




-
群落结构是指群落中各生物在空间和时间上的配置状况[1],主要表现为一定的种类成分、垂直和水平结构以及群落的外貌[2]。不同群落类型的组成、结构、功能和动态方面表现出很大差异。研究海岛植被群落结构特征,对认识海岛植被具有一定的参考价值,对海岛植被恢复与保护、理解地理隔离对物种和群落类型分布等具有重要理论意义。海域隔离以及和陆地间的岛屿模型联系,使海岛作为一个较为独特和脆弱的地貌单元,形成了特殊的地貌景观。海岛上,物种的独立进化一般使海岛具有一定数量的特有种和主要群落。另外,相比大陆,受洋流、季风、人为等干扰的影响,海岛植被群落较不稳定,群落结构易发生变化。关于亚热带海岛植被的研究,陈征海等[3-6]对浙江海岛的植物区系、海岛砂生植被、海岛盐生植被、海岛乡土树种的基本特征进行了深入的研究,也分别对海岛盐生植被、砂生植被、乡土树种资源进行了详细的调查统计,指出浙江海岛植物区系组成复杂,地理成分多样,纬向性明显,与邻近地区有着不同程度的联系,但有关普陀山植被资源整体认识的研究还比较欠缺。普陀山是亚热带常绿阔叶林海岛型地带性植被的典型代表,是中国四大佛教名山之一,也是东部区域经济、政治、文化、宗教影响力最强的岛屿。据记载,普陀山拥有台湾蚊母树Distylium gracile林、檵木Loropetalum chinensis林、红山茶Camellia longicaudata林等特殊的群落类型,也有樟树Cinnamomum camphora和铁冬青Ilex rotunda等性状变异较大的树种。普陀山于20世纪60年代开始大规模移植黑松Pinus thunbergii,至20世纪90年代,黑松在普陀山植被群落中成为优势种,占据主导地位。这使得当地主要林型分布面积缩减,多样性降低。然而,1992年后,松材线虫Bursaphelenchus xylophilus病在舟山地区爆发,进而影响到普陀山[7-8],致使黑松大面积死亡,群落结构趋于简单,稳定性下降。鉴于保护当地植被、增强景观质量的要求,以及在适地适树、近自然造林、演替规律的理念下,普陀山园林管理中心自1996年起,大规模砍伐感染松材线虫病的黑松。现在,黑松分布已经很少,而一些青冈Cyclobalanopsis glauca林、檵木林、樟树林、枫香Liquidambar formosana林等主要林型随森林演替又逐渐形成了一定的外貌和规模。但是,经查证,截至目前,有关演替过程中林型结构的变化,以及未受人工干扰前普陀山拥有的台湾蚊母树林、檵木林等特殊群落类型,仅有普陀山植物分类学[9-11],以及很少群落结构和多样性范畴的研究[7-8, 12-13]。对于大规模砍伐黑松后出现的主要林型群落结构的研究却未见报道。因此,系统了解普陀山主要林型的群落结构特征,对解决普陀山植被恢复与保护、群落结构优化、专题风景林的构建等问题具有重要的理论和现实意义。
-
普陀山为国家5A级重点风景名胜区,属浙江省舟山市,位于钱塘江口、舟山群岛东南部海域,总面积为12.5 km2,最高峰佛顶山海拔292 m。普陀山属于亚热带海洋性季风气候,四季分明,气候湿润,雨水充沛,阳光充足,素有“海岛植物园”之称。普陀山植被在中国和浙江植被划分中属“中亚热带常绿阔叶林北部亚地带”的浙闽山丘甜槠Castanopsis eyrei-木荷Schima superba林区(IVAiia-2)中天台、括苍山山地丘陵、岛屿植被片(IVAiia-2U)。普陀山因人类活动较早,原始植被破坏严重,仅寺院附近幸存一批典型的常绿阔叶林和大树、古树。20世纪70年代后,加强植被保护,次生阔叶林恢复较快,现成为浙江沿海岛屿中植被最丰富的岛屿之一。
-
2014年10-12月用法瑞学派的典型样地调查法对普陀山植被进行了全面踏查。针对调查得到的所有群落类型,基于群落内建群种的独特性、海岛型植被的分布特点、各群落类型在普陀山的分布区域面积和频度,从所有群落中遴选出佛顶山台湾蚊母树林、天竺山庄青冈林、正趣亭樟树林、法雨寺青冈幼林、半山庵枫香林、炸药库檵木林、逸云庵罗汉松Podocarpus macrophyllus林、善财洞红山茶林为普陀山主要林型,代表了普陀山亚热带常绿阔叶林、针阔混交林、针叶林等典型植被类型。因青冈是普陀山典型的常绿阔叶林地带性植物,分布广泛,不同地段的林型处于不同的发育阶段,因此,为了研究不同发育阶段的青冈林的群落结构特征,我们选择了位于不同地点的青冈林和青冈幼林。
对于遴选得到的主要林型,在林内建立3个20 m × 20 m的标准固定样地(罗汉松林和红山茶林分布面积较小,仅各建立1个样地,表 1)。按照国家长期生态研究技术规范,对每个样地进行石桩标记(按照10 m × 10 m区分),再按英美学派的每木调查法详细记录样地内所有胸径(diameter at breast height, DBH)≥1 cm的木本植物的物种名称、基径、胸径、树高、枝下高、叶下高、树冠面积、空间坐标等,并对调查的每木个体进行挂牌标记。同时记录样地的环境特征,包括地形、地貌、海拔、坡向、坡位等。此外,还估测了林内乔木层和灌木层的平均高度和平均郁闭度。
表 1 浙江普陀山主要林型的样地特征
Table 1. Plot characteristics of main forests in Mount Putuo of Zhejiang Province
主要林型 坡度/(°) 坡向 地点 样方大小/(m×m) 乔木层郁闭度/% 乔木层平均高度/m 灌木层郁闭度/% 灌木层平均高度/m 红山茶林 24 东南 善财洞 20×20 70 8.9 30 4.2 10 东 天竺山庄 20×20 60 8.5 40 2.8 青冈林 15 东北 天竺山庄 20×20 75 9.5 30 4.3 15 东北 天竺山庄 20×20 55 8.5 35 4.0 20 南 法雨寺旁 20×20 70 5.5 30 2.8 青冈幼林 20 南 法雨寺旁 20×20 75 6.4 45 3.4 13 南 法雨寺旁 20×20 78 7.8 65 2.7 台湾蚊 8 东北 佛顶山 25×50 80 9.6 20 3.7 母树林 23 正东 慧济禅林处 20×20 82 9.5 65 3.2 15 西南 慧济寺墙后 10×40 83 9.3 5 2.7 3 东南 正趣亭 20×20 60 8.5 45 4.5 樟树林 3 东南 正趣亭 20×20 65 9.0 40 5.0 3 西南 正趣亭 20×20 45 7.8 65 4.5 19 东南 半山庵 20×20 70 11.0 40 4.0 枫香林 19 东南 半山庵 20×20 30 13.5 40 3.3 18 东 半山庵 20×20 70 10.5 35 3.5 櫃木林 30 南 金沙山顶坡 10×10 50 3.5 10 西北 炸药库 10×10 10 6.0 75 4.0 罗汉松林 8 东北 逸云庵 30×30 63 10.5 17 2.7 -
本研究从主要林型的林分结构、胸径级分布、群落重要值、物种丰富度、Shannon-Wiener指数、Pielou均匀度指数、平均密度、林型纵横向营养投资权衡关系等来分析普陀山主要林型的群落结构特征。具体计算方法如下。
$$\text{相对密度=}\left( \text{某一种的个体数}/\text{全部种的个体数} \right)\times 100\text{ }\!\!%\!\!\text{ }。$$ (1) $$\text{相对盖度=}\left( \text{某一种的所有植株的盖度之和}/\text{所有种的盖度之和} \right)\times 100\text{ }\!\!%\!\!\text{ }。$$ (2) $$\text{相对频度=}\left( \text{某一种的频度之和}/\text{全部种的频度之和} \right)\times 100\text{ }\!\!%\!\!\text{ }。$$ (3) $$\text{重要值=}\left( \text{相对密度}+\text{相对盖度+相对频度} \right)/3。$$ (4) Shannon-Wiener指数:表示样方中总物种个体数及其每个物种分布的均匀程度,考虑了群落内树种的分散程度。计算公式为:
$$H'-\sum{\left| {{n}_{i}}/N\lg \right.\left. \left( {{n}_{i}}/N \right) \right|}。$$ (5) 式(5)中:ni代表第i种的个体数目,N代表群落中所有个体数目。
Pielou指数:指群落的实测多样性与最大多样性(即物种数相同的情况下完全均匀群落的多样性)之比,可以检验群落水平的相似度,计算公式为:
$${{J}_{\text{h}}}=H'/\ln S。$$ (6) 式(6)中:H′指Shannon-Wiener指数,S为物种总数。
林型纵横向营养投资权衡关系是表征群落结构的一个重要方面,反映了植物在纵横向生长上营养投资的权衡策略。纵向投资策略是指植物在垂向生长上表现出的外貌形态属性,譬如树高[14];相对而言,在水平方向上表现出的外貌形态为植物横向投资,可用植物胸径、树冠面积等衡量[14]。对于普陀山海岛型植被,常年受季风影响,为增加抗风胁迫能力,植物通常采取增加横向生长,减小纵向生长的投资策略来增加自身对海岛环境的适应性。
数据分析中,首先计算主要林型内物种的重要值以及优势种的平均高度和平均胸径;其次,采用胸径级分布的方法来分析普陀山主要林型的群落结构特征,将各林型内所有胸径≥1 cm的个体以8 cm为径级区间统计个数,分析其径级结构;再者,用丰富度、平均密度、Shannon-Wiener指数、Pielou均匀度指数等进一步探究主要林型的群落结构特征;最后,以树高和树冠面积分别作为植物纵横向营养投资策略指标,分析其纵横向营养投资策略, 并用回归分析法拟合树高与胸径和树冠面积间的关系。Shannon-Wiener指数、Pielou均匀度指数在R 3.2.3中计算,树高与胸径和树冠面积的拟合关系以及绘图在Origin 8.5中完成。
-
本研究对普陀山主要林型的标准固定样地进行了每木调查,总计2 884株,隶属79个物种。常见乔木物种有枫香、青冈、樟树、台湾蚊母树、罗汉松、麻栎Quercus acutissima等,灌木物种有檵木、红山茶、柃木Eurya japonica和四川山矾Symplocos setchuensis等。
主要林型优势种特征(表 2)看出,枫香林以枫香、麻栎、樟树、四川山矾等为主,枫香和麻栎占60.00%;红山茶林的优势种为红山茶,普陀樟Cinnamomum pedunculatum,柃木和柞木Xylosma racemosum,其中红山茶的重要值达到了30.21%;檵木林的优势种有檵木、柃木和白栎Quercus fabri,其中檵木重要值占41.23%;罗汉松、樟树和珊瑚树Viburnum odoratissimum为罗汉松林的优势种,其中罗汉松对整个林分的贡献为40.05%;青冈林的优势种有青冈、杨梅Myrica rubra和石栎Lithocarpus glaber等,其中青冈占45.06%;青冈幼林中,青冈的重要值达到了69.02%;台湾蚊母树林中,台湾蚊母树和天仙果Ficus erecta重要值共占50%以上,还有青冈和红楠Machilus thunbergii等;樟树林的优势种是樟树、枫香和柃木,其中樟树的重要值为37.40%。整体上,各林型中优势种所占比例较大,林分结构较稳定。
表 2 浙江普陀山主要林型优势物种特征
Table 2. Dominant species characteristic of main forests in Mount Putuo of Zhejiang Province
主要林型 优势物种 平均树高/m 平均胸径/cm 重要值/% 枫香林 枫香 4.31±2.60 6.09±5.66 32.37 麻栎 9.15±3.69 21.57±9.86 27.56 樟树 13.17±2.50 30.02±12.63 11.02 四川山矾 5.20±1.37 10.65±4.36 3.77 红山茶林 红山茶 3.96±0.77 6.21±4.31 30.21 普陀樟 5.10±1.65 13.31±11.48 15.07 柃木 3.30±0.73 4.71±4.75 10.22 柞木 4.42±1.48 10.83±11.22 9.19 櫃木林 櫃木 2.97±0.63 4.49±4.65 41.23 柃木 3.55±0.64 6.17±3.04 24.09 白栎 4.46±0.52 9.39±2.46 4.59 罗汉松林 罗汉松 3.61±3.68 12.68±23.42 40.05 樟树 7.72±5.46 31.56±36.20 16.50 珊瑚树 3.09±0.76 2.46±1.47 8.73 枫香 5.55±3.43 12.31±16.24 7.62 青冈林 青冈 5.59±1.99 6.48±4.31 45.06 石栎 5.66±2.02 8.48±6.15 16.19 櫃木 3.60±1.64 3.21±1.94 7.12 青冈幼林 青冈 4.42±1.70 5.33±3.39 69.02 杨梅 6.12±0.86 12.22±4.35 6.51 石栎 6.39±1.59 8.97±7.89 5.29 台湾蚊母树林 台湾蚊母树 6.20±3.35 18.44±18.99 38.50 天仙果 3.84±1.28 3.66±2.15 16.16 青冈 6.46±1.99 9.77±6.73 9.25 红楠 6.02±2.94 11.70±11.77 7.90 樟树林 樟树 6.42±2.81 17.68±16.68 37.40 枫香 4.63±2.81 7.32±7.96 7.72 柃木 2.39±0.93 2.24±3.03 6.53 -
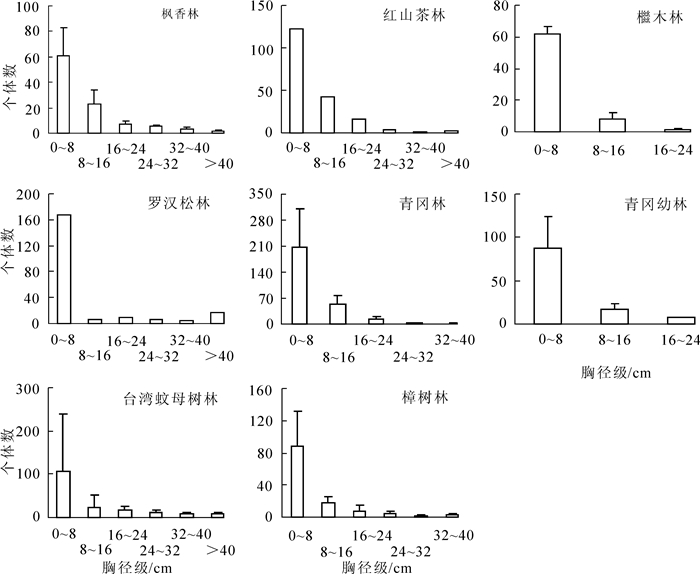
群落的胸径级分布特征也是表征群落结构的主要指标之一[8, 15-16]。普陀山主要林型胸径级分布如图 1所示,枫香林、红山茶林、青冈林、台湾蚊母树林、樟树林的胸径级分布呈“L”型;檵木林、青冈幼林的胸径级分布为单柱型;罗汉松林胸径级分布属于“U”型。胸径在0~8 cm分布个体数最多的是青冈林(205株),最少的是枫香林(61株);胸径>40 cm个体数最多的是罗汉松林(17株),檵木林、青冈林、青冈幼林没有胸径>40 cm的个体。

图 1 浙江普陀山主要林型的胸径级分布
Figure 1. DBH-class distribution of main forests in Mount Putuo of Zhejiang Province
-
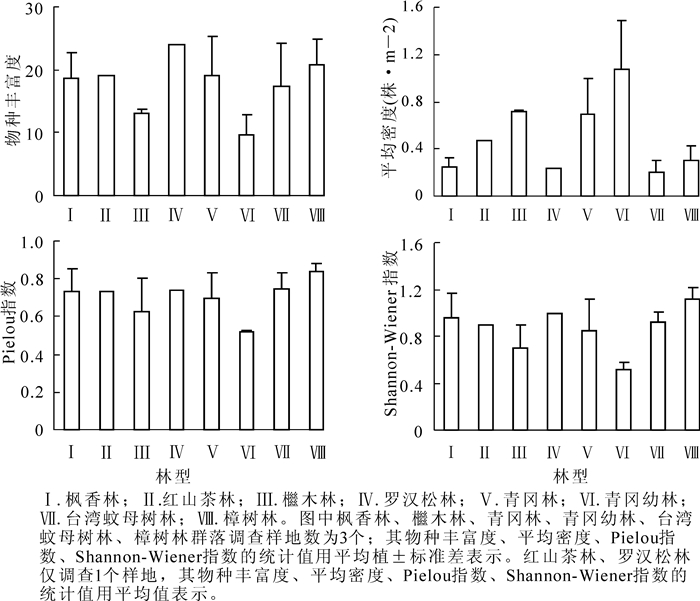
普陀山主要林型多样性特征显示(图 2),物种丰富度依次为罗汉松林>樟树林>青冈林>红山茶林>枫香林>台湾蚊母树林>檵木林>青冈幼林。罗汉松林的物种丰富度最大,为24,青冈幼林最小,为10;植株的平均密度表现为青冈幼林>檵木林>青冈林>红山茶林>樟树林>枫香林>罗汉松林>台湾蚊母树林。青冈幼林的平均密度最大,为1.07株·m-2,台湾蚊母树林最小,为0.20株·m-2;Pielou指数最大和最小的林型分别是樟树林和青冈幼林,分别为0.84和0.52;Shannon-Wiener指数最大的是樟树林,为1.19,最小的是青冈幼林,为0.52。

图 2 浙江普陀山主要林型的物种多样性特征
Figure 2. Species diversity characteristics of main forests in Mount Putuo, Zhejiang Province
-
以树高/胸径、树冠面积/胸径衡量植物在纵向和横向生长上的营养投资策略,结果显示(表 3):枫香林在纵向生长上营养投资最大(μ=1.24, σ2=4.26),红山茶林最小(μ=0.84, σ2=0.54);罗汉松林在横向生长上营养投资最大(μ=1.20, σ2=4.8),青冈林最小(μ=0.48, σ2=0.22)。青冈林和青冈幼林在纵向生长上营养投资显著大于横向生长营养投资(P<0.05),台湾蚊母树林在营养投资策略上,横向显著大于纵向(P<0.05),其他林型在横向和纵向生长上营养投资差异不显著(P>0.05)。
表 3 主要林型纵横向营养投资策略
Table 3. Nutritional investment strategies in vertical and horizontal direction of main forests
主要林型 树高/胸径 树冠面积/胸径 均值(μ) 方差(σ2) 均值(μ) 方差(σ2) 枫香林 1.24 4.26 1.15 1.04 红山茶林 0.84 0.54 0.70 0.17 櫃木林 0.88 0.37 0.64 0.22 罗汉松林 1.21 1.06 1.20 4.80 青冈林 1.08 0.36 0.48 0.22 青冈幼林 1.15 0.62 0.50 0.27 台湾蚊母树林 0.87 0.24 1.12 1.52 樟树林 1.17 1.14 0.94 1.95 树高、胸径和树冠面积间的关系乘幂函数拟合较好(表 4),其中,树高(y)与胸径(x)满足y=2.84(x-0.89)0.35(R2=0.71, P<0.01);胸径(x)与树冠面积(z)的拟合曲线为:y=3.3z0.29(R2=0.51, P<0.01)。
表 4 树高与胸径和树冠面积间的曲线拟合
Table 4. Curve fitting between tree height, DBH and crown area
主要林型 相关因子 曲线方程 枫香林 树高(y)与胸径(x) y=1.52(x+0.27)0.6(R2=0.81, P<0.01) 树高(y)与树冠面积(z) y=2.35(z-0.11)0.45(R2=0.55, P<0.01) 红山茶林 树高(y)与胸径(x) y=5.60+(5.99+4.22)/[1+exp((x+2.68)/5.51)] (R2=0.54, P<0.01) 树高(y)与树冠面积(z) y=5.92/(1+1.14exp(-0.27z)) (R2=0.43, P<0.01) 櫃木林 树高(y)与胸径(x) y=2.7(x-1.43)0.16(R2=0.38, P<0.01) 树高(y)与树冠面积(z) y=4.12+[(2.84-4.11)/(1+exp((z-5.16)/0.94))] (R2=0.22, P<0.01) 罗汉松林 树高(y)与胸径(x) y=10.97+(10.97+1.30)/[1+exp((x-7.64)/6.65)] (R2=0.86, P<0.01) 树高(y)与树冠面积(z) y=2.08+0.33z-0.002z2(R2=0.70, P<0.01) 青冈林 树高(y)与胸径(x) y=8.41+(8.41+5.67)/[1+exp((x-0.11)/4.10)] (R2=0.70, P<0.01) 树高(y)与树冠面积(z) y=3.86+0.48z-0.01z2(R2=0.40, P<0.01) 青冈幼林 树高(y)与胸径(x) y=0.45+0.34x+0.13x2(R2=0.59, P<0.01) 树高(y)与树冠面积(z) y=3.73+0.35z(R2=0.36, P<0.01) 台湾蚊母树林 树高(y)与胸径(x) y=3.12(x-0.94)0.31(R2=0.83, P<0.01) 树高(y)与树冠面积(z) y=3.26(z-0.7)0.28(R2=0.71, P<0.01) 樟树林 树高(y)与胸径(x) y=9.31+(-584.94-9.31)/[1+exp((x+48.16)/11.21)](R2=0.81, P<0.01) 树高(y)与树冠面积(z) y=2.46(z+0.11)0.40(R2=0.62, P<0.01) 整体 树高(y)与胸径(x) y=2.84(x-0.89)0.35(R2=0.71, P<0.01) 树高(y)与树冠面积(z) y=3.3z0.29(R2=0.51, P<0.01) -
普陀山主要林型中,枫香林的乔木层平均高度最大,檵木林最小。因为枫香林的树木高大,长势良好,分布在山坳,林内均匀分布有大径级的枫香、樟树、麻栎等,林分结构稳定;檵木林分布在山坡,林分结构单一,受海风胁迫较大,稳定性较差。樟树林的灌木层平均高度最大,罗汉松林最小。因樟树林的灌木层主要由柃木、天仙果等组成,结构紧密,灌木相对较高。罗汉松林的灌木层主要由分布在古树下的幼树和小灌木组成,整体较矮。台湾蚊母树林的乔木层郁闭度最高,因其内分布一些大径级的古树,灌木稀少,林下空旷,大树分布均一。樟树林的乔木层郁闭度最低,因林内乔木主要是樟树,分布稀疏,树冠面积较小。青冈幼林的灌木层郁闭度最大,台湾蚊母树林最小,因青冈幼林中以青冈灌丛为主,结构紧密,而台湾蚊母树林灌木稀少,分布稀疏,林下空旷。
物种组成上,主要林型的物种与大陆相似,属于亚热带常绿阔叶林,以壳斗科Fagaceae和樟科Lauraceae占优势[13]。与邻近的天童地区植物组成相比,天童主要以木荷、石栎为主,而在普陀山,木荷未有天然分布群落,石栎有较少分布。这可能源于普陀山离大陆较远,属于物种消失的边缘。再加上地理隔离和种子扩散限制,可能使种子未扩散至普陀山[15]。另外,普陀山主要林型在天童山未有成片分布。
主要林型间优势种的差异较大,枫香林位于山坳中,以高大的枫香、麻栎和樟树为主;红山茶林分布在山坡上,以红山茶、普陀樟、柃木等低矮、耐旱的物种为主;檵木一般与柃木伴生出现,分布在养分贫瘠、风力较大的山脊,植株紧密,抗风性较好,但景观效果较差;罗汉松林分布在湿润的低地;高大的罗汉松、樟树古树下面,分布有珊瑚树、枫香等较耐阴物种的幼树;青冈和青冈幼林的优势种是青冈、石栎和檵木,青冈和石栎是伴生种;台湾蚊母树林的优势种是台湾蚊母树,其林分结构单一,物种稀少;樟树林的优势种有樟树、枫香和柃木。
普陀山主要林型的胸径级分布结果显示,枫香林、红山茶林、青冈林、台湾蚊母树林、樟树林胸径级分布呈“L”型,表现为从小胸径到大胸径个体数连续减小的趋势,群落处于发展期,群落结构稳定。林内有少部分大径级的枫香、麻栎、樟树、台湾蚊母树等,有一小部分中径级的柃木、四川山矾、普陀樟、柞木等,以及大部分小径级的胡颓子Elaeagnus pungens,栀子Gardenia jasminoides,茶Camellia sinensis,天仙果,珊瑚树等组成,林型属于生长型。檵木林、青冈幼林属于单柱型,大径级或小径级的树种分布较多,缺乏中等径级的乔木或亚乔木,没有林冠层,空间资源没能充分利用。这些林型由于建群种的缺失,其抵抗性差、林分结构单一,生长力较差,结构不太稳定。罗汉松林胸径级分布属于“U”型,其内大径级和小径级的树种分布较多,大径级树种以罗汉松和樟树古树为主,几乎没有亚乔木,林下分布较多小径级的珊瑚树、青冈幼树,高大乔木和林下幼树的数量之比为1∶3,说明其发育过程中受到严重干扰,古树竞争力弱,林下小苗多,更新快,一直没能形成亚乔木层,群落结构不太稳定。
-
物种多样性反映了群落中物种的丰富程度及群落的稳定性与动态,是群落组织结构的重要特征之一。普陀山主要林型中,罗汉松林的物种丰富度最大,青冈幼林最小,因为罗汉松林物种丰富、植株高大、较多大径级古树均匀分布,其林下更新层幼树较少;而青冈幼林相对物种单一,青冈所占比例较大,主要分布在相对贫瘠的山坡上,由于种间竞争和生境的限制,所以青冈幼林中物种丰富度较低。台湾蚊母树林的平均密度最小,青冈幼林最大,因台湾蚊母树林物种分布稀疏,主要分布在受风胁迫较大的佛顶山,林下更新层植被极少;而青冈幼林物种成簇分布,植株径级较小,分布密集,因而平均密度大。Shannon-Wiener指数表示植株的分散程度,其值越大,说明植株分散程度越高。青冈幼林的Shannon-Wiener指数最小,樟树林最大,说明青冈幼林植株分布最密集,均质性最低。樟树林内物种分布均匀且稀疏,与同纬度的天童山常绿阔叶林的物种多样性相比,天童山的Shannon-Wiener指数为3.3~4.2,而普陀山主要林型的Shannon-Wiener指数只有0.52~1.19。同样,Pielou指数(0.52~0.84)也相对天童山(0.87~0.89)较小[17],说明海岛植被物种多样性相对大陆较低,这与其生境有关,因普陀山常年受海风干扰。此外,普陀山主要林型间物种多样性差异较大,因为这些林型属于亚热带常绿阔叶林,其物种多样性远比落叶阔叶林间的差异大[18]。
-
植物纵横向营养投资权衡策略是衡量林分结构和功能的一个重要指标。蒋子涵等[15]研究了择伐对阔叶红松Pinus koraiensis林主要树种横向和纵向生长的影响。结果显示:择伐使林分稀疏,显著减小了主要树种的树高胸径比(P<0.01), 即林分稀疏的森林,植物在横向生长上营养投资较多,表现为植物低矮,冠幅较大。本研究的各林型中,枫香林在纵向生长上营养投资最大,红山茶林最小。主要是枫香林以高大的枫香为主,其高度较大,树冠面积相对较小,其将营养大部分用于纵向生长上。而红山茶林以低矮的红山茶为优势种,整体较矮,树冠相对其高度的营养投资占优。在植物横向生长营养投资方面,罗汉松林最大,青冈林最小。因罗汉松林以大径级的古树占主导,罗汉松古树有17株,树龄均在300 a以上,平均胸径为51.8 cm,最大达120.0 cm,长势良好[19]。其枝干向四周延伸,树冠面积较大,冠径能达到8~10 m,其在横向生长方面营养投资相对较大。而青冈林径级整体较小,树冠面积较小,大多数青冈成簇分布,其横向生长营养投资较小。整体上,抗风是普陀山植物的主要功能,大多数植物为适应强风,以横向生长投资策略为主,而生长在山坳中的物种以纵向营养投资为主。由树木的胸径、树高、树冠面积的相互关系得出:胸径、树高、树冠面积间存在显著的正相关关系[16, 20],而生境优越、避风性较好的山坳中的稀疏林分中,植物的横向生长率与树高、胸径之间存在显著相关性(P<0.05)[14]。表 4所示的方程也证明了它们间的正相关关系。对主要林型的胸径、树高、树冠面积分别进行曲线拟合,彼此间有显著的正相关关系(P<0.01)。拟合发现可以用乘幂曲线模型y=axb很好地拟合这三者间的关系(P<0.01),在实际操作中可以作为应用模型,只精确测量树木的胸径,就可以估测出树木的高度和树冠面积。
-
普陀山主要林型中,红山茶林、台湾蚊母树林、罗汉松林属于寺庙附近幸存的古树群落,而枫香林、檵木林、青冈林、青冈幼林、樟树林等是大规模砍伐黑松后形成的具有一定群落外貌的林型。这些林型是普陀山亚热带常绿阔叶林的典型代表。主要林型除罗汉松林外,其他林型结构稳定、径级分布合理、生长力旺盛,物种多样性较丰富,各林型中物种呈均匀分布,且林型间差异不大。普陀山主要林型由于常年受海风胁迫,相对于内陆地区,植物在横向生长上营养投资相对较大。总之,对普陀山主要林型群落结构综合特征的研究,对进一步认识普陀山植被、了解植物对海岛环境的适应策略具有重要的意义。
Community structure for main forests in Mount Putuo, Zhejiang Province
-
摘要: 群落结构是植物群落研究的核心内容,对于指导营林建设、植被恢复与保护、群落结构优化、专题风景林构建具有重要的理论和现实意义。以浙江普陀山主要林型枫香Liquidambar formosana林、红山茶Camellia longicaudata林、檵木Loropetalum chinensis林、罗汉松Podocarpus macrophyllus林、青冈Cyclobalanopsis glauca林、青冈幼林、台湾蚊母树Distylium gracile林、樟树Cinnamomum camphora林等为研究对象,分析群落结构特征。结果如下:①林分结构上,各林型外貌相对完整,优势种所占比例较大,林分结构相对稳定;②胸径级分布看,枫香林、红山茶林、青冈林、台湾蚊母树林、樟树林呈“L”型;檵木林、青冈幼林为单柱型;罗汉松林为“U”型;③物种多样性上,罗汉松林的物种丰富度最大,青冈幼林的物种丰富度、Pielou指数、Shannon-Wiener指数最小,平均密度最大;台湾蚊母树林的平均密度最小,樟树林的Pielou指数和Shannon-Wiener指数最大。④植物纵横向营养投资上,枫香林在纵向生长上营养投资最大,红山茶林最小;罗汉松林在横向生长上营养投资最大,青冈林最小。青冈林和青冈幼林在纵向生长上营养投资显著大于横向生长(P<0.05),台湾蚊母树林在横向生长上营养投资显著大于纵向生长(P<0.05)。树高与胸径(R2=0.71, P<0.01)和树冠面积(R2=0.51, P<0.01)的关系幂函数拟合较好。总体上,普陀山主要林型林分结构较稳定,胸径级分布较合理,物种多样性较丰富,抗风胁迫能力较强,但还需进一步优化群落结构,增加物种多样性,提高植物对海岛环境的适应性。Abstract: Community structure is a theoretically and practically important plant community research field in areas of forest development, vegetative restoration and protection, community structure optimization, and special scenic spot development. In this study, forest types of Liquidambar formosana, Camellia longicaudata, Podocarpus macrophyllus, Cyclobalanopsis glauca, young Cyclobalanopsis glauca, Loropetalum chinensis, Distylium gracile, and Cinnamomum camphora were selected to analyze their structural characteristics in subtropical forests of Zhoushan Island, Zhejiang Province. Analysis included Pielou and Shannon-Wiener indices and power-fitted curves for DBH and tree crown. Results showed that stand structure and forest appearance were relatively complete indicating that forest types were dominant species and stand structure was relatively stable. With respect to diameter-class distribution, “L” type forests were Liquidambar formosana, Camellia longicaudata, Cyclobalanopsis glauca, D. gracile, and Cinnamomum camphora; whereas, unimodal type forests were Loropetalum chinensis and young Cyclobalanopsis glauca, with P. macrophyllus forest as a “U” type for diameter-class distribution. With respect to biodiversity indices, the P. macrophyllus forest had the highest species richness; the young Cyclobalanopsis glauca forest had the lowest species richness, Pielou and Shannon-Wiener indices but highest average stem density; the D. gracile forest had the lowest average stem density; and the Cinnamomum camphora forest showed the lowest Shannon-Wiener and Pielou indices. Nutrition investment in the vertical direction, was highest for the Liquidambar formosana forest and lowest in the Camellia longicaudata forest(P<0.05). Horizontal nutrition investment was highest in the P. macrophyllus forest and lowest in the Cyclobalanopsis glauca forest(P<0.05). In the Cyclobalanopsis glauca and the Cyclobalanopsis glauca young forest, vertical nutrition investment was greater than horizontal nutrition investment(P<0.05); the D. gracile forest was reversed(P<0.05). In addition, power curves of tree height fit for each DBH (R2 = 0.71, P<0.01) and crown area (R2 = 0.51, P<0.01) were highly significant. Thus, stand structure in the study area was stable, diameter-class distribution was reasonable, species diversity was high, and wind resistance was strong, but some discrepancies existed meaning more efforts to optimize community structure, increase species diversity, and improve plant adaptation to island habitats were needed.
-
图 1 浙江普陀山主要林型的胸径级分布
枫香林、櫃木林、青冈林、青冈幼林、台湾蚊母树林、樟树林群落调查样地数为3个,其胸径级个体数用平均数±标准差表示,红山茶林、罗汉松林仅调查样地数为1个,其胸径级个体数用平均数表示。
Figure 1 DBH-class distribution of main forests in Mount Putuo of Zhejiang Province
图 2 浙江普陀山主要林型的物种多样性特征
Ⅰ.枫香林;Ⅱ.红山茶林;Ⅲ.櫃木林;Ⅳ.罗汉松林;Ⅴ.青冈林;Ⅵ.青冈幼林;Ⅶ.台湾蚊母树林;Ⅷ.樟树林。图中枫香林、櫃木林、青冈林、青冈幼林、台湾蚊母树林、樟树林群落调查样地数为3个,其物种丰富度、平均密度、Pielou指数、Shannon-Wiener指数的统计值用平均数±标准差表示,红山茶林、罗汉松林仅调查了样地为1个,其物种丰富度、平均密度、Pielou指数、Shannon-Wiener指数的统计值用平均数表示。
Figure 2 Species diversity characteristics of main forests in Mount Putuo, Zhejiang Province
表 1 浙江普陀山主要林型的样地特征
Table 1. Plot characteristics of main forests in Mount Putuo of Zhejiang Province
主要林型 坡度/(°) 坡向 地点 样方大小/(m×m) 乔木层郁闭度/% 乔木层平均高度/m 灌木层郁闭度/% 灌木层平均高度/m 红山茶林 24 东南 善财洞 20×20 70 8.9 30 4.2 10 东 天竺山庄 20×20 60 8.5 40 2.8 青冈林 15 东北 天竺山庄 20×20 75 9.5 30 4.3 15 东北 天竺山庄 20×20 55 8.5 35 4.0 20 南 法雨寺旁 20×20 70 5.5 30 2.8 青冈幼林 20 南 法雨寺旁 20×20 75 6.4 45 3.4 13 南 法雨寺旁 20×20 78 7.8 65 2.7 台湾蚊 8 东北 佛顶山 25×50 80 9.6 20 3.7 母树林 23 正东 慧济禅林处 20×20 82 9.5 65 3.2 15 西南 慧济寺墙后 10×40 83 9.3 5 2.7 3 东南 正趣亭 20×20 60 8.5 45 4.5 樟树林 3 东南 正趣亭 20×20 65 9.0 40 5.0 3 西南 正趣亭 20×20 45 7.8 65 4.5 19 东南 半山庵 20×20 70 11.0 40 4.0 枫香林 19 东南 半山庵 20×20 30 13.5 40 3.3 18 东 半山庵 20×20 70 10.5 35 3.5 櫃木林 30 南 金沙山顶坡 10×10 50 3.5 10 西北 炸药库 10×10 10 6.0 75 4.0 罗汉松林 8 东北 逸云庵 30×30 63 10.5 17 2.7  下载: 导出CSV
下载: 导出CSV
表 2 浙江普陀山主要林型优势物种特征
Table 2. Dominant species characteristic of main forests in Mount Putuo of Zhejiang Province
主要林型 优势物种 平均树高/m 平均胸径/cm 重要值/% 枫香林 枫香 4.31±2.60 6.09±5.66 32.37 麻栎 9.15±3.69 21.57±9.86 27.56 樟树 13.17±2.50 30.02±12.63 11.02 四川山矾 5.20±1.37 10.65±4.36 3.77 红山茶林 红山茶 3.96±0.77 6.21±4.31 30.21 普陀樟 5.10±1.65 13.31±11.48 15.07 柃木 3.30±0.73 4.71±4.75 10.22 柞木 4.42±1.48 10.83±11.22 9.19 櫃木林 櫃木 2.97±0.63 4.49±4.65 41.23 柃木 3.55±0.64 6.17±3.04 24.09 白栎 4.46±0.52 9.39±2.46 4.59 罗汉松林 罗汉松 3.61±3.68 12.68±23.42 40.05 樟树 7.72±5.46 31.56±36.20 16.50 珊瑚树 3.09±0.76 2.46±1.47 8.73 枫香 5.55±3.43 12.31±16.24 7.62 青冈林 青冈 5.59±1.99 6.48±4.31 45.06 石栎 5.66±2.02 8.48±6.15 16.19 櫃木 3.60±1.64 3.21±1.94 7.12 青冈幼林 青冈 4.42±1.70 5.33±3.39 69.02 杨梅 6.12±0.86 12.22±4.35 6.51 石栎 6.39±1.59 8.97±7.89 5.29 台湾蚊母树林 台湾蚊母树 6.20±3.35 18.44±18.99 38.50 天仙果 3.84±1.28 3.66±2.15 16.16 青冈 6.46±1.99 9.77±6.73 9.25 红楠 6.02±2.94 11.70±11.77 7.90 樟树林 樟树 6.42±2.81 17.68±16.68 37.40 枫香 4.63±2.81 7.32±7.96 7.72 柃木 2.39±0.93 2.24±3.03 6.53
下载: 导出CSV
表 3 主要林型纵横向营养投资策略
Table 3. Nutritional investment strategies in vertical and horizontal direction of main forests
主要林型 树高/胸径 树冠面积/胸径 均值(μ) 方差(σ2) 均值(μ) 方差(σ2) 枫香林 1.24 4.26 1.15 1.04 红山茶林 0.84 0.54 0.70 0.17 櫃木林 0.88 0.37 0.64 0.22 罗汉松林 1.21 1.06 1.20 4.80 青冈林 1.08 0.36 0.48 0.22 青冈幼林 1.15 0.62 0.50 0.27 台湾蚊母树林 0.87 0.24 1.12 1.52 樟树林 1.17 1.14 0.94 1.95
下载: 导出CSV
表 4 树高与胸径和树冠面积间的曲线拟合
Table 4. Curve fitting between tree height, DBH and crown area
主要林型 相关因子 曲线方程 枫香林 树高(y)与胸径(x) y=1.52(x+0.27)0.6(R2=0.81, P<0.01) 树高(y)与树冠面积(z) y=2.35(z-0.11)0.45(R2=0.55, P<0.01) 红山茶林 树高(y)与胸径(x) y=5.60+(5.99+4.22)/[1+exp((x+2.68)/5.51)] (R2=0.54, P<0.01) 树高(y)与树冠面积(z) y=5.92/(1+1.14exp(-0.27z)) (R2=0.43, P<0.01) 櫃木林 树高(y)与胸径(x) y=2.7(x-1.43)0.16(R2=0.38, P<0.01) 树高(y)与树冠面积(z) y=4.12+[(2.84-4.11)/(1+exp((z-5.16)/0.94))] (R2=0.22, P<0.01) 罗汉松林 树高(y)与胸径(x) y=10.97+(10.97+1.30)/[1+exp((x-7.64)/6.65)] (R2=0.86, P<0.01) 树高(y)与树冠面积(z) y=2.08+0.33z-0.002z2(R2=0.70, P<0.01) 青冈林 树高(y)与胸径(x) y=8.41+(8.41+5.67)/[1+exp((x-0.11)/4.10)] (R2=0.70, P<0.01) 树高(y)与树冠面积(z) y=3.86+0.48z-0.01z2(R2=0.40, P<0.01) 青冈幼林 树高(y)与胸径(x) y=0.45+0.34x+0.13x2(R2=0.59, P<0.01) 树高(y)与树冠面积(z) y=3.73+0.35z(R2=0.36, P<0.01) 台湾蚊母树林 树高(y)与胸径(x) y=3.12(x-0.94)0.31(R2=0.83, P<0.01) 树高(y)与树冠面积(z) y=3.26(z-0.7)0.28(R2=0.71, P<0.01) 樟树林 树高(y)与胸径(x) y=9.31+(-584.94-9.31)/[1+exp((x+48.16)/11.21)](R2=0.81, P<0.01) 树高(y)与树冠面积(z) y=2.46(z+0.11)0.40(R2=0.62, P<0.01) 整体 树高(y)与胸径(x) y=2.84(x-0.89)0.35(R2=0.71, P<0.01) 树高(y)与树冠面积(z) y=3.3z0.29(R2=0.51, P<0.01)
下载: 导出CSV
-
[1] 陈灵芝. 中国的生物多样性现状及其保护对策[M]. 北京: 科学出版社, 1993. [2] 姜汉侨, 段昌群, 杨树华, 等. 植物生态学[M]. 北京: 高等教育出版社, 2004:49-72. [3] 陈征海, 唐正良, 孙海平, 等. 浙江海岛乡土树种资源调查研究[J]. 浙江林业科技, 1995, 15(6): 1-7. CHEN Zhenghai, TANG Zhengliang, SUN Haiping, et al. Investigation on resources of indigenous plant for forestation in the islands of Zhejiang Province [J]. J Zhejiang For Sci Technol, 1995, 15(6): 1-7. [4] 陈征海, 唐正良, 张晓华, 等. 浙江海岛砂生植被研究(Ⅰ)植被的基本特征[J]. 浙江林学院学报, 1995, 12(4): 388-398. CHEN Zhenghai, TANG Zhengliang, ZHANG Xiaohua, et al. Sand vegetation of the islands in Zhejiang(Ⅰ) basic characteristics of vegetation [J]. J Zhejiang For Coll, 1995, 12(4): 388-398. [5] 陈征海, 唐正良, 张晓华, 等. 浙江海岛盐生植被研究(Ⅰ)植被的基本特征[J]. 生态学杂志, 1996, 15(1): 14-19. CHEN Zhenghai, TANG Zhengliang, ZHANG Xiaohua, et al. A study of the halophilous vegetation on the islands of Zhejiang Province(Ⅰ) basic characteristic of the vegetation [J]. Chin J Ecol, 1996, 15(1): 14-19. [6] 陈征海, 唐正良, 裘宝林. 浙江海岛植物区系的研究[J]. 云南植物研究, 1995, 17(4): 405-412. CHEN Zhenghai, TANG Zhengliang, QIU Baolin. A study on the flora of the islands of Zhejiang Province [J]. Acta Bot Yunnan, 1995, 17(4): 405-412. [7] 钱逸凡, 伊力塔, 胡军飞, 等. 普陀山主要植物种生态位特征[J]. 生态学杂志, 2012, 31(3): 561-568. QIAN Yifan, Yilita, HU Junfei, et al. Niche characteristic of main plant species in Putuo Mountain, Zhejiang Province of East China [J]. Chin J Ecol, 2012, 31(3): 561-568. [8] 徐悦, 钱逸凡, 伊力塔, 等. 普陀山森林群落结构与生物多样性研究[J]. 浙江林业科技, 2013, 33(5): 74-80. XU Yue, QIAN Yifan, Yilita, et al. Study on forest community structure and biodiversity in Putuoshan Island[J]. J Zhejiang For Sci Technol, 2013, 33(5): 74-80. [9] 李根有, 陈征海, 胡军飞, 等. 发现于浙江普陀山岛的2个植物新变种[J]. 浙江林学院学报, 2010, 27(6): 908-909. LI Genyou, CHEN Zhenghai, HU Junfei, et al. Two new plant varieties from Putuo Island, Zhejiang Province [J]. J Zhejiang For Coll, 2010, 27(6): 908-909. [10] 金水虎, 马丹丹, 欧丹燕, 等. 普陀山4个植物新类群[J]. 西北植物学报, 2010, 30(8): 1701-1702. JIN Shuihu, MA Dandan, OU Danyan, et al. New taxa of plant in Putuo Mountain [J]. Acta Bot Boreal-Occident Sin, 2010, 30(8): 1701-1702. [11] 马丹丹, 金水虎, 胡军飞, 等. 发现于普陀山的植物区系新资料[J]. 浙江大学学报:理学版, 2011, 38(2): 215-217. MA Dandan, JIN Shuihu, HU Junfei, et al. Some new floristic findings of Putuoshan Island, Zhejiang [J]. J Zhejiang Univ Sci Ed, 2011, 38(2): 215-217. [12] 庄杰. 舟山海岛典型植物群落结构及优化对策研究[D]. 临安:浙江农林大学, 2012. ZHUANG Jie. The Research of Zhoushan Archipelago Typical Plant Community Structure and Countermeasures of Optimization [D]. Lin'an: Zhejiang A & F University, 2012. [13] 高浩杰, 王国明, 郁庆君. 舟山市种子植物物种多样性及其分布特征[J]. 植物科学学报, 2015, 33(1): 61-71. GAO Haojie, WANG Guoming, YU Qingjun. Distribution characteristic and species diversity of seed plants in Zhoushan, Zhejiang [J]. Plant Sci J, 2015, 33(1): 61-71. [14] 蒋子涵, 金光泽. 择伐对阔叶红松林主要树种径向与纵向生长的影响[J]. 生态学报, 2010, 30(21): 5843-5852. JIANG Zihan, JIN Guangze. Effects of selection cutting on diameter growth and vertical growth among major tree species in the mixed broadleaved-Korean pine forest [J]. Acta Ecol Sin, 2010, 30(21): 5843-5852. [15] 杨同辉, 王良衍. 浙江天童舟山新木姜子群落结构及其更新类型[J]. 浙江林学院学报, 2010, 27(6): 877-883. YANG Tonghui, WANG Liangyan. Community structure and regeneration types of Neolitsea sericea in Tiantong, Zhejiang Province [J]. J Zhejiang For Coll, 2010, 27(6): 877-883. [16] 赵维俊, 刘贤德, 金铭, 等. 祁连山青海云杉林群落结构特征分析[J]. 干旱区研究, 2012, 29(4): 615-620. ZHAO Weijun, LIU Xiande, JIN Ming, et al. Analysis on community structure of Picea crassifolia forests in Qilian Mountains [J]. Arid Zone Res, 2012, 29(4): 615-620. [17] 张光富. 浙江天童山区灌丛群落的物种多样性及其与演替的关系[J]. 生物多样性, 2000, 8(3): 271-276. ZHANG Guangfu. Species diversity of a shrub community in Tiantong region, Zhejiang Province and its implication for succession [J]. Chin Biodiversity, 2000, 8(3): 271-276. [18] 贺金生, 陈伟烈, 李凌浩. 中国中亚热带东部常绿阔叶林主要类型的群落多样性特征[J]. 植物生态学报, 1998, 22(4): 303-311. HE Jinsheng, CHEN Weilie, LI Linghao. Community diversity of the main types of the evergreen broad-leaved forest in the eastern part of the middle subtropical China [J]. Acta Phytoecol Sin, 1998, 22(4): 303-311. [19] 卢小根. 普陀山树木奇观[M]. 北京:中国文联出版社, 2004. [20] 刘翠玲, 潘存德, 梁瀛, 等. 鳞毛蕨(DryoPteris filixmas)天山云杉林种群结构分析[J]. 干旱区研究, 2006, 23(1): 60-65. LIU Cuiling, PAN Cunde, LIANG Ying, et al. Analysis on the population structure of Picea schrenkiana Fisch. Et Mey. -DryoPteris filixmas stand [J]. Arid Zone Res, 2006, 23(1): 60-65. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2016.05.007
 点击查看大图
点击查看大图
计量
- 文章访问数: 5191
- HTML全文浏览量: 1154
- PDF下载量: 363
- 被引次数: 0
