




-
随着城市化加速发展,不透水地面逐渐增多,城市热岛效应(urban heat island effect,UHI)日益凸显。学者们多从热环境时空变化[1−2]、影响机制及驱动力[3]、城市热岛效应缓解方法[4]等方面展开研究,但较少关注景观网络在缓解热岛效应中的作用。这些研究表明,地表温度受斑块间的热交换影响显著,热源与热汇的连通性是影响热流动的重要因素。然而,地表温度难以反映整体格局和连通性,需要合适的方法来准确描述热环境的空间格局。对城市热岛的研究侧重于整体区域尺度或是斑块水平上的统计分析,忽视了景观网络对缓解城市热环境的作用。陈利顶等[5]利用“源-汇”理论,将热环境与生态过程结合,为解决城市热环境问题提供新思路。
城市绿地对缓解城市热岛效应具有重要作用[6],当绿地覆盖面积在40%以下时,绿地系统的空间格局将对环境的增势以及降温产生主要影响[7],在有限的城市空间中增加大面积的绿地已经难以实现,因此通过优化绿地空间格局来缓解城市热岛效应尤为重要。基于景观生态学“源-汇”理论,识别城市热岛像元与绿地像元,构建多层级生态网络,将是缓解城市热环境的重要手段。生态网络构建方法主要包括形态空间格局分析方法(morphological spatial pattern analysis,MSPA)、最小累积阻力模型以及重力模型等 [8−9]。MSPA方法强调景观内部结构性的连接,可以准确地将前景要素划分为核心、孤岛、孔隙、边缘、环道、桥接和支线等7类,为后期廊道以及生态节点的识别提供理论依据[10−12]。近年来MSPA方法也逐渐应用到城市热岛的研究中,以达到缓解城市热岛效应的目的。景观连通性指数包括整体连通性指数(integral index of connectivity,IIC)、可能连通性指数(probability of connectivity,PC)等,反映了景观对生态过程中能量流动的促进或阻碍作用大小,良好的景观连通性有助于构建稳定的生态环境[13]。最小累积阻力模型是指物种从源地向目标迁移扩散过程中,穿越不同景观表面所需耗费的最小代价的模型[14],最小累积阻力模型与重力模型相结合能更好地识别生态廊道间的相互作用强度,以筛选具有重要作用的关键廊道。目前,大多数研究利用MSPA、景观连通性指数、最小累积阻力模型等方法进行绿地生态网络的构建,但利用该方法体系构建缓解城市热环境的多层级生态网络的研究相对较少。
本研究以成都市中心城区为研究对象,基于“源-汇”理论,利用MSPA与景观连通性指数,筛选研究区“源”“汇”景观,利用最小累积阻力模型、重力模型以及水文分析模块构建“源-源”“汇-汇”“源-汇”景观廊道以及生态节点,最终形成具备“补偿-运输-作用”功能的多层级景观网络格局,确定需要重点保护的生态用地、重要廊道以及关键节点,提出优化策略,为成都市生态网络空间的构建提供有效支撑。
-
成都市位于川西平原,30°22′~30°96′N,103°68′~104°49′E,地势较为平坦,由于地形影响,夏季炎热,冬季寒冷。本研究的中心城区(图1)包括郫都区、新都区、青白江区、温江区、金牛区、成华区、龙泉驿区、青羊区、武侯区、锦江区、双流区等11个行政区,总面积为3 732.06 km2。中心城区处于全国两大静风区之一,建筑及人口密度高,地表通风能力弱[15],不利于城市内部热量扩散,城市热环境矛盾突出,因此具有研究城市热岛效应的典型特征。
图 1 研究区范围示意图
Figure 1. The study area
-
所用数据包括2020年空间分辨率为30 m的Landsat 8 OLI卫星影像数据(
http://earthexplorer.usgs.gov ),空间分辨率为30 m的DEM高程数据(https://www.gscloud.cn/ ),以及《成都市国土空间总体规划(2020—2035年)》(草案)等相关规划图件。利用ENVI软件对获取的遥感影像预处理后利用覃志豪等[16]的单窗算法反演地表温度;利用监督分类的方法,将研究区2020年的土地利用类型细分为草地、林地、耕地、水域以及城乡建设用地等5种类型,通过实地调研踏勘与高分辨率遥感影像的目视判别,对分类结果进行校正,最终解译精度达89%以上。 -
通过计算城市区域与周边地区的平均温度之差来确定相对热岛强度,相对热岛强度越高,相应区域内热岛效应越明显[17]。在ArcGIS中通过计算不同用地类型的相对热岛强度来判别“源-汇”景观。按照LAL等[18]和贾玉雪等[19]的研究将计算结果中相对热岛强度(H)≥0的斑块定义为对城市热环境有促进作用的“源”景观,H<0的景观定义为“汇”景观。
-
将“源”景观作为前景,赋值为2,“汇”景观作为背景,赋值为1,并将其转化为30 m×30 m栅格数据;运用Guidos Toolbox软件对其进行MSPA分析,设置8邻域的连通规则,边缘宽度为1,获得7种景观类型:核心区、边缘、孤岛、桥接区、环道、支线和孔隙。按相同步骤将“汇”景观作为前景,“源”景观作为背景,得到“汇”景观的空间形态格局。
-
景观连通性指数可以衡量不同空间单元之间景观要素的连通性。量化景观要素在生态源地之间进行扩散或者迁移的难易程度,也是衡量生态过程之间联系程度的重要指标[20]。利用Conefor 2.6软件,通过计算IIC、PC以及斑块重要性(dI’)来衡量不同核心斑块的重要程度[21]。考虑研究区内斑块的面积和连通性,通过反复测试计算,设定斑块连接性阈值为2 000,连通概率为0.5。最后,基于景观连通性指数dI’值大小综合评估核心区斑块的景观重要程度。
-
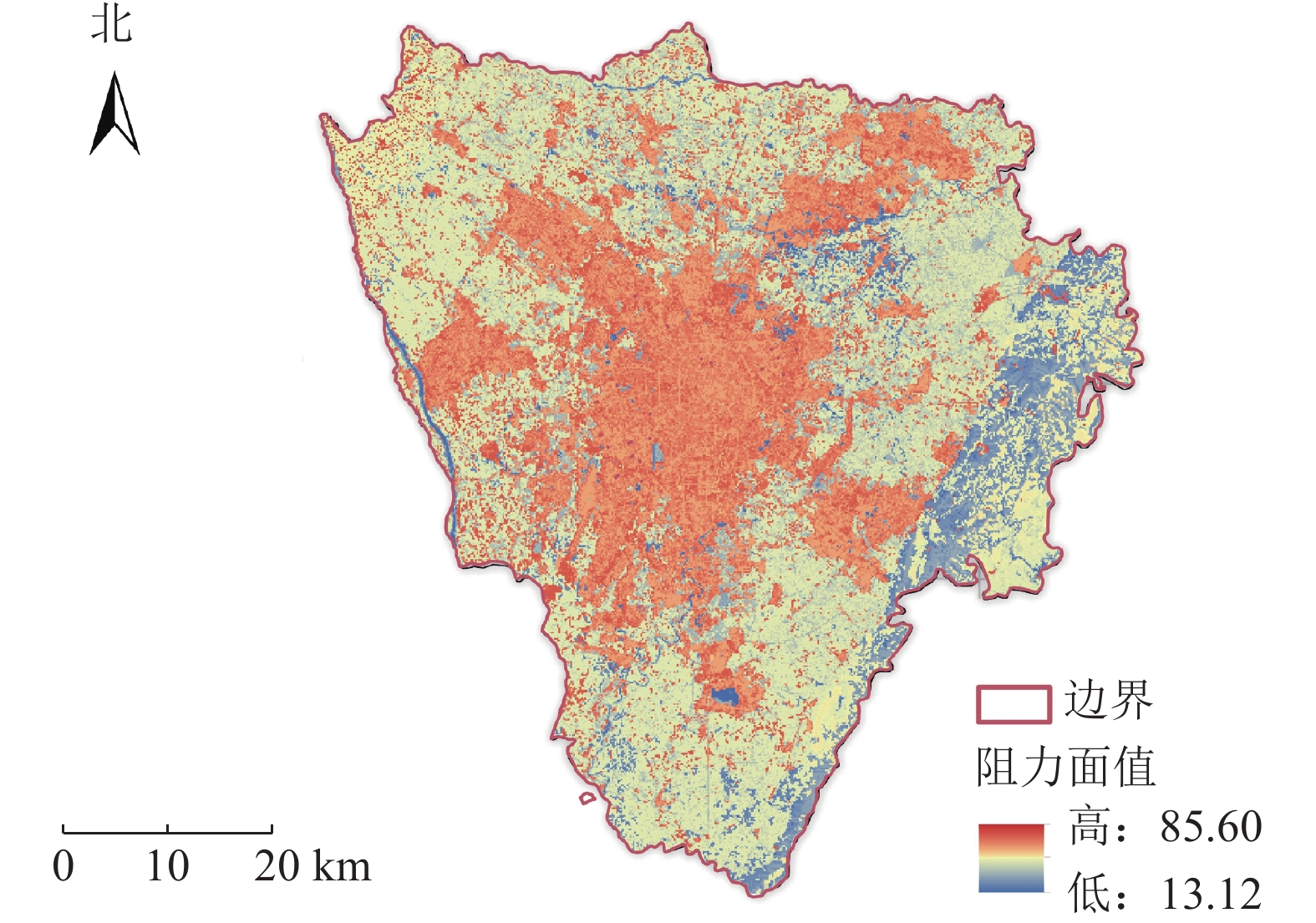
根据研究区现状以及数据的可获取性,最终选取用地类型、高程、坡度以及归一化植被指数(NDVI)来构建综合阻力面。其中高程决定了城市内不同区域的温度分布,坡度影响空气流动和热量累积,不同用地类型对城市热环境产生不同影响,而NDVI则反映了植被覆盖情况,对城市温度、热岛效应和空气质量有重要影响。采用专家打分法确定因子阻力值,并采用层次分析法(AHP)计算其权重值(表1),通过叠加分析最终生成综合阻力面(图2)。可以看出,研究区内阻力值的呈现由中心向四周扩散,逐渐递减,尤其是东南方向的递减最为明显。
表 1 赋予不同影响因子的阻力值
Table 1. Resistance values assigned to different impact factors
影响因子 类型分级 赋予阻力值 所占权重 影响因子 类型分级 赋予阻力值 所占权重 用地分类 林地 10 0.520 坡度/( º ) 0~10 10 0.078 水地 20 10~20 30 草地 30 20~30 50 耕地 50 30~40 70 未利用土地 70 40~50 90 建设用地 100 >50 100 高程/m <200 10 0.078 归一化植被
指数(NDVI)−1.00~−0.20 10 0.201 200~400 30 −0.20~0.30 30 400~600 50 0.30~0.50 50 600~800 70 0.50~0.70 70 800~1 000 90 0.70~1.00 90 >1 000 100 
图 2 阻力面示意图
Figure 2. Resistance surface
-
在ArcGIS中,利用Cost-distance工具构建研究区的累积耗费距离表面。利用Cost-path构建多对多的潜在生态廊道,以连接不同的“源-汇”景观。最后,利用重力模型[22]计算生态廊道间的相互作用强度,通过筛选合适的强度阈值,确保所有的“源-汇”景观均被连通,从而提取出“源-源”“汇-汇”“源-汇”生态廊道。
-
识别生态廊道中的关键点和障碍点能够为物种的迁徙及物种保护区的划分和规划提供科学依据[23−24]。在ArcGIS中,运用水文分析模块,对累积耗费距离表面进行水流方向、汇流累积量等一系列分析计算。通过对比不同阈值设定下最小阻力路径的完整性与连通性,最后确定阈值为500构建研究区内的低阻力廊道。运用ArcGIS中的Intersect工具将低阻力值廊道与“源-源”“汇-汇”廊道进行相交分析从而获得不同生态节点,包括生态障碍点与生态关键点,其中生态关键点是生态廊道中能量流动密度较大的点,需要对关键点进行有效利用与保护;对障碍点则需进行生态修复来提升廊道整体的连接度,以保障冷热能的有效传递。
-
将“源-源”“汇-汇”“源-汇”廊道共同相交[25],得到一级补偿廊道;将“汇-汇”“源-汇”廊道进行相交,得到二级输送引导廊道;将“源-源”“源-汇”廊道进行相交,得到三级作用廊道,完成廊道的“补偿-运输-作用”的完整体系,构建完整的多层级“源-汇”生态景观网络。
-
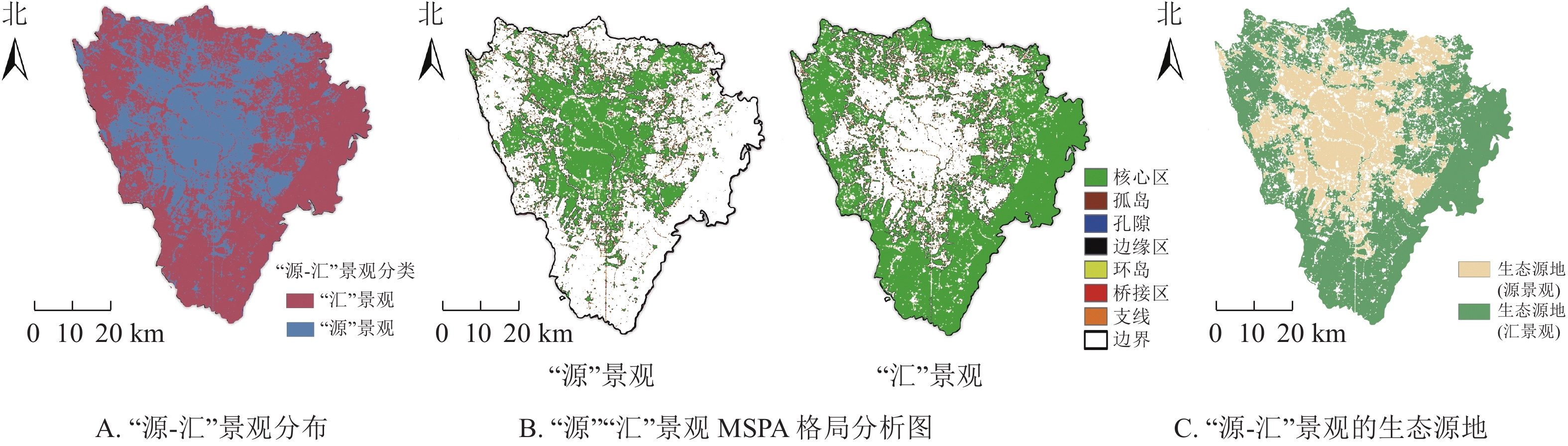
从图3A可以看出:“源”景观在研究区中部呈现聚集特征,“汇”景观大多分散分布在研究区的西北部以及东南部。其中,“汇”景观斑块总计
98342 个,占研究区域总面积的62.1%,以大面积的带状水域和块状绿地为主;“源”景观斑块总计212 231个,占研究区域总面积的37.9%,由大面积建设用地组成。
图 3 生态源地的识别示意图
Figure 3. Ecological source of the “source - sink” landscape
-
从“源-汇”景观的MSPA格局分析(图3B)可以看出:“源”核心景观密集地分布在研究区中部,多为城市建设用地;研究区西北以及东南两侧的“源”景观核心斑块较为破碎,景观连通性较差。研究区中部的“汇”景观由于城市绿地破碎化严重导致空间连通性较差。对比不同景观要素类型面积比(表2)发现:“源”景观核心区面积为1 169.33 km2,占“源”景观前景要素总面积的31.83%;“汇”景观核心区面积为2 053.78 km2,占“汇”景观前景要素总面积的55.91%,对城市热岛效应起重要作用。最终,分别筛选面积在0.01 km2以上的源、汇核心斑块作为重要核心斑块,以进行景观连通性分析。
表 2 “源”“汇”景观要素不同类型面积占比
Table 2. Area proportion of different types of “source” “sink” landscape elements
景观类型 “汇”景观面
积占比/%“源”景观面
积占比/%核心区 55.91 31.83 孤岛 0.07 0.14 孔隙 2.46 1.61 边缘区 3.32 3.77 环岛 0.06 0.04 桥接区 0.06 0.08 支线 0.21 0.26 -
将dI’值大于0.1的斑块作为研究的生态源地,分别筛选出24 个“源”“汇”生态源地(图3C),其中“源”景观生态源地占研究区总面积的28.81%,“汇”景观生态源地占研究区总面积的53.60%。“汇”生态源地主要分布在青白江生态带、龙泉山国家森林片区、三圣乡片区、青龙湖湿地公园片区、兴隆湖湿地公园片区以及江安河流域段,而“源”景观生态源地主要分布在金牛区、成华区、锦江区、青羊区以及武侯区(简称“五城区”)。
-
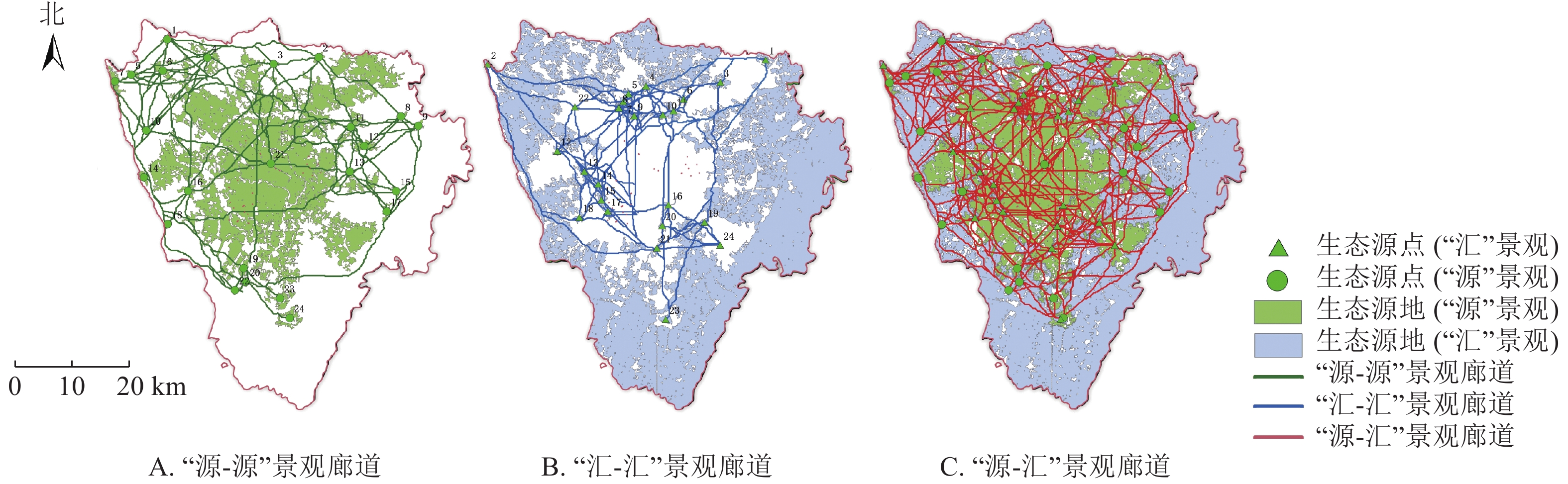
基于最小成本路径构建“源-源”廊道276条,“汇-汇”廊道266条,“源-汇”廊道690条。利用重力模型最终筛选出“源-源”廊道102条,总长度为2 081.6 km,“汇-汇”廊道141条,总长度为1 907.8 km,“源-汇”廊道325条,总长度为7 698.0 km (图4)。其中“源”景观23、24号生态源点与“汇”景观23号生态源点仅由单条景观廊道连通,表明它们在整个热环境中相对独立,呈孤岛状分布,受到其他景观斑块的影响较小,导致在整个热传导的过程中不能发挥良好的作用。

图 4 “源-汇”景观廊道分布示意图
Figure 4. “Source-sink” landscape corridor distribution
-
统计不同行政区内各廊道的占比情况(表3)发现:新都区、郫都区与双流区内各类“源-汇”重要廊道最多,主要以毗河、岷江等河流,部分廊道呈簇团状分布,说明在研究区冷热交换过程中起到了重要作用;由于五城区分布有大片的“源”景观生态源地,且建筑密度相对较高,区域内各层级“源-汇”重要廊道分布较少,导致其在冷热交换过程中发挥的作用较小。尤其是成华区建筑密度为14.49%,各层级廊道占比都相对较低,区域内大量热空气堆积不易扩散,热岛效应明显。
表 3 研究区各行政区景观廊道分布
Table 3. Distribution of landscape corridors in each administrative district
行政区 建筑密度/% “源-源”廊道长度/km 所占比例/% “汇-汇”廊道长度/km 所占比例/% “源-汇”廊道长度/km 所占比例/% 新都区 20.50 825.32 26.6 642.65 17.2 23 563.22 21.87 郫都区 20.43 518.02 16.7 810.47 21.8 17 422.12 16.17 双流区 12.72 509.82 16.4 689.71 18.6 23 944.23 22.22 温江区 19.12 357.46 11.5 233.84 6.2 9 438.32 8.76 龙泉驿区 11.16 345.57 11.1 171.33 4.6 9 768.86 9.06 金牛区 18.99 229.24 7.3 379.07 6.2 5 423.08 5.03 青白江区 15.01 201.95 6.5 31.16 0.8 3 699.39 3.43 成华区 14.49 90.75 2.9 18.13 0.5 2 827.71 2.62 锦江区 18.01 11.05 0.3 177.75 4.7 3 295.24 3.06 青羊区 28.09 7.04 0.2 303.22 8.2 3 500.85 3.25 武侯区 22.20 5.26 0.2 409.52 11.0 4 882.50 4.53 -
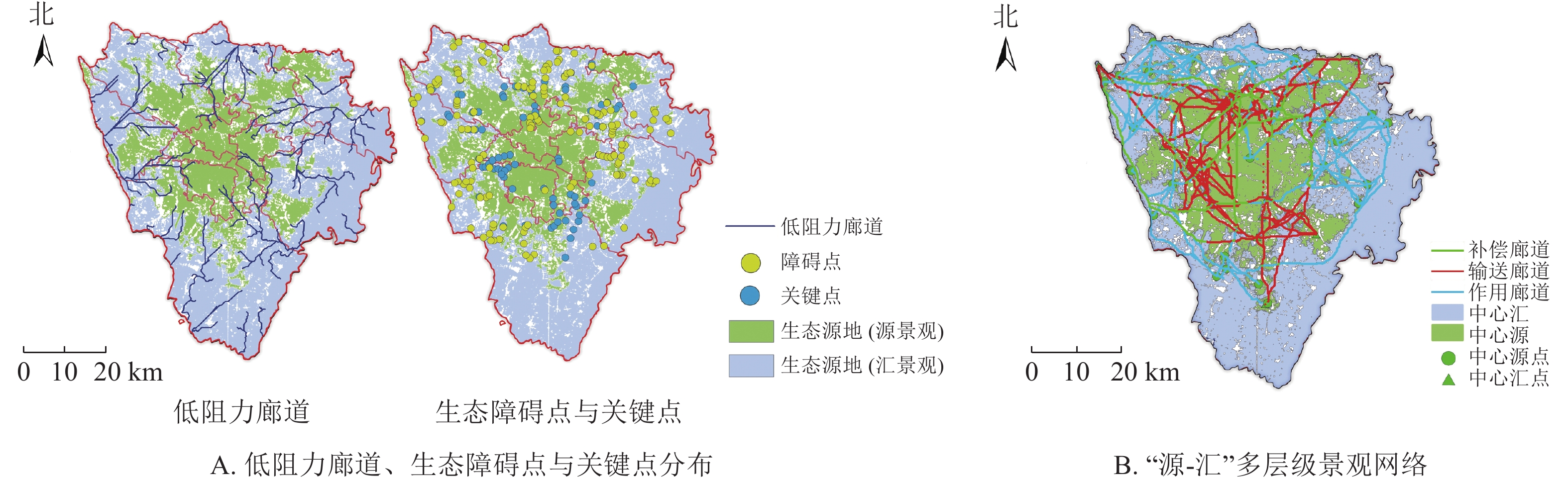
利用水文分析模块获取低阻力廊道95条,将低阻力廊道与“源-汇”廊道相交分析得到生态障碍点148个,生态关键点103个(图5A),其中生态关键点在青羊区、武侯区以及锦江区与双流区交汇处出现堆积现象,导致该区域出现功能廊道不能充分利用的问题。生态关键点整体分布与“汇”景观生态源地分布情况大致相同,这意味着加强“汇”景观源地的生态建设,降低周边阻力值,将会有效提升网络连通性。而生态障碍点大多聚集在建筑密度相对较高的区域,生态障碍点堆积处出现大量的热能无法有效被传输,加强生态障碍点的生态修复对城市热量的传导具有重要作用。

图 5 低阻力廊道、生态障碍点与关键点分布(A)及“源-汇”多层级景观网络示意图(B)
Figure 5. Distribution of low-resistance corridors, ecological barrier points and key points (A), and “source-sink” multi-level landscape network (B)
-
多层级“源-汇”景观网络中(图5B),一级补偿廊道36条,主要由岷江、毗河和其他河流廊道构成,分布在研究区西部以及北部,在城市中发挥着冷热空气交换的重要作用,是补充能量的主要途径;二级输送引导廊道125条,主要分布在西部、南部以及北部的三环路附近,起到将冷空气运输和分配的作用,是实现能量传递的次要路线;三级作用廊道86条,主要分布在研究区西北以及东北部,主要承担实现热空气的运输和分配的任务,是“源-汇”景观能量交换过程的末端环节。3种廊道共同作用,实现了廊道的“补偿-输送-作用”的功能,以达到缓解城市热岛效应的作用。
-
优化生态网络中的“源-汇”景观源地对改善城市热环境具有重要作用。在五城区中大量建筑密度高、人口高度密集的“源”景观生态源地,应加强垂直绿化、屋顶绿化等来增加植被覆盖率;在源地周围增加社区口袋公园、绿化带等构建缓冲区,以改善城市生态环境。对于龙泉驿区、双流区等植被覆盖率高、生态质量好的“汇”景观生态源地,可建设生态公园和自然保护区、引进生态景观设计等,以增强其改善气候环境的生态效能。
-
对区域内生态障碍点来说,可以推广绿色建筑以有效地吸收太阳辐射,对新建城区的建筑布局进行合理规划,降低建筑密度、增加绿色基础服务设施以降低城市表面的温度,有效改善城市热岛效应。对生态关键点可退耕还林、扩大区域植被绿化面积、建立生态缓冲区等来降低生态关键点周边的阻力值,以确保生态关键点与生态廊道的连通性,保障热量之间的相互流通。
-
一级补偿廊道多依托水系以及绿道进行构建,是调节气候的关键要素,可对其进行生态规划保护,包括河道整治、整合岸线资源以及拓宽绿道宽度等。二级输送引导廊道相当于城市通风廊道,可对廊道布局、地形特征和内部设施等方面合理规划,以保证冷空气的输送,尤其需注重绿地植被结构的优化设计,确保其通透性。三级作用廊道主要起热交换的作用,可以拓宽廊道横截面、加强沿线绿化建设等提高其作用效率。
-
通过生态缓冲区的建设提高生态关键点与障碍点的生态环境质量,增强与生态廊道的有效连接与过渡,强化“源”“汇”景观生态源地之间的相互渗透,增加绿色基础设施建设,减弱高密度建成区对自然生态环境的干扰,推进城市生态环境多层次、立体化、网络化的建设思路,整体提升生态网络缓解城市热岛效应的能力。
-
本研究共筛选“源” “汇”景观源地24个,“源-源”廊道102条,“汇-汇”廊道141条,“源-汇”廊道325条,生态关键点103个,生态障碍点148个。多层级景观网络中,一级补偿廊道36条,二级输送廊道125条,三级作用廊道86条,分布在研究区北部、南部与西北部。
与其他研究相比[15−16],本研究利用MSPA方法提取研究区内与城市热环境相关的“源-汇”景观核心斑块,计算景观连通性筛选“源-汇”景观生态源地,使生态源地识别过程更科学,减少生态源地识别的主观性;运用最小累积阻力模型与重力模型,最终构建多层级生态网络优化格局,该研究方法框架将为缓解城市热环境提出新的研究思路。综合运用水文分析模块构建的低阻力廊道与不同类型的“源”“汇”景观生态廊道相交,获取缓解城市热环境的生态关键点与生态障碍点,同时,将不同类型的“源”“汇”生态廊道进行相交,构建多层级的“源-汇”生态网络,分析城市建设开发状况与生态网络的空间格局关系,可更直观地揭示出生态网络脆弱区域存在的生态问题。
本研究仅对2020年的城市热环境数据展开分析,城市景观格局不断地发生变化,根据不同时期城市景观格局与城市热岛效应之间的动态变化关系,筛选具有高稳定性、高连通性的源地,综合构建缓解热环境的优化生态网络空间格局,将是后期研究的重点方向。生态网络建设是一个复杂的过程,涉及诸多因素,需要从不同尺度进行多层次分析和构建。增强城市与周边地区生态斑块之间的联系,保护核心生态斑块的完整性,保护区域的生物多样性并促进城市的可持续发展是其最终目的,因此从多尺度协同角度出发来构建综合生态网络,也是后期研究的重要方向。
Research on optimization of urban thermal environment ecological network based on MSPA and minimum cumulative resistance model
-
摘要:
目的 建立合理的城市热环境生态网络,以缓解城市热岛效应。 方法 以成都市中心城区为研究对象,基于“源-汇”理论,利用形态学空间格局分析方法(MSPA)与景观连通性指数,分别筛选研究区内景观稳定性好、连通性高的“源”“汇”景观作为生态源地;运用最小累积阻力模型、重力模型以及水文分析模块构建和筛选研究区内的重要生态廊道,从而建立缓解城市热环境问题的多层级“源-汇”景观网络。 结果 分别筛选出24个具有重要连通性的“源”“汇”核心斑块作为重要生态源地。构建“源-源”廊道102条,“汇-汇”廊道141条,“源-汇”廊道325条,生态节点103个,障碍点148个。通过综合相交分析,一级补偿廊道主要由岷江、毗河和其他河道构成,二级输送廊道主要分布在“汇”景观较为密集的区域,三级作用廊道主要分布在高强度建设区域。 结论 基于“源-汇”理论构建的多层级生态网络优化格局是有效缓解城市热岛效应的重要举措。图4表3参25 -
关键词:
- 形态空间格局分析方法(MSPA) /
- 最小累积阻力模型 /
- “源-汇”理论 /
- 城市热环境 /
- 景观生态网络
Abstract:Objective The acceleration of urbanization leads to the change of urban underlying surface, serious landscape fragmentation, and continuous intensification of urban heat island effect, especially in the urban center where buildings and populations are highly clustered. The establishment of reasonable ecological network has become one of the important ways to improve the urban ecological environment. Method Based on the “source-sink” theory, this study used morphological spatial pattern analysis (MSPA) and landscape connectivity index to select the “source” and “sink” landscapes with good landscape stability and high connectivity as ecological sources in the study area. The minimum cumulative resistance model, gravity model and hydrological analysis module were used to construct and screen important ecological corridors in the study area, so as to establish a multi-level “source-sink” landscape network to alleviate urban thermal environment problems. Result The results showed that 24 core patches with important connectivity were selected as important ecological sources. There were 102 “source-source” corridors, 141 “source-sink” corridors, 325 “source-sink” corridors, 103 ecological nodes and 148 obstacle points. Through comprehensive superposition, the first-level compensation corridor was mainly composed of Minjiang River, Pihe River and other river channels, the second-level transport corridor was mainly distributed in the area with dense “sink” landscape, and the third-level action corridor was mainly distributed in the high-intensity construction area. Conclusion The optimization pattern of a multi-level ecological network based on the “source-sink” theory is an important measure for effectively mitigating the urban heat island effect. [Ch, 4 fig. 3 tab. 25 ref.] -
图 5 低阻力廊道、生态障碍点与关键点分布(A)及“源-汇”多层级景观网络示意图(B)
Figure 5 Distribution of low-resistance corridors, ecological barrier points and key points (A), and “source-sink” multi-level landscape network (B)
表 1 赋予不同影响因子的阻力值
Table 1. Resistance values assigned to different impact factors
影响因子 类型分级 赋予阻力值 所占权重 影响因子 类型分级 赋予阻力值 所占权重 用地分类 林地 10 0.520 坡度/( º ) 0~10 10 0.078 水地 20 10~20 30 草地 30 20~30 50 耕地 50 30~40 70 未利用土地 70 40~50 90 建设用地 100 >50 100 高程/m <200 10 0.078 归一化植被
指数(NDVI)−1.00~−0.20 10 0.201 200~400 30 −0.20~0.30 30 400~600 50 0.30~0.50 50 600~800 70 0.50~0.70 70 800~1 000 90 0.70~1.00 90 >1 000 100  下载: 导出CSV
下载: 导出CSV
表 2 “源”“汇”景观要素不同类型面积占比
Table 2. Area proportion of different types of “source” “sink” landscape elements
景观类型 “汇”景观面
积占比/%“源”景观面
积占比/%核心区 55.91 31.83 孤岛 0.07 0.14 孔隙 2.46 1.61 边缘区 3.32 3.77 环岛 0.06 0.04 桥接区 0.06 0.08 支线 0.21 0.26
下载: 导出CSV
表 3 研究区各行政区景观廊道分布
Table 3. Distribution of landscape corridors in each administrative district
行政区 建筑密度/% “源-源”廊道长度/km 所占比例/% “汇-汇”廊道长度/km 所占比例/% “源-汇”廊道长度/km 所占比例/% 新都区 20.50 825.32 26.6 642.65 17.2 23 563.22 21.87 郫都区 20.43 518.02 16.7 810.47 21.8 17 422.12 16.17 双流区 12.72 509.82 16.4 689.71 18.6 23 944.23 22.22 温江区 19.12 357.46 11.5 233.84 6.2 9 438.32 8.76 龙泉驿区 11.16 345.57 11.1 171.33 4.6 9 768.86 9.06 金牛区 18.99 229.24 7.3 379.07 6.2 5 423.08 5.03 青白江区 15.01 201.95 6.5 31.16 0.8 3 699.39 3.43 成华区 14.49 90.75 2.9 18.13 0.5 2 827.71 2.62 锦江区 18.01 11.05 0.3 177.75 4.7 3 295.24 3.06 青羊区 28.09 7.04 0.2 303.22 8.2 3 500.85 3.25 武侯区 22.20 5.26 0.2 409.52 11.0 4 882.50 4.53
下载: 导出CSV
-
[1] 宫阿都, 李京, 王晓娣, 等. 北京城市热岛环境时空变化规律研究[J]. 地理与地理信息科学, 2005, 21(6): 15 − 18. GONG Adu, LI Jing, WANG Xiaodi, et al. Study on temporal and spatial distribution characteristics of the urban heat island in Beijing [J]. Geography and Geo-Information Science, 2005, 21(6): 15 − 18. [2] 刘孟竹, 李雅丽, 张红娟, 等. 1996—2017年张家口市区景观格局与地表热环境的时空变化[J]. 水土保持通报, 2021, 41(6): 303 − 309. LIU Mengzhu, LI Yali, ZHANG Hongjuan, et al. Spatiotemporal variations of landscape pattern and urban thermal environment in Zhangjiakou City during 1996−2017 [J]. Bulletin of Soil and Water Conservation, 2021, 41(6): 303 − 309. [3] 郭宇. 上海市热环境时空变化特征及其驱动机制研究[D]. 上海: 上海应用技术大学, 2022. GUO Yu. Temporal and Spatial Variation Characteristics of Urban Thermal Environment and Driving Mechanism in Shanghai City [D]. Shanghai: Shanghai Institute of Technology, 2022. [4] 杨务发, 余坤勇, 赵各进, 等. 基于热岛效应的福州市绿色廊道优化[J]. 浙江农林大学学报, 2022, 39(4): 876 − 883. YANG Wufa, YU Kunyong, ZHAO Gejin, et al. Optimization of greenways in Fuzhou based on heat island effect[J]. Journal of Zhejiang A&F University, 2022, 39(4): 876 − 883. [5] 陈利顶, 傅伯杰, 赵文武. “源”“汇”景观理论及其生态学意义[J]. 生态学报, 2006, 26(5): 1444 − 1449. CHEN Liding, FU Bojie, ZHAO Wenjie. Source-sink landscape theory and its ecological significance [J]. Acta Ecologica Sinica, 2006, 26(5): 1444 − 1449. [6] 周媛. 多元目标导向下的成都中心城区绿地生态网络构建[J]. 浙江农林大学学报, 2019, 36(2): 359 − 365. ZHOU Yuan. Developing urban greenspace ecological network in Chengdu City center based on multiple objectives [J]. Journal of Zhejiang A&F University, 2019, 36(2): 359 − 365. [7] 赵晨晓, 刘春卉, 魏家星. 缓解城市热岛效应的南京市绿色基础设施网络构建方法[J]. 浙江农林大学学报, 2021, 38(6): 1127 − 1135. ZHAO Chenxiao, LIU Chunhui, WEI Jiaxing. Green infrastructure network construction method for mitigating urban heat island effect in Nanjing [J]. Journal of Zhejiang A&F University, 2021, 38(6): 1127 − 1135. [8] 胡凤宁, 周亮. 城市绿色基础设施降温作用及其影响因素研究进展[J]. 生态学报, 2023, 43(11): 4445 − 4460. HU Fengning, ZHOU Liang. Cooling effect of urban green infrastructure and its impacting factors: a review [J]. Acta Ecologica Sinica, 2023, 43(11): 4445 − 4460. [9] 周媛, 陈明坤, 黎贝, 等. 基于空间连通性动态变化的城市绿地生态网络优化[J]. 中国城市林业, 2023, 21(4): 23 − 32. ZHOU Yuan, CHEN Mingkun, LI Bei, et al. Optimization of urban green space ecological network based on dynamic change of spatial connectivity [J]. Journal of Chinese Urban Forestry, 2023, 21(4): 23 − 32. [10] 周媛, 唐密, 陈娟, 等. 基于形态学空间格局分析与图谱理论的成都市绿地生态网络优化[J]. 生态学杂志, 2023, 42(6): 1527 − 1536. ZHOU Yuan, TANG Mi, CHEN Juan, et al. Optimization of urban green space ecological network in Chengdu based on morphological spatial pattern analysis and graph theory [J]. Chinese Journal of Ecology, 2023, 42(6): 1527 − 1536. [11] 乔治, 陈嘉悦, 王楠, 等. 基于MSPA和电路理论的京津冀城市群热环境空间网络[J]. 环境科学, 2023, 44(6): 3034 − 3042. QIAO Zhi, CHEN Jiayue, WANG Nan, et al. Spatial network of urban heat environment in Beijing-Tianjin-Hebei urban agglomeration based on MSPA and circuit theory [J]. Environmental Science, 2023, 44(6): 3034 − 3042. [12] 潘越, 龚健, 杨建新, 等. 基于生态重要性和MSPA核心区连通性的生态安全格局构建——以桂江流域为例[J]. 中国土地科学, 2022, 36(4): 86 − 95. PAN Yue, GONG Jian, YANG Jianxin, et al. Construction of ecological security pattern based on ecological lmportance and connectivity of MSPA-core area: a case study of guifiang river basin [J]. China Land Science, 2022, 36(4): 86 − 95. [13] 刘婷, 欧阳帅, 勾蒙蒙, 等. 基于MSPA模型的新型城市热景观连通性分析[J]. 生态学报, 2023, 43(2): 615 − 624. LIU Ting, OUYANG Shuai, GOU Mengmeng, et al. Analysis connectivity of urban heat island in a new-type urbanization based on MSPA model [J]. Acta Ecologica Sinica, 2023, 43(2): 615 − 624. [14] 刘莉莉. 基于MCR模型的鄠邑区生态安全格局构建[J]. 河南科技, 2023, 42(19): 112 − 116. LIU Lili. Construction of ecological security pattern in Huyi District based on MCR model [J]. Henan Science and Technology, 2023, 42(19): 112 − 116. [15] 李喆, 陈圣宾, 陈芝阳. 地表温度与土地利用类型间的空间尺度依赖性——以成都为例[J]. 生态环境学报, 2022, 31(5): 999 − 1007. LI Zhe, CHEN Shengbin, CHEN Zhiyang. Spatial scale dependence between land surface temperature and land use types: a case study of Chengdu City [J]. Ecology and Environmental Sciences, 2022, 31(5): 999 − 1007. [16] 覃志豪, ZHANG Minghua, ARNON K, 等. 用陆地卫星TM6数据演算地表温度的单窗算法[J]. 地理学报, 2001, 56(4): 456 − 466. TAN Zhihao, ZHANG Minghua, ARNON K, et al. Mono-window algorithm for retrieving land surface temperature from Landsat TM6 data [J]. Acta Geographica Sinica, 2001, 56(4): 456 − 466. [17] 贾海峰, 刘雪华. 环境遥感原理与应用[M]. 北京: 清华大学出版社, 2006. JIA Haifeng, LIU Xuehua. Principles and Applications of Environmental Remote Sensing [M]. Beijing: Tsinghua University Press, 2006. [18] LAL S, SHEEL V. A study of the atmospheric photochemical loss of N2O based on trace gas measurements [J]. Chemosphere-Global Change Science, 2000, 2(3/4): 455 − 463. [19] 贾玉雪, 帅红, 韩龙飞. 基于“源-汇”理论的资江下游地区非点源污染风险区划[J]. 应用生态学报, 2020, 31(10): 3518 − 3528. JIA Yuxue, SHUAI Hong, HAN Longfei. Zonation on non-point source pollution risk in the lower reaches of ZiJiang River based on the “source-sink” theory [J]. Chinese Journal of Applied Ecology, 2020, 31(10): 3518 − 3528. [20] 邵润钰, 罗紫薇, 胡希军, 等. 基于MSPA和MCR模型的株洲市生态网络构建与优化[J]. 西北林学院学报, 2024, 39(2): 217 − 227. SHAO Runyu, LUO Ziwei, HU Xijun, et al. Construction of Zhuzhou ecological network based on MSPA and MCR models [J]. Journal of Northwest Forestry University, 2024, 39(2): 217 − 227. [21] 陈胜兰, 丁山, 魏甫, 等. 基于生态景观连通性的浏阳市自然保护地整合优化评价[J]. 中南林业调查规划, 2023, 42(4): 21 − 25. CHEN Shenglan, DING Shan, WEI Fu, et al. Integrated optimization evaluation of natural protected areas in Liuyang City of Hunan Province based on ecological landscape connectivity [J]. Central South Forest Inventory and Planning, 2023, 42(4): 21 − 25. [22] 颜钰, 曾真, 陈秀铭, 等. 基于MCR模型的泉州市绿地生态网络构建与生态节点识别研究[J]. 山东林业科技, 2024, 54(1): 11 − 20. YAN Yu, ZENG Zhen, CHEN Xiuming, et al. Research on the construction of greenland ecological network and ecological nodes identification in Quanzhou city based on MCR modeling [J]. Journal of Shandong Forestry Science and Technology, 2024, 54(1): 11 − 20. [23] 郭家新, 胡振琪, 李海霞, 等. 基于MCR模型的市域生态空间网络构建[J]. 农业机械学报, 2021, 52(3): 275 − 284. GUO Jiaxin, HU Zhenqi, LI Haixia, et al. Construction of municipal ecological space network based on MCR model [J]. Transactions of the Chinese Society for Agricultural Machinery, 2021, 52(3): 275 − 284. [24] 韦宝婧, 苏杰, 胡希军, 等. 基于“HY-LM”的生态廊道与生态节点综合识别研究[J]. 生态学报, 2022, 42(7): 2995 − 3009. WEI Baojing, SU Jie, HU Xijun, et al. Comprehensive identification of eco-corridors and eco-nodes based on principle of hydrological analysis and Linkage Mapper [J]. Acta Ecologica Sinica, 2022, 42(7): 2995 − 3009. [25] 李程蓉, 陈天. 缓解城市热环境的多层级“源-汇”景观网络构建[J]. 生态学报, 2023, 43(8): 3068 − 3078. LI Chengrong, CHEN Tian. Construction of multi-level “source-sink” landscape network to alleviate urban thermal environment [J]. Acta Ecologica Sinica, 2023, 43(8): 3068 − 3078. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230505
 点击查看大图
点击查看大图
计量
- 文章访问数: 2017
- HTML全文浏览量: 980
- PDF下载量: 55
- 被引次数: 0
