5087 − 5094. [15] 李荣, 姜在民, 张硕新, 等. 木本植物木质部栓塞脆弱性研究新进展[J]. 植物生态学报, 2015, 39(8): 838 − 848. LI Rong, JIANG Zaimin, ZHANG Shuoxin, et al. A review of new research progress on the vulnerability of xylem embolism of woody plants [J]. J Plant Ecol, 2015, 39(8): 838 − 848. [16] 金鹰, 王传宽. 九种不同材性的温带树种叶水力性状及其权衡关系[J]. 植物生态学报, 2016, 40(7): 702 − 710. JIN Ying, WANG Chuankuan. Leaf hydraulic traits and their trade-offs for nine Chinese temperate tree species with different wood properties [J]. J Plant Ecol, 2016, 40(7): 702 − 710. [17] COCHARD H, BARIGAH S T, KLEINHENTZ M, et al. Is xylem cavitation resistance a relevant criterion for screening drought resistance among Prunus species [J]. J Plant Physiol, 2008, 165(9): 976 − 982. [18] JACOBSEN A L, BRANDON P R. Going with the flow: structural determinants of vascular tissue transport efficiency and safety [J]. Plant Cell Environ, 2018, 41(12): 2715 − 2717. [19] 李荣, 党维, 蔡靖, 等. 6个耐旱树种木质部结构与栓塞脆弱性的关系[J]. 植物生态学报, 2016, 40(3): 255 − 263. LI Rong, DANG Wei, CAI Jing, et al. Relationships between xylem structure and embolism vulnerability in six species of drought tolerance trees [J]. Chin J Plant Ecol, 2016, 40(3): 255 − 263. [20] ZIV B, SAARONI H, PARGAMENT R, et al. Trends in rainfall regime over Israel, 1975 − 2010, and their relationship to large-scale variability [J]. Reg Environ Change, 2014, 14(5): 1751 − 1764. [21] SHI Benlin, ZHU Xinyu, HU Yunchuan, et al. Drought characteristics of Henan Province in 1961 − 2013 based on standardized precipitation evapotranspiration index [J]. J Geogr Sci, 2017, 27(3): 311 − 325. [22] 周洪华, 李卫红. 胡杨木质部水分传导对盐胁迫的响应与适应[J]. 植物生态学报, 2015, 39(1): 81 − 91. ZHOU Honghua, LI Weihong. Responses and adaptation of xylem hydraulic conductivity to salt stress in Populus euphratica [J]. Chin J Plant Ecol, 2015, 39(1): 81 − 91. [23] HACKE U G, SPERRY J S, POCKMAN W T, et al. Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure [J]. Oecologia, 2001, 126(4): 457 − 461. [24] MATHENY A M, BOHRER G, VOGEL C S, et al. Species-specific transpiration responses to intermediate disturbance in a northern hardwood forest [J]. J Geophys Res Biogeosci, 2014, 119: 2292 − 2311. [25] PETERS E B, MCFADDEN J P, MONTGOMERY R A. Biological and environmental controls on tree transpiration in a suburban landscape[J/OL]. J Geophys Res Biogeosci, 2015, 115: G04006[2021-08-28]. doi: 10.1029/2009JG001266. [26] COCHARD H, BREDA N, GRANIER A, et al. Vulnerability to air embolism of three European oak species (Quercus petraea (Matt) Leibl, Q. pubescens Willd, Q. robur L. ) [J]. Ann For Sci, 1992, 49(3): 225 − 233. [27] 丁俊杰, 张鑫, 楚光明, 等. 3种荒漠植物导管特征及其可塑性研究[J]. 干旱区资源与环境, 2016, 30(9): 171 − 177. DING Junjie, ZHANG Xin, CHU Guangming, et al. Study on vessel characteristics and plasticity of three desert plants [J]. J Arid Land Resour Environ, 2016, 30(9): 171 − 177. [28] NARDINI A, PEDA G, ROCCA N L. Trade-offs between leaf hydraulic capacity and drought vulnerability: morphoanatomical bases, carbon costs and ecological conesquences [J]. New Phytol, 2012, 196(3): 788 − 798. [29] HARGRAVE K R, KOLB K J, EWERS F W, et al. Conduit diameter and drought-induced embolism in Salvia mellifera Greene (Labiatae) [J]. New Phytol, 1994, 126(4): 695 − 705. [30] HACKE U G, SPERRY J S, WHEELER J K, et al. Scaling of angiosperm xylem structure with safety and efficiency [J]. Tree Physiol, 2006, 26(6): 689 − 701. [31] SCHULDT B, KNUTZEN F, DELZON S, et al. How adaptable is the hydraulic system of European beech in the face of climate change-related precipitation reduction?[J] New Phytol, 2016, 210: 443 − 458. [32] GLEASON S M, WESTOBY M, JANSEN S, et al. Weak tradeoff between xylem safety and xylem-specific hydraulic efficiency across the world’s woody plant species [J]. New Phytol, 2016, 209(1): 123 − 136. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210628
 点击查看大图
点击查看大图
计量
- 文章访问数: 3656
- HTML全文浏览量: 668
- PDF下载量: 86
- 被引次数: 0
