




-
群落是物种与所处生境相互作用的综合。植物群落的物种α多样性与群落稳定性密切相关,体现着群落整体的稳定性[1]。对于濒危植物而言,研究其所在群落的物种组成与结构特征,可以更好地理解其种群动态与生存现状,从而为保护提供科学依据[2]。但传统的物种α多样性研究仅刻画群落中物种数量层面变化,难以反映种间的进化关系及功能差异。近年来,谱系多样性和功能多样性被引入群落生态学研究,用以揭示群落在进化历史和生态功能层面的组织特征,能够更有效地反映群落构建过程[3]。谱系多样性是群落各物种间谱系距离的总和,代表物种间进化差异的累积,有助于揭示群落构建中的进化历史与谱系限制[4−5]。一般认为,在环境压力较大、资源相对有限的生境中,群落共存物种往往在系统发育关系上表现出较高相关性,即呈现谱系聚集格局,这通常被解释为环境过滤作用与系统发育生态位保守性共同作用的结果[6]。相反,谱系过度分散则可能表明物种间竞争排斥或生态位分化占主导地位,系统发育上更相关的物种会更强烈地竞争群落中的有限资源[7−8]。WEBB等[4]在阐述谱系结构研究方法时,指出功能性状的进化特征对群落构建的解释具有重要意义。功能性状作为反映植物资源利用和适应策略的关键属性,反映植物在资源利用、养分权衡方式以及生存和适应的生态策略[9−10]。因此,将物种α多样性与谱系和功能多样性相结合,有助于深化对濒危植物群落结构特征及其生态过程的认识[11]。
蛛网萼Platycrater arguta隶属于绣球花科Hydrangeaceae蛛网萼属Platycrater,为东亚特有的单种属落叶灌木。该物种具有喜湿润环境、种子繁殖困难和种群数量稀少等特点,同时受人类干扰和环境变化的双重威胁,被列为Ⅱ级保护植物[12−13]。生物地理学研究表明:蛛网萼在中国东部与日本南部存在间断分布,其遗传结构与谱系分化受到东海陆桥历史“过滤效应”及第四纪气候波动的显著影响,显示出独特的生物地理格局[14],凸显了该物种的进化与保护价值。目前蛛网萼的研究主要集中在生理机制[15−16]、繁殖生态学[12,17]、线粒体基因组学[18]和野外资源调查与评估[13,19]等方面,但对于蛛网萼所在群落的群落物种α多样性、谱系结构和功能多样性等方面知之甚少。
浙江钱江源-百山祖国家公园候选区百山祖园区(以下简称百山祖国家公园)地处东亚生物多样性热点地区的核心地带,拥有典型的中亚热带常绿阔叶林生态系统,园区内分布有多种国家重点保护植物,是研究濒危物种群落特征与保育策略的理想区域[20]。本研究以百山祖国家公园内蛛网萼天然群落为研究对象,通过样方调查,结合物种α多样性、谱系结构、功能多样性及物种共现网络分析,探讨蛛网萼群落的多样性特征,以期为蛛网萼及其生境的保护提供科学理论依据。
-
百山祖国家公园属中亚热带海洋性季风气候,年均气温约18 ℃,年降水量约1 700 mm。研究区以浙江凤阳山-百山祖国家级自然保护区为基础建立,总面积为50 351.27 hm2,海拔范围为237~1 929 m,保存了浙闽赣交界山地具有代表性和典型性的植被和生态系统,植被类型包括常绿阔叶林、常绿落叶阔叶混交林、针阔叶混交林及高海拔针叶林和灌丛等[21−22]。
-
野外调查于2022年和2023年的8月进行,建立了19个具有代表性的样方(表1),样方设置于蛛网萼天然分布的群落内。除百瀑沟Z01样方面积为10 m × 10 m外,其余样方面积均为20 m × 20 m。按照植物群落清查方法和技术规范[23],记录样方的地理位置、植被类型、生境特征(包括海拔和地形等)、蛛网萼及其他维管植物信息;物种信息包括物种名称、所在层次、多盖度等级(参考Braun-Blanquet分级)[24]、群集度等级、平均高度及最高高度等。
表 1 蛛网萼所处的天然群落样方调查信息
Table 1. General survey information of plots in the natural communities of P. arguta
样方名称 简称 植被类型 样方面积/(m×m) 经度N/(°) 纬度W/(°) 海拔/m 地形 百瀑沟Z01 BPGZ01 针阔混交林 10×10 119.193022 27.751369 1 420 石壁 百瀑沟Z02 BPGZ02 落阔混交林 20×20 119.192653 27.751344 1 401 石壁 百瀑沟Z03 BPGZ03 落阔混交林 20×20 119.191944 27.751228 1 384 石壁 百瀑沟Z04 BPGZ04 常绿阔叶林 20×20 119.189444 27.749722 1 339 石壁 百瀑沟Z05 BPGZ05 常绿阔叶林 20×20 119.189722 27.748417 1 307 石壁 三井溪Z01 SJXZ01 常绿阔叶林 20×20 119.207111 27.795397 1 195 石壁 三井溪Z02 SJXZ02 针阔混交林 20×20 119.207744 27.795544 1 196 石壁 三井溪Z10 SJXZ10 针阔混交林 20×20 119.209306 27.796828 1 194 石壁 三井溪Z12 SJXZ12 针阔混交林 20×20 119.210203 27.800578 1 169 石壁 盘龙地桥边01 PLDQ01 常绿阔叶林 20×20 119.22875 27.764806 1 093 山地 盘龙地桥边02 PLDQ02 针阔混交林 20×20 119.236906 27.765797 1 146 山涧 百管站公路边01 GL01 常绿阔叶林 20×20 119.192439 27.750347 1 416 石壁 半坑01 BK01 常绿阔叶林 20×20 119.150131 27.717856 744 山地 溧阳村01 LYC01 针阔混交林 20×20 119.182414 27.820133 1 042 山地 溧阳村03 LYC03 针阔混交林 20×20 119.203747 27.822078 1 070 山地 久住垟公路边01 JZY01 针阔混交林 20×20 119.248611 27.732456 1 323 山地 哨卡01 SK01 落叶阔叶林 20×20 119.194519 27.909483 1 228 山地 猕猴谷-桑瀑飞雪01 MHG01 常绿阔叶林 20×20 119.194444 27.916667 940 溪沟 炉岙01 LA01 毛竹落叶林 20×20 119.206719 27.916352 1 123 山地 -
样方调查到的物种名称及科、属均依《浙江植物志(新编)》[25]确定。根据浙江种子植物属的分布区类型[26]和吴征镒《中国种子植物属的分布区类型》[27]统计调查群落内种子植物属的区系地理成分。根据《国家重点保护野生植物名录》认定群落中濒危物种的保护等级,木本植物的叶生活型参考《浙江植物志(新编)》[25]。以每个样方群落为1个独立单元,分别计算乔木层、灌木层和草本层物种的相对群集度、相对多盖度、相对高度和重要值(IV)[28]。以重要值为测度依据,计算群落的物种丰富度(S)、Shannon-Wiener多样性指数(Hʹ)、Simpson多样性指数(D)及Pielou均匀度指数(J)[29−30]。
-
为探究蛛网萼种群所处群落物种之间的空间共现模式,找出与蛛网萼具有较强生态共现关系的物种,基于物种存在/缺失矩阵计算所有物种对之间的Jaccard相似性指数构建无向共现网络并计算相关网络拓扑参数[31]。在网络中,每个节点代表1个物种,每条边为相连的2个物种间Jaccard相似性指数的大小,表示这2个物种间的共现程度。仅保留了相似性指数大于0.4的边,以筛选出具有中等及以上共现程度的物种对,共现网络可视化与分析通过R4.3.2和Gephi实现。
-
由于蕨类和裸子植物的谱系枝长过长,且物种数少,谱系和功能多样性研究只关注被子植物。195种被子植物隶属的科属信息使用“物种2000中国节点”(http://sp2000.org.cn)确定,用V.PhyloMaker2软件包[32]内的phylo.maker函数的S3方法,以GBOTB.extended.TPL为骨架生成被子植物谱系树,以Faith谱系多样性指数(PD)测定样方内被子植物的谱系多样性[4]。使用R软件中的Picante软件包计算平均谱系距离(MPD)和平均最近谱系距离(MNTD)。MPD代表了物种间亲缘关系的平均情况,而MNTD侧重指示分支末端的谱系关系,两者可转换成净亲缘关系指数(NRI)和最近种间亲缘关系指数(NTI)。若NRI>0,样方的物种在谱系结构上聚集;若NRI<0,样方的物种在谱系结构上发散;若NRI=0,样方的物种在谱系结构上随机[4,33]。
-
选取9项与植物生存策略密切相关的功能性状评估群落功能多样性:生活型、果实类型、开花期、结果期、授粉方式、种子传播类型、比叶面积(SLA)、植物高度(H)和种子质量(SM)。其中,生活型等6项性状数据参考“植物智”(https://www.iplant.cn/)及《浙江植物志(新编)》[25];植物高度来自实地调查;SLA与SM数据源自中国植物性状数据库V2[34]及TRY数据库[35]。针对SLA与SM的数据缺失,采用R包mice的链式方程多元插补法进行填补[36]。采用重要值作为物种权重,计算4项关键功能多样性指数[37]:功能丰富度(FRic)、功能均匀度(FEve)、功能分异指数(FDiv)以及Rao二次熵指数(RaoQ)。所有功能多样性指数的计算均通过R语言中的FD包实现。
-
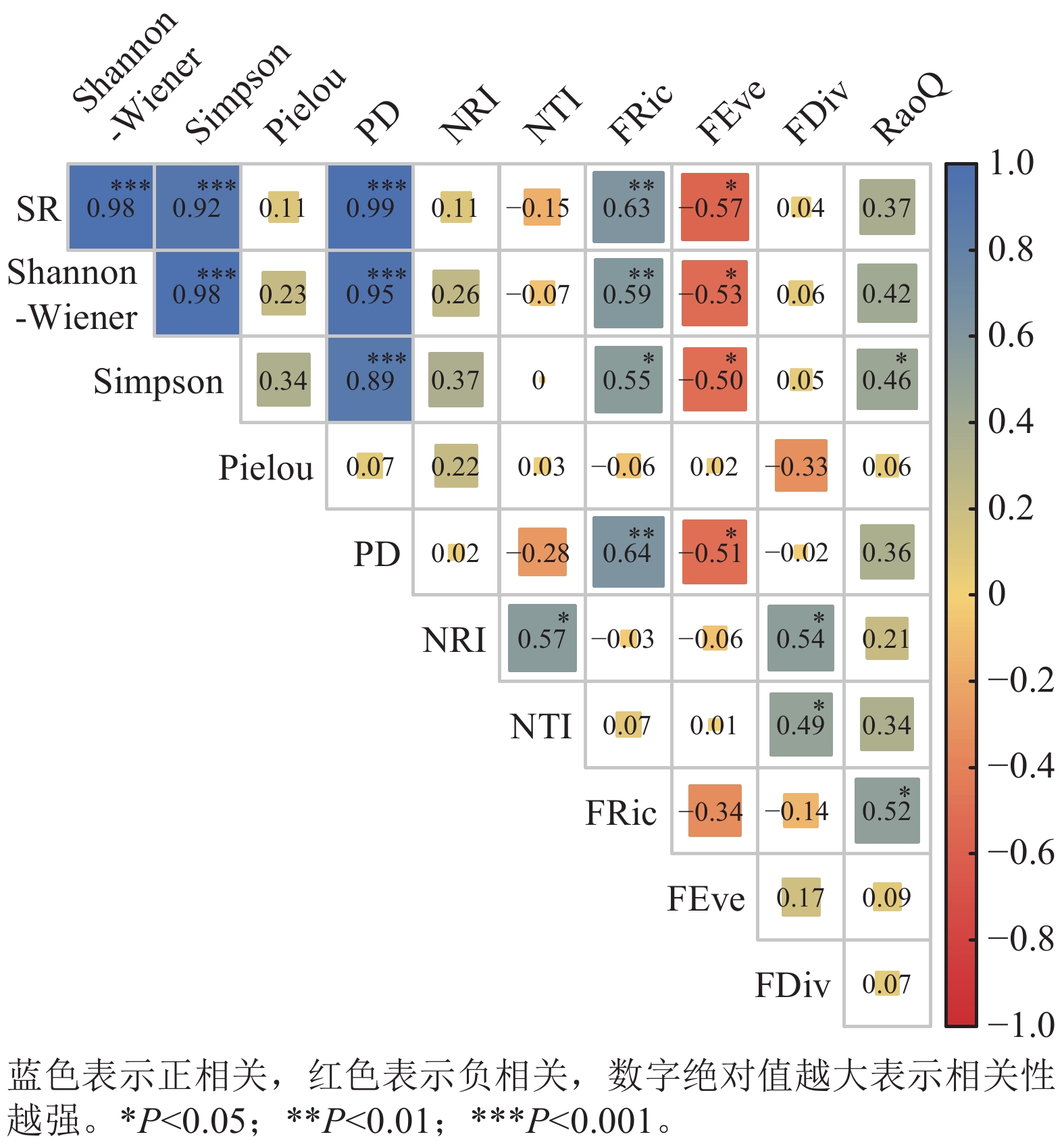
采用Blomberg’s K[38]检验功能性状的系统发育信号强度及显著性(与零模型比较)。K值指示信号强弱:K = 1为性状进化符合布朗运动,即功能性状与系统发育结构无关;K<1为性状进化近随机,系统发育信号弱,群落系统发育结构与性状格局不一致;K>1为强烈的系统发育信号及功能性状保守性,群落系统发育结构与功能性状结构一致[39]。以Pearson相关性检验物种、谱系与功能多样性间相关性。以单因素方差分析比较蛛网萼群落不同层次物种、谱系多样性差异。
-
调查共记录维管植物224种(含变型、变种),隶属于89科162属(附录1)。群落以种子植物为主,共76科145属203种,其中,双子叶植物63科119属171种,占科、属和物种总数的70.79%、73.46%和76.33%;单子叶植物9科19属24种;裸子植物4科7属8种。蕨类植物共记录21种,主要出现于林下湿润生境。外来入侵物种仅菊科Asteraceae小蓬草Erigeron canadensis 1种。生活型组成显示群落以木本植物为主(135种),含常绿阔叶树65种、落叶阔叶树54种、针叶树8种,构成典型的中亚热带森林植被类型。物种组成中,蔷薇科Rosaceae和樟科Lauraceae物种数最多(各13种),其次为杜鹃花科Ericaceae和禾本科Poaceae(各10种)以及山茶科(7种);39科含2~6种,45科仅1种。优势属包括悬钩子属Rubus(7种),润楠属Machilus(6种),冬青属Ilex和杜鹃属Rhododendron(各5种),鳞毛蕨属Dryopteris、柃木属Eurya和山矾属Symplocos(各4种)等。
群落中共记录国家重点保护野生植物9种,Ⅰ级保护有红豆杉科Taxaceae的南方红豆杉Taxus mairei;Ⅱ级保护8种,包括蛛网萼、福建柏Fokienia hodginsii、白豆杉Pseudotaxus chienii、鹅掌楸Liriodendron chinense、闽楠Phoebe bournei、浙江楠Phoebe chekiangensis、中华猕猴桃Actinidia chinensis和独蒜兰Pleione bulbocodioides。
-
以属为基本单位分析19个样方的种子植物区系地理成分,可归为4个大类13个类型(附录2)。其中世界分布类含5属(3.45%):蓼属Polygonum、鼠尾草属Salvia、狸藻属Utricularia、飞蓬属Erigeron和薹草属Carex,均为草本。热带分布类占优势,达62属(42.76%),以热带亚洲分布(20属)和泛热带分布(19属)为主;禾本科(6属8种)和山茶科Theaceae(4属7种)属种较多。温带分布类比例最高,共75属(51.72%),东亚分布(27属)和北温带分布(25属)为核心。中国特有分布类仅3属:杉木属Cunninghamia、白豆杉属Pseudotaxus和车前紫草属Sinojohnstonia。
-
根据样方调查结果中的重要值的大小确定群落内该层次的优势种。基于19个样方统计,蛛网萼在所有调查样方中均有记录,但仅在8个样方中为其所在层次的重要值最高种,表现为群落优势种(附录3)。样方平均物种丰富度29.26种(11~54种)。样方人为干扰小、原生性强,群落层次分明,可分为乔木层(0~18种,平均7.58种)、灌木层(2~21种,平均9.21种)和草本层(4~20种,平均10.42种)。乔木层中主要优势种包括浙闽樱桃Cerasus schneideriana(42.01)、福建柏Fokienia hodginsii(37.39)、小叶青冈Cyclobalanopsis gracilis(34.15)和褐叶青冈Cyclobalanopsis stewardiana(33.15)。灌木层中主要优势种包括蛛网萼(65.87)、中华野海棠Bredia sinensis(61.11)、宜昌荚蒾Viburnum erosum(52.22)和扁枝越橘Vaccinium japonicum var. sinicum(21.87)。草本层中主要优势种包括里白Diplopterygium glaucum(36.05)、盾蕨Lepisorus ovatus(33.77)、芒Miscanthus sinensis(33.09)和华东瘤足蕨Plagiogyria japonica(29.43)。
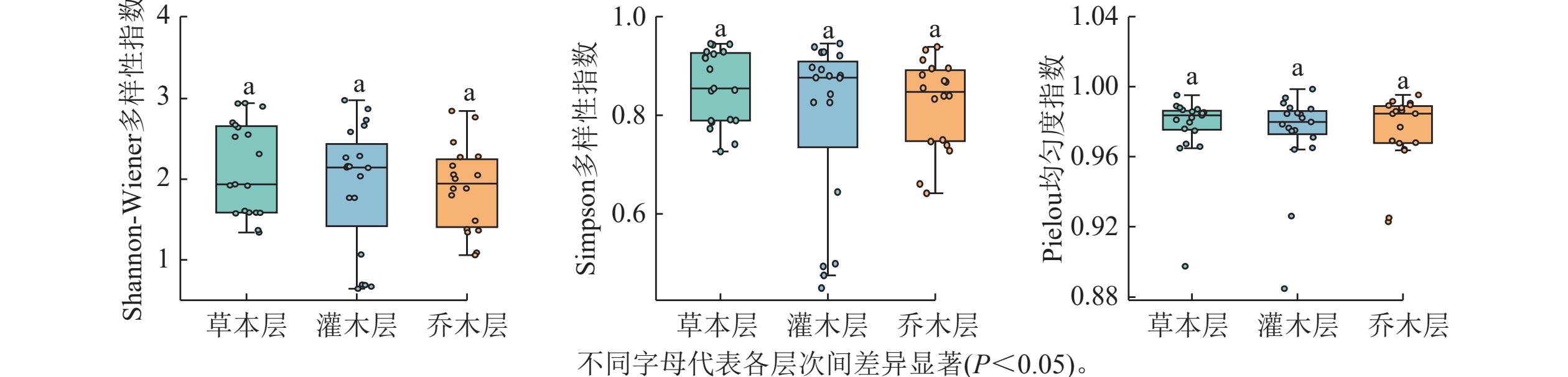
在19个样方中,不同层次的物种α多样性指数如图1所示。Shannon-Wiener多样性指数总体表现从高到低大致依次为草本层、灌木层、乔木层,但差异不显著,表现出群落演替前期特征。其中乔木层平均值为1.90,变化范围为0~2.85;灌木层平均值为1.91,变化范围为0.64~2.98;草本层平均值为2.14,变化范围为1.34~2.94。Simpson多样性指数间差异也不显著,乔木层平均值为0.82,变化范围为0.64~1.00;灌木层平均值为0.79,变化范围为0.45~0.95;草本层平均值为0.86,变化范围为0.73~0.94。Pielou均匀度指数变化小,乔木层平均值为0.97,变化范围为0~1.00;灌木层平均值为0.97,变化范围为0.88~1.00;草本层为0.98,变化范围为0.90~1.00。当考虑每个样方中所有物种时,Shannon-Wiener指数为2.25~3.81;Simpson多样性指数为0.88~0.97;Pielou均匀度指数为0.88~0.99。
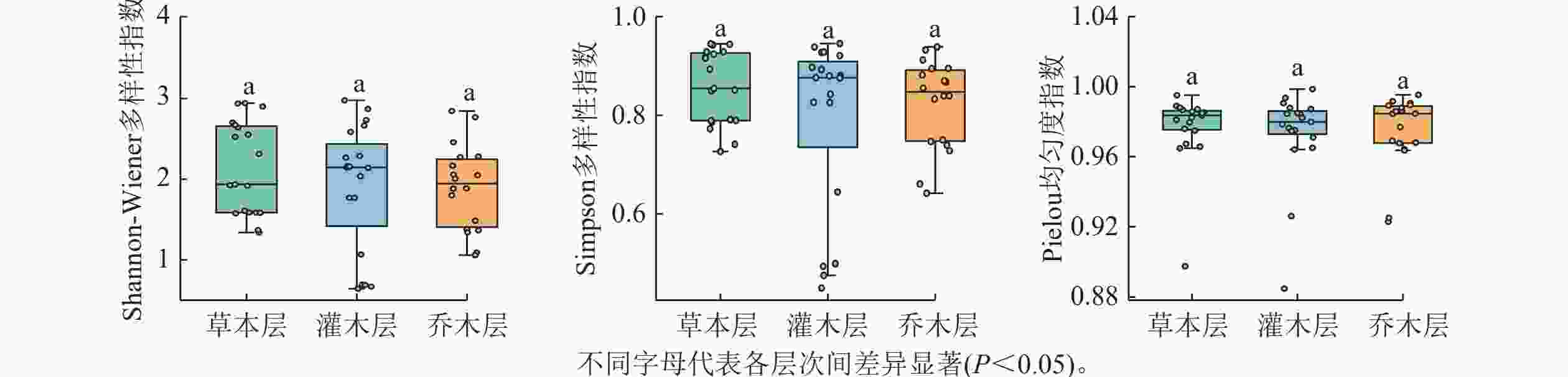
图 1 调查样方不同层次各物种多样性指数的比较
Figure 1. Comparison of species diversity indices across different vegetation layers in the surveyed plots
-
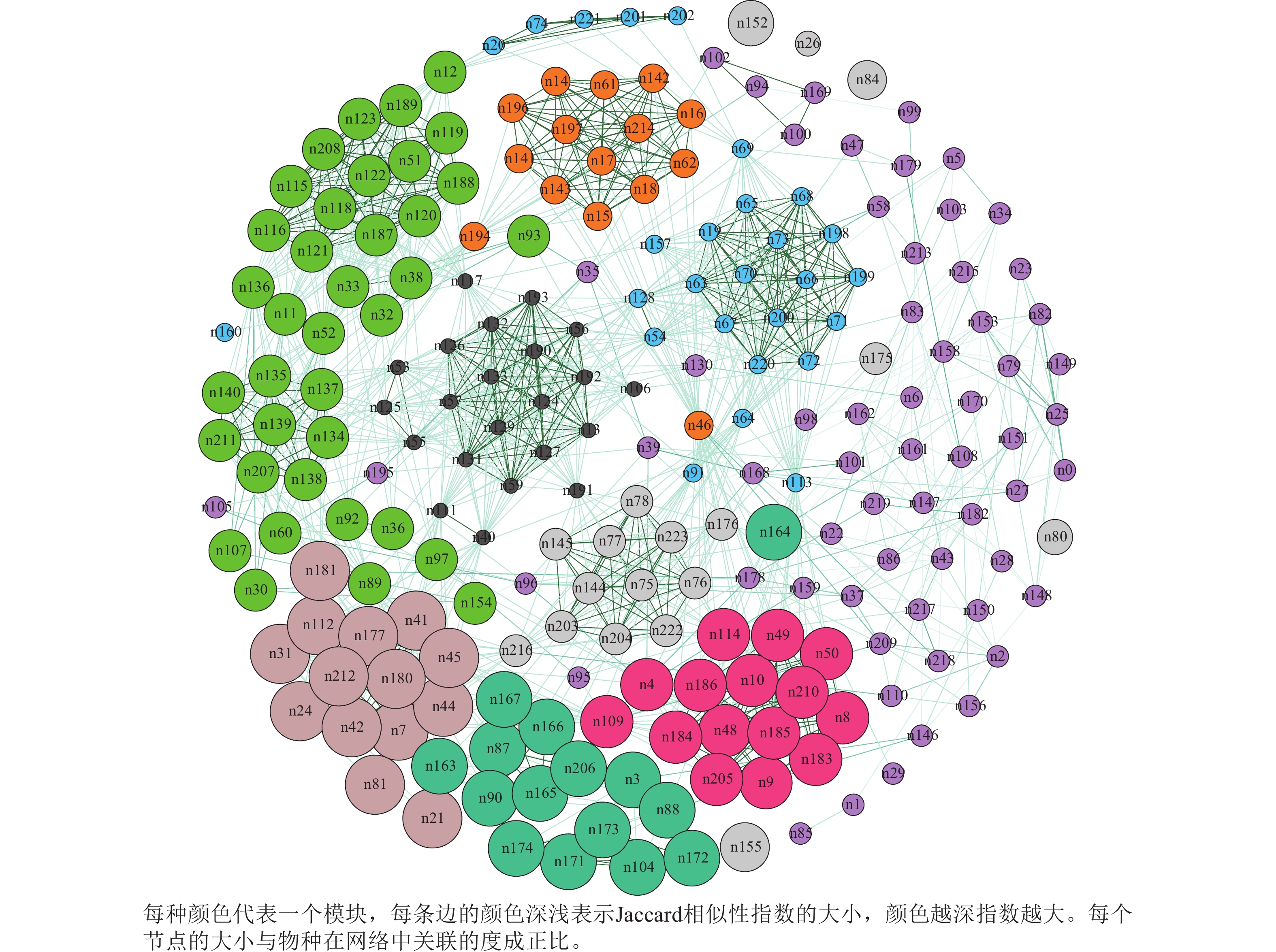
图2显示:蛛网萼群落共现网络的总节点数和边数分别为224和1 490个,平均度(节点连接性)为13.30,图密度为0.06,平均聚类系数为0.65,模块化指数为0.70,分为14个模块;蛛网萼与草本植物玉山针蔺的共现频率最高,为0.79,其次,华东瘤足蕨、麂角杜鹃、中华野海棠、杉木Cunninghamia lanceolata和树参Dendropanax dentiger的相似性指数分别为0.68、0.63、0.58、0.47和0.47,也均与蛛网萼表现出较强的共现关系。
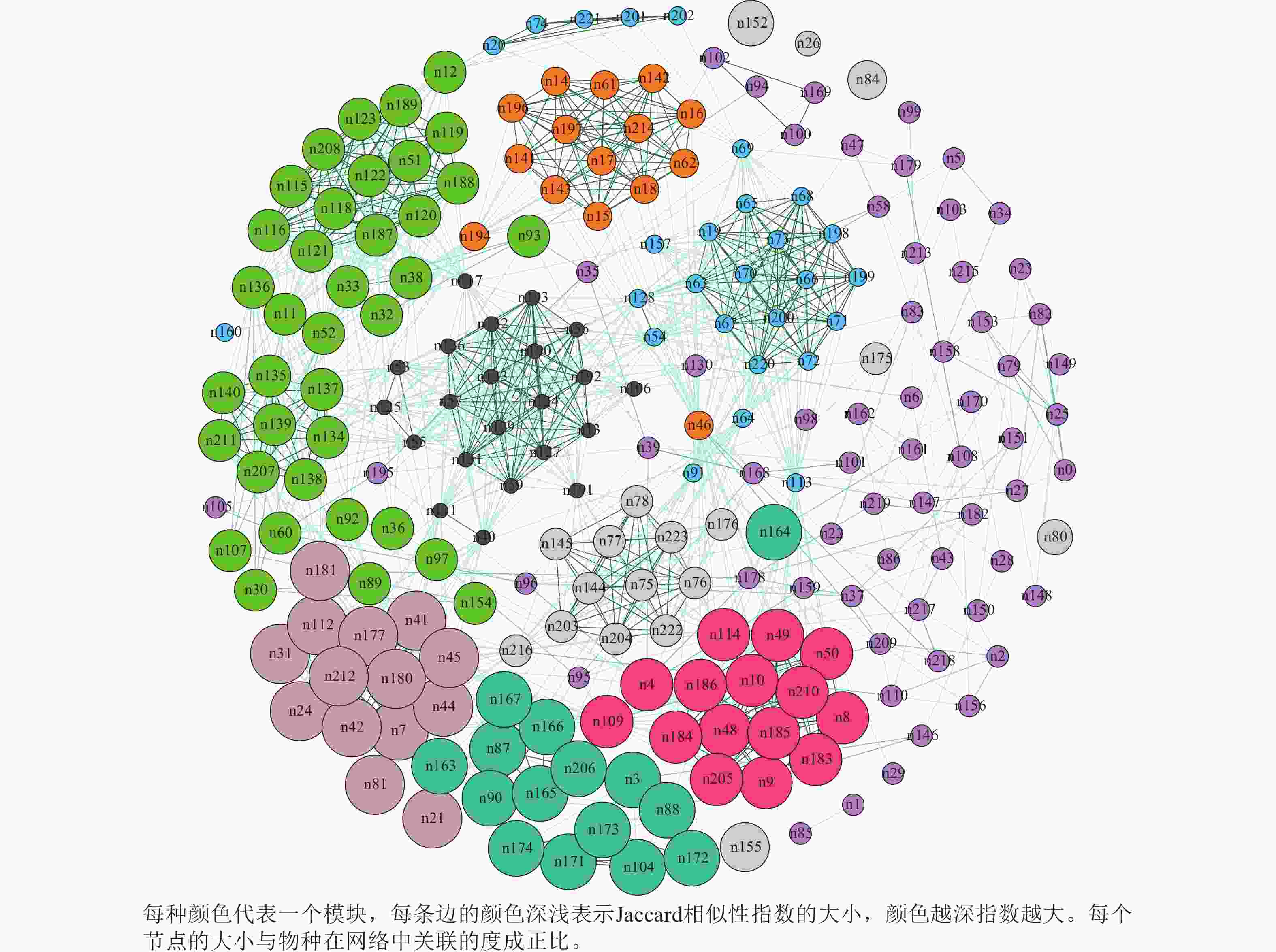
图 2 蛛网萼种群所处群落共现网络图
Figure 2. Co-occurrence network of the community inhabited by P. arguta
-
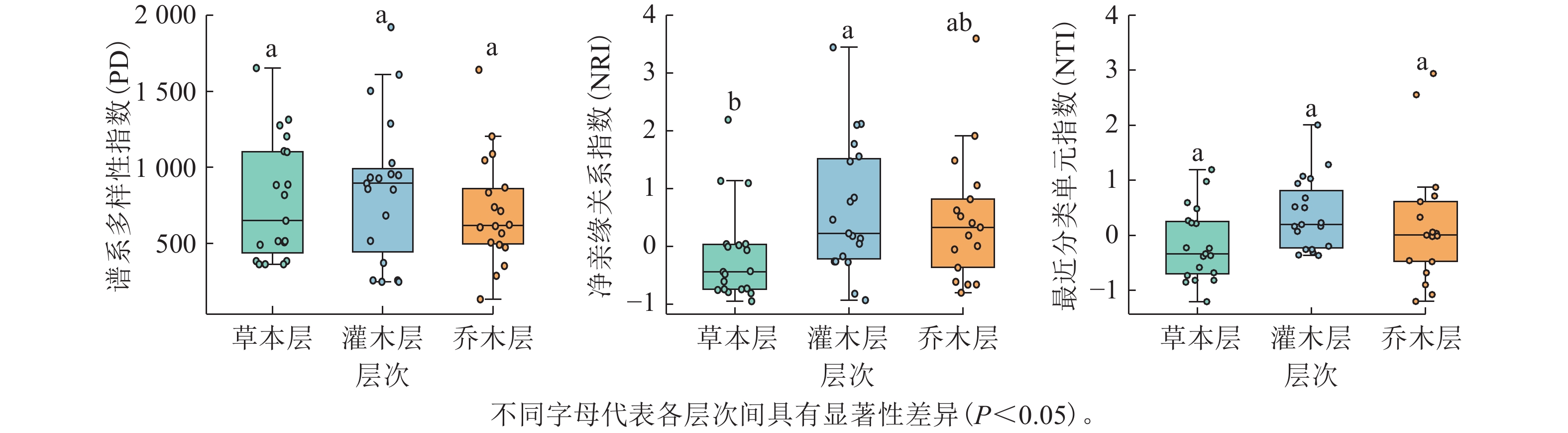
去除蕨类和裸子植物后,19个样方不同层次被子植物谱系多样性分析结果显示(图3):谱系多样性指数(PD)在乔木层(0~1 641.18,均值为674.46)、灌木层(250.48~1 920.73,均值为859.32)、草本层(365.49 ~ 1 652.81,均值为779.31)间无显著差异;当考虑所有被子植物时,PD为1 022.71~4 167.25(均值为2 287.60)。净亲缘关系指数(NRI)从高到低依次为灌木层(−0.93~3.44,均值为0.65)、乔木层(−0.80 ~ 3.59,均值为0.46)、草本层(−0.95~2.19,均值为−0.16),且灌/乔木层与草本层差异显著(P<0.05);当考虑所有被子植物时,NRI均值为0.29(−1.21 ~ 2.92)。最近分类单元指数(NTI)各层间亦无显著差异,乔木层(−1.20~2.94,均值为0.20)、灌木层(−0.36~2.01,均值为0.38)、草本层(−1.20~1.19,均值为−0.17);当考虑所有被子植物时,NTI均值为0.08(−2.19~2.55)。
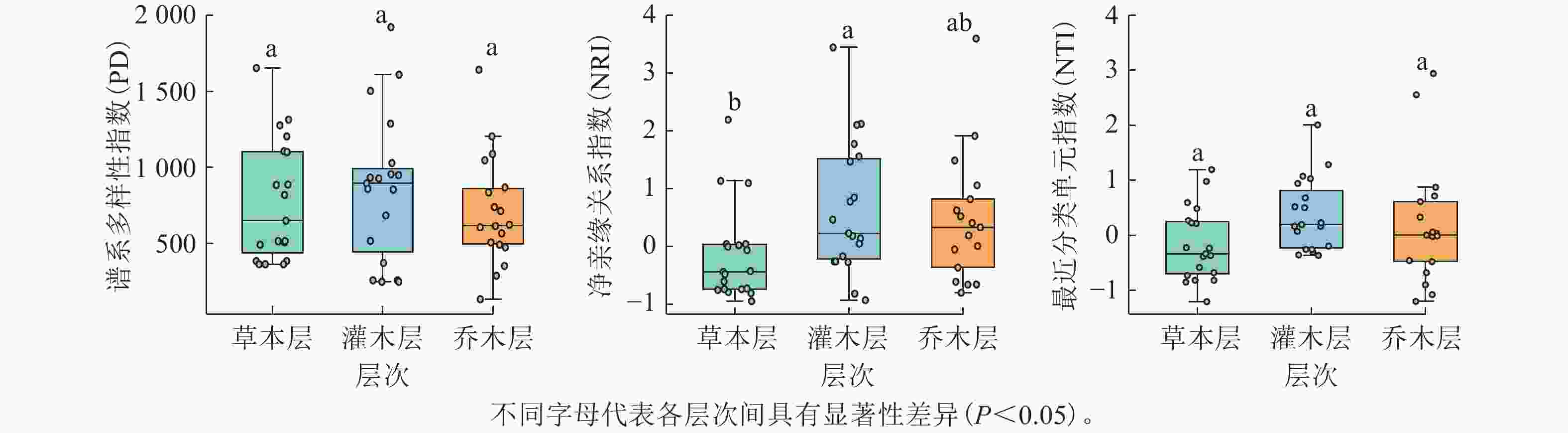
图 3 调查样方不同层次各谱系多样性指数的比较
Figure 3. Comparison of phylogenetic diversity indices across different vegetation layers in the surveyed plots
-
从群落植物功能多样性来看,19个蛛网萼样方中,功能丰富度指数(Fric)的范围为0.02~319.36,平均值为55.84,表现出较大的波动。功能均匀度指数(Feve)的范围为0.72~0.88,平均值为0.79;功能分异指数(Fdiv)的范围为0.73~0.86,平均值为0.81;Rao二次熵指数(RaoQ)的范围为7.63~16.10,平均值为12.32。
-
选取的功能性状系统发育信号检验结果如表2所示。样方中9个功能性状的K均小于1,表明样方植被群落功能性状未表现出较强的系统发育保守性。其中果实类型、种子传播类型、比叶面积、开花期和结果期均表现出比随机进化模型更弱的系统发育信号(K<1,P<0.05),表明系统发育关系对果实类型、种子传播类型、比叶面积、开花期和结果期种间变异的影响相较于其他4个功能性状更大。
表 2 蛛网萼所处群落功能性状的系统发育信号
Table 2. Phylogenetic signals of functional traits in the P. arguta community
功能性状 K P 功能性状 K P 果实类型 0.73 0.001 授粉方式 0.13 0.110 种子传播类型 0.31 0.001 植物高度 0.13 0.184 比叶面积 0.23 0.001 种子质量 0.16 0.442 开花期 0.17 0.02 生活型 0.07 0.88 结果期 0.15 0.029 -
如图4所示,蛛网萼所处群落的物种丰富度SR与Shannon-Wiener、Simpson和PD均呈极显著正相关(P<0.001)。Shannon-Wiener与Simpson间也为极显著正相关(P<0.001),且都与PD极显著正相关(P<0.001)。NRI与NTI呈显著正相关(P<0.05),除与FDiv呈显著正相关外(P<0.05),NRI和NTI与其他功能多样性指数无显著相关性,与物种多样性指数也无显著相关性。SR、物种多样性指数(Shannon-Wiener和Simpson)和PD均与FRic呈显著正相关(P<0.05),与FEve呈显著负相关(P<0.05)。RaoQ与Simpson指数和FRic均为显著正相关(P<0.05)。
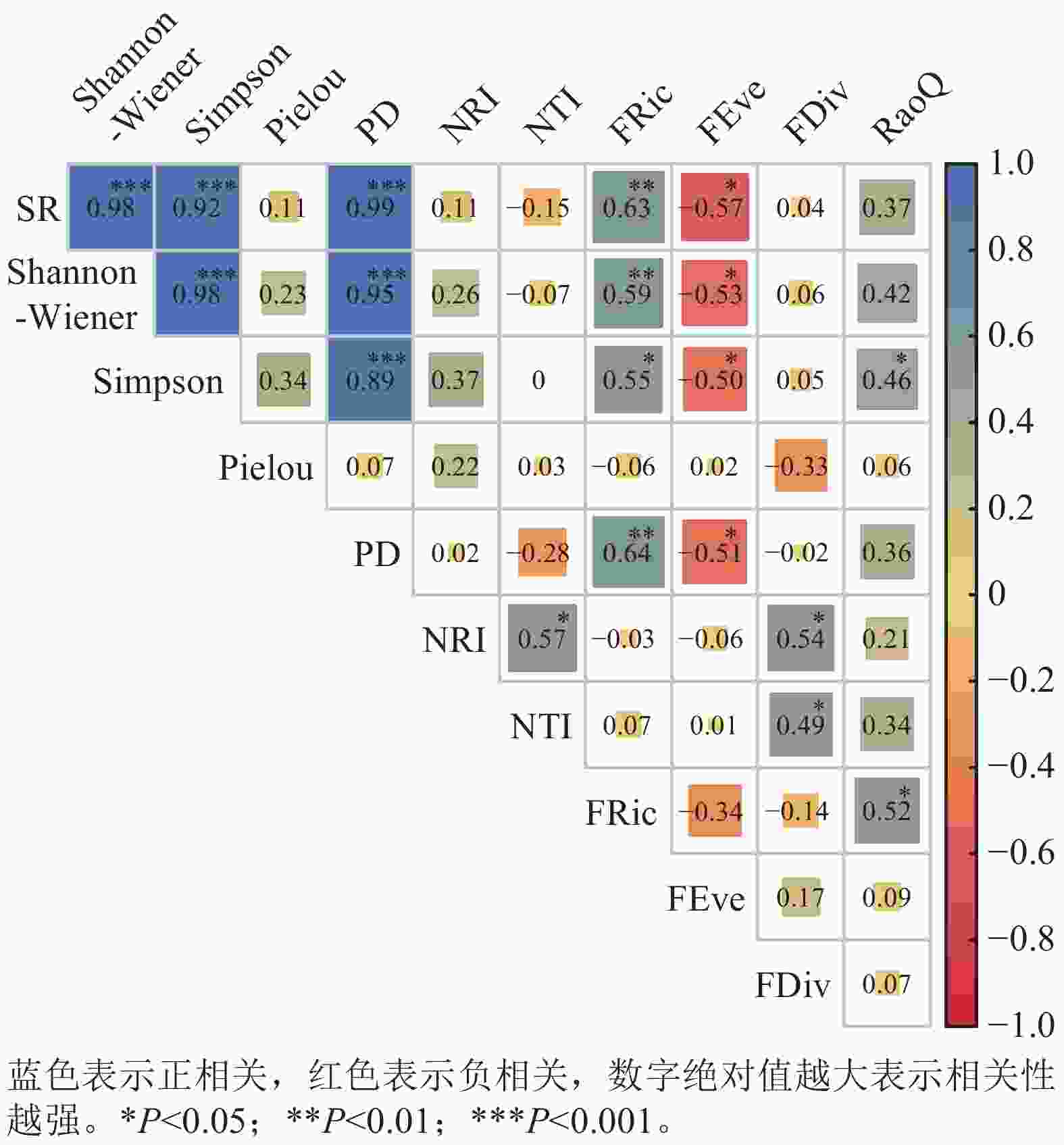
图 4 蛛网萼所处群落物种多样性、谱系多样性和功能多样性的相关性分析
Figure 4. Correlation analysis of species, phylogenetic, and functional diversity in the P. arguta community
-
百山祖国家公园作为中亚热带常绿阔叶林关键分布区,生物多样性丰富[40]。对19个有蛛网萼的样方调查显示,群落含维管植物89科162属224种,蛛网萼仅在8个样方中为所在层次的优势种,在其余样方为重要伴生种,表明蛛网萼在不同群落中的生态地位存在明显差异。群落乔木层以常绿阔叶树种为主,其重要值普遍高于落叶树种,优势种包括小叶青冈、褐叶青冈和福建柏等。灌木层以蛛网萼为主要优势种,其他优势种包括中华野海棠、宜昌荚蒾等。草本层优势种多为蕨类和喜湿植物(如华东瘤足蕨、玉山针蔺等),与蛛网萼的生境需求一致[13]。区系分析表明,群落植物属以北温带分布(18.62%)和东亚分布(17.24%)为主,温带成分占比高于热带,呈温带特征,与百山祖25 hm2样地监测结果吻合[41]。群落α多样性分析显示Shannon-Wiener指数(2.25~3.81)接近成熟群落标准(4~5),而Pielou均匀度指数(0.88~0.99)显著高于典型值(0.7~0.8)[42]。各垂直层次间多样性差异不显著,表明群落物种分布均匀。这与同为濒危植物的蝴蝶树群落的乔/灌木层多样性突出、草本层偏低形成对比[43],暗示蛛网萼群落处于演替过渡阶段,多样性结构可能受干扰历史、保护强度与生境异质性共同调控。
-
本研究基于Jaccard相似性指数构建了蛛网萼群落的物种共现网络,分析显示群落内部结构复杂,具有明显的模块化特征(模块化指数0.70,14个模块)。虽然基于存在/缺失数据的共现网络不能直接等同于种间相互作用[44],但能有效揭示潜在的生态关联,如生境过滤、互惠或竞争[45]。与蛛网萼共现频率高的物种,反映了相似的生态偏好或生境特征。玉山针蔺与蛛网萼的相似性指数最高,表明其空间分布高度一致且稳定,华东瘤足蕨、麂角杜鹃和中华野海棠等物种与蛛网萼的相似性也较高。华东瘤足蕨和中华野海棠等同样偏好潮湿生境,表明蛛网萼倾向于与喜湿润、阴生环境的物种共同分布,其共现模式可能主要由相似的生态位需求及生境过滤共同驱动。
-
谱系与功能多样性共同揭示森林群落构建机制[46]。去除蕨类与裸子植物影响后,当考虑所有被子植物时,各样方谱系多样性指数(PD)在1 022.71–4 167.25之间变化(均值为2 287.60),表明不同样方间的被子植物进化谱系组成存在较大的空间异质性。蛛网萼群落仅乔木层、灌木层与草本层的净亲缘关系指数存在显著差异。乔木层与灌木层的净亲缘关系指数和最近分类单元指数均值均大于0,表现为环境过滤驱动谱系聚集;草本层二者均值均小于0,反映竞争排斥导致谱系发散。群落整体谱系结构呈微弱聚集(均值大于0),与中高海拔群落谱系结构多呈聚集模式的规律一致[47]。功能丰富度均值为55.84,样方间变异较大,可能与物种数、功能性状异质性或极端性状物种的存在相关。功能均匀度均值为0.79,功能离散度均值为0.81,表明物种资源利用均匀且种间竞争较强[48]。Rao二次熵均值为12.32(7.63 ~ 16.10),反映物种间存在中等强度竞争与生态位重叠[49]。蛛网萼群落9个功能性状的K值均小于1,其中种子传播类型、果实类型、比叶面积、开花期和结果期均表现出比随机进化模型更弱的系统发育信号(K<1,P<0.05),表明功能性状主要受环境异质性驱动,与进化历史关联较弱[50]。功能性状与系统发育的非对应性可能源于趋同演化或分类尺度效应[51–52]。谱系多样性指数(PD)与物种丰富度多项参数呈极显著正相关,与Pielou均匀度无相关性(图4),此与多数研究结论一致[51,53],但存在区域差异(如好望角森林因进化树失衡导致错位)[54]。功能多样性(FRic)与SR、Shannon和Simpson指数均呈显著正相关,支持“物种多样性增加驱动功能丰富度提升”的理论[55–56]。谱系结构(NRI与NTI)与功能多样性整体相关性较弱,仅与功能离散度(FDiv)显著正相关,印证了谱系距离不一定反映功能距离的规律[57]。蛛网萼群落功能性状的分布与系统发育关系并不完全一致,且功能多样性与物种多样性之间显著相关,说明群落功能结构的形成受谱系约束有限,环境条件可能在塑造功能差异格局中发挥作用,同时不同多样性维度对群落结构的响应具有一定差异。
-
本研究基于物种、谱系和功能多样性,系统分析了百山祖国家公园蛛网萼天然群落的结构特征。结果表明,蛛网萼所处群落维管植物种类丰富,以温带区系成分为主,蛛网萼仅在部分样方中占据优势地位,显示其在不同群落中的生态地位存在差异;群落物种分布均匀,处于演替过渡阶段,受环境干扰较强;群落整体以及乔木层和灌木层的谱系结构均呈谱系聚集格局,由生境过滤驱动,而草本层表现为谱系发散,反映竞争排斥;功能丰富度与物种丰富度显著正相关,而多数功能性状的系统发育信号较弱,表明群落功能结构的形成受系统发育约束有限,环境异质性可能在塑造功能差异格局中发挥主要作用。
百山祖国家公园蛛网萼的保护应聚焦于所处群落的整体结构稳定性及多样性维持。除了蛛网萼,还应重视群落中的优势种及稳定共现物种。通过长期监测其物种组成变化,评估群落结构稳定性。同时,考虑到蛛网萼群落在不同多样性维度的响应存在差异,保护与监测中不宜仅依赖单一多样性指标。
Community structure and diversity of the endangered plant Platycrater arguta in Baishanzu National Park
-
摘要:
目的 探索濒危植物蛛网萼Platycrater arguta的天然群落结构、物种多样性特征,为保护区制定科学保护策略提供理论依据。 方法 利用样方调查法,于浙江钱江源-百山祖国家公园候选区百山祖园区(百山祖国家公园)设立19个代表性样方开展群落学调查。 结果 ①19个样方中共记录到224种维管植物(含变种和变型),隶属于89科162属,其中蛛网萼在8个样方中重要值居前,呈优势状态;区系组成以温带成分为主,具显著东亚特征。②群落各层次物种分布均匀,处于演替过渡期,受环境干扰较强。③蛛网萼与草本植物玉山针蔺Trichophorum subcapitatum的生态共现关系最为显著,显示出相似生境偏好。④群体整体以及乔木层与灌木层的谱系结构呈谱系聚集,草本层呈谱系发散。⑤群落功能丰富度变异显著,多数功能性状的系统发育信号较弱。⑥物种α多样性与谱系多样性和功能丰富度均呈显著正相关,但谱系结构与功能多样性的相关性较弱。 结论 百山祖国家公园蛛网萼天然群落维管植物种类丰富,以温带区系为主;群落处于演替过渡阶段,整体谱系结构呈弱聚集格局,不同层次间存在一定差异;功能性状主要受环境异质性驱动,系统发育约束作用有限。蛛网萼倾向于与喜湿阴生物种共存,应协同保护其关键共现物种及生境,维持群落结构稳定性。此外,群落监测与评估应综合多维度多样性指标,避免仅依赖单一指标。图4表2参57 Abstract:Objective To explore the natural community structure and species diversity characteristics of Platycrater arguta (Hydrangeaceae), an endangered shrub endemic to East Asia, providing a basis for formulating scientific conservation strategies within protected areas. Method Community surveys were conducted using the quadrat method. Nineteen representative quadrats were established within the candidate area of Qianjiangyuan-Baishanzu National Park Baishanzu area in Zhejiang Province (Baishanzu National Park). Result (1) A total of 224 vascular plant species (including varieties/forms), belonging to 162 genera and 89 families, were recorded across 19 quadrats. P. arguta exhibited high importance values and was dominant in 8 quadrats. The floristic composition was dominated by temperate elements, exhibiting distinct East Asian characteristics. (2) Species were evenly distributed across all community layers, indicating a transitional successional stage subject to strong environmental disturbance. (3) P. arguta showed the most significant ecological co-occurrence relationship with the herb Trichophorum subcapitatum. Simpson, indicating similar habitat preferences. (4) Phylogenetic structure analysis revealed that the entire community, the tree and shrub layers were phylogenetically clustered, while the herb layer was phylogenetically overdispersed. (5) Community functional richness varied significantly, and phylogenetic signals for major functional traits were generally weak. (6) Species α-diversity was significantly positively correlated with both phylogenetic diversity and functional richness, but the correlation between phylogenetic structure and functional diversity was weak. Conclusion Baishanzu National Park harbours P. arguta natural communities with high vascular plant diversity of temperate affinity. Currently in transitional successional stages, these communities show weak phylogenetic clustering that varies among strata. Environmental heterogeneity rather than phylogenetic history shapes functional traits. P. arguta preferentially associates with shade-tolerant, hygrophilous species, necessitating coordinated conservation of these key associates and their habitats. Multi-dimensional diversity metrics should be adopted for monitoring instead of single indicators. [Ch, 4 fig. 2 tab. 57 ref.] -
图 1 调查样方不同层次各物种多样性指数的比较
Figure 1 Comparison of species diversity indices across different vegetation layers in the surveyed plots
图 2 蛛网萼种群所处群落共现网络图
Figure 2 Co-occurrence network of the community inhabited by P. arguta
图 3 调查样方不同层次各谱系多样性指数的比较
Figure 3 Comparison of phylogenetic diversity indices across different vegetation layers in the surveyed plots
图 4 蛛网萼所处群落物种多样性、谱系多样性和功能多样性的相关性分析
Figure 4 Correlation analysis of species, phylogenetic, and functional diversity in the P. arguta community
表 1 蛛网萼所处的天然群落样方调查信息
Table 1. General survey information of plots in the natural communities of P. arguta
样方名称 简称 植被类型 样方面积/(m×m) 经度N/(°) 纬度W/(°) 海拔/m 地形 百瀑沟Z01 BPGZ01 针阔混交林 10×10 119.193022 27.751369 1 420 石壁 百瀑沟Z02 BPGZ02 落阔混交林 20×20 119.192653 27.751344 1 401 石壁 百瀑沟Z03 BPGZ03 落阔混交林 20×20 119.191944 27.751228 1 384 石壁 百瀑沟Z04 BPGZ04 常绿阔叶林 20×20 119.189444 27.749722 1 339 石壁 百瀑沟Z05 BPGZ05 常绿阔叶林 20×20 119.189722 27.748417 1 307 石壁 三井溪Z01 SJXZ01 常绿阔叶林 20×20 119.207111 27.795397 1 195 石壁 三井溪Z02 SJXZ02 针阔混交林 20×20 119.207744 27.795544 1 196 石壁 三井溪Z10 SJXZ10 针阔混交林 20×20 119.209306 27.796828 1 194 石壁 三井溪Z12 SJXZ12 针阔混交林 20×20 119.210203 27.800578 1 169 石壁 盘龙地桥边01 PLDQ01 常绿阔叶林 20×20 119.22875 27.764806 1 093 山地 盘龙地桥边02 PLDQ02 针阔混交林 20×20 119.236906 27.765797 1 146 山涧 百管站公路边01 GL01 常绿阔叶林 20×20 119.192439 27.750347 1 416 石壁 半坑01 BK01 常绿阔叶林 20×20 119.150131 27.717856 744 山地 溧阳村01 LYC01 针阔混交林 20×20 119.182414 27.820133 1 042 山地 溧阳村03 LYC03 针阔混交林 20×20 119.203747 27.822078 1 070 山地 久住垟公路边01 JZY01 针阔混交林 20×20 119.248611 27.732456 1 323 山地 哨卡01 SK01 落叶阔叶林 20×20 119.194519 27.909483 1 228 山地 猕猴谷-桑瀑飞雪01 MHG01 常绿阔叶林 20×20 119.194444 27.916667 940 溪沟 炉岙01 LA01 毛竹落叶林 20×20 119.206719 27.916352 1 123 山地  下载: 导出CSV
下载: 导出CSV
表 2 蛛网萼所处群落功能性状的系统发育信号
Table 2. Phylogenetic signals of functional traits in the P. arguta community
功能性状 K P 功能性状 K P 果实类型 0.73 0.001 授粉方式 0.13 0.110 种子传播类型 0.31 0.001 植物高度 0.13 0.184 比叶面积 0.23 0.001 种子质量 0.16 0.442 开花期 0.17 0.02 生活型 0.07 0.88 结果期 0.15 0.029
下载: 导出CSV
-
[1] POMMERENING A, SVENSSON A, ZHAO Zhonghua, et al. Spatial species diversity in temperate species-rich forest ecosystems: revisiting and extending the concept of spatial species mingling[J]. Ecological Indicators, 2019, 105: 116−125. DOI: 10.1016/j.ecolind.2019.05.060. [2] 杨阳, 马立辉, 王海洋. 濒危植物树枫杜鹃(Rhododendron changii)种群结构及伴生群落特征[J]. 生态学杂志, 2019, 38(5): 1352−1362. YANG Yang, MA Lihui, WANG Haiyang. Population structure and companion community characteristics of the endangered species, Rhododendron changii[J]. Chinese Journal of Ecology, 2019, 38(5): 1352−1362. DOI: 10.13292/j.1000-4890.201905.013. [3] CADOTTE M W, CARSCADDEN K, MIROTCHNICK N. Beyond species: functional diversity and the maintenance of ecological processes and services[J]. Journal of Applied Ecology, 2011, 48(5): 1079−1087. DOI: 10.1111/j.1365-2664.2011.02048.x. [4] WEBB C O, ACKERLY D D, MCPEEK M A, et al. Phylogenies and community ecology[J]. Annual Review of Ecology and Systematics, 2002, 33: 475−505. DOI: 10.1146/annurev.ecolsys.33.010802.150448. [5] CADOTTE M W, DINNAGE R, TILMAN D. Phylogenetic diversity promotes ecosystem stability[J]. Ecology, 2012, 93(sppl 8): S223–S233. DOI: 10.1890/11-0426.1. [6] THORNHILL A H, MISHLER B D, KNERR N J, et al. Continental-scale spatial phylogenetics of Australian angiosperms provides insights into ecology, evolution and conservation[J]. Journal of Biogeography, 2016, 43(11): 2085−2098. DOI: 10.1111/jbi.12797. [7] ACKERLY D D, SCHWILK D W, WEBB C O. Niche evolution and adaptive radiation: testing the order of trait divergence[J]. Ecology, 2006, 87(sppl7): S50−S61. DOI: 10.1890/0012-9658. [8] 牛克昌, 刘怿宁, 沈泽昊, 等. 群落构建的中性理论和生态位理论[J]. 生物多样性, 2009, 17(6): 579−593. NIU Kechang, LIU Yining, SHEN Zehao, et al. Community assembly: the relative importance of neutral theory and niche theory[J]. Biodiversity Science, 2009, 17(6): 579−593. [9] 刘晓娟, 马克平. 植物功能性状研究进展[J]. 中国科学(生命科学), 2015, 45(4): 325−339. LIU Xiaojuan, MA Keping. Plant functional traits−concepts, applications and future directions[J]. Science in China (Series C), 2015, 45(4): 325−339. DOI: 10.1360/N052014-00244. [10] LI Ying, LIU Congcong, XU Li, et al. Leaf trait networks based on global data: representing variation and adaptation in plants[J]. Frontiers in Plant Science, 2021, 12: 710530. DOI: 10.3389/fpls.2021.710530. [11] 张鑫, 刘旻霞, 陈雪娇, 等. 甘南亚高寒草甸不同坡位植物群落物种共存机制[J]. 生态学报, 2025, 45(6): 2582−2594. ZHANG Xin, LIU Minxia, CHEN Xuejiao, et al. Species coexistence mechanism of plant communities on different slopes of Gannan sub-Alpine meadows[J]. Acta Ecologica Sinica, 2025, 45(6): 2582−2594. DOI: 10.20103/j.stxb.202404220898. [12] 曹丽雯, 支肖, 戴嘉禾, 等. 蛛网萼未成熟种子培养与离体繁殖[J]. 浙江大学学报(农业与生命科学版), 2019, 45(2): 157−163. CAO Liwen, ZHI Xiao, DAI Jiahe, et al. Immature seed culture and in vitro propagation of Platycrater arguta[J]. Journal of Zhejiang University (Agriculture and Life Sciences), 2019, 45(2): 157−163. DOI: 10.3785/j.issn.1008-9209.2018.03.261. [13] 宫莉霞, 林韧安, 陈旭华, 等. 珍稀濒危植物蛛网萼生境调查及其保护建议[J]. 农业科技通讯, 2022(5): 230−232. GONG Lixia, LIN Ren’an, CHEN Xuhua, et al. Investigation on the habitat of cobweb Calyx, a rare and endangered plant, and its protection suggestions[J]. Bulletin of Agricultural Science and Technology, 2022(5): 230−232. [14] QI Xinshuai, YUAN Na, COMES H P, et al. A strong ‘filter’ effect of the East China Sea land bridge for East Asia’s temperate plant species: inferences from molecular phylogeography and ecological niche modelling of Platycrater arguta(Hydrangeaceae)[J]. BMC Evolutionary Biology, 2014, 14: 41. DOI: 10.1186/1471-2148-14-41. [15] WEI Chunyan, LUO Guangyu, JIN Zexin, et al. Physiological and structural changes in leaves of Platycrater arguta seedlings exposed to increasing light intensities[J]. Plants, 2024, 13(9): 1263. DOI: 10.3390/plants13091263. [16] ZHANG Xule, MA Xiaohua, HU Yaping, et al. Effects of exogenous spraying of melatonin on the growth of Platycrater arguta under drought stress[J]. Frontiers in Plant Science, 2025, 15: 1516302. DOI: 10.3389/fpls.2024.1516302. [17] 张丽芳, 裘利洪. 蛛网萼开花物候、花部特征及繁育系统研究[J]. 广西植物, 2017, 37(10): 1301−1311. ZHANG Lifang, QIU Lihong. Flowering phenology, floral traits and breeding system of Platycrater arguta[J]. Guihaia, 2017, 37(10): 1301−1311. DOI: 10.11931/guihaia.gxzw201612006. [18] ZHANG Xule, FENG Lei, MA Xiaohua, et al. Characterization and analysis of the complete mitochondrial genome of Platycrater arguta[J]. Current Issues in Molecular Biology, 2025, 47(7): 521. DOI: 10.3390/cimb47070521. [19] 毛夷仙, 袁荣斌, 程英, 等. 江西武夷山国家级自然保护区蛛网萼群落的发现与保护建议[J]. 南方林业科学, 2016, 44(2): 67−68,72. MAO Yixian, YUAN Rongbin, CHENG Ying, et al. The discovery and conservation recommendations of Platycrater arguta community in Jiangxi Wuyishan National Nature Reserve[J]. South China Forestry Science, 2016, 44(2): 67−68,72. DOI: 10.16259/j.cnki.36-1342/s.2016.02.015. [20] 姚嘉, 张聪伶, 李时轩, 等. 百山祖连续海拔样带植物群落特征[J]. 生物多样性, 2024, 32(12): 45−59. YAO Jia, ZHANG Congling, LI Shixuan, et al. Characteristics of plant communities in the Baishanzu continuous elevational transect[J]. Biodiversity Science, 2024, 32(12): 45−59. DOI: 10.17520/biods.2024052. [21] 龙丹, 吴逸卿, 周伟龙, 等. 百山祖国家公园与邻近地区常绿阔叶林群落特征比较[J]. 浙江农林大学学报, 2025, 42(1): 12−22. LONG Dan, WU Yiqing, ZHOU Weilong, et al. Differences in community characteristics of evergreen broad-leaved forests between Baishanzu National Park and adjacent areas[J]. Journal of Zhejiang A&F University, 2025, 42(1): 12−22. DOI: 10.11833/j.issn.2095-0756.20240456. [22] 金孝锋, 丁炳扬, 郑朝宗, 等. 浙江百山祖自然保护区种子植物区系分析[J]. 云南植物研究, 2004, 26(6): 605−618. JIN Xiaofeng, DING Bingyang, ZHENG Chaozong, et al. The floristic analysis of seed plants in Baishanzu nature reserve from Zhejiang Province[J]. Acta Botanica Yunnanica, 2004, 26(6): 605−618. DOI: 10.3969/j.issn.2095-0845.2004.06.002. [23] 方精云, 王襄平, 沈泽昊, 等. 植物群落清查的主要内容、方法和技术规范[J]. 生物多样性, 2009, 17(6): 533−548. FANG Jingyun, WANG Xiangping, SHEN Zehao, et al. Methods and protocols for plant community inventory[J]. Biodiversity Science, 2009, 17(6): 533−548. DOI: 10.3724/SP.J.1003.2009.09253. [24] BUTLER B J, BARCLAY J S, FISHER J P. Plant communities and flora of Robins Island (Long Island), New York[J]. Journal of the Torrey Botanical Society, 1999, 126(1): 63. DOI: 10.2307/2997256. [25] 《浙江植物志(新编)》编辑委员会. 浙江植物志(新编)[M]. 杭州: 浙江科学技术出版社, 2021. Editorial Board of Flora of Zhejiang (New Edition). Flora of Zhejiang (New Edition) [M]. Hanghzou: Zhejiang Science and Technology Publishing House, 2021. [26] 金孝锋, 鲁益飞, 丁炳扬, 等. 浙江种子植物物种编目[J]. 生物多样性, 2022, 30(6): 27−35. JIN Xiaofeng, LU Yifei, DING Bingyang, et al. Species cataloging of the seed plants in Zhejiang, East China[J]. Biodiversity Science, 2022, 30(6): 27−35. DOI: 10.17520/biods.2021408. [27] 吴征镒. 中国种子植物属的分布区类型[J]. 云南植物研究, 1991, 13(增刊Ⅳ): 1–139. WU Zhengyi. The areal-types of Chinese genera of seed plants [J]. Acta Botanica Yunnanica, 1991, 13(Suppl Ⅳ): 1–139. [28] 邓文婕, 吴华征, 李添翔, 等. 洞头国家级海洋公园主要植被类型及其特征[J]. 植物生态学报, 2024, 48(2): 254−268. DENG Wenjie, WU Huazheng, LI Tianxiang, et al. Main vegetation types and characteristics in Dongtou National Marine Park, Zhejiang, China[J]. Chinese Journal of Plant Ecology, 2024, 48(2): 254−268. [29] HURLBERT S H. The nonconcept of species diversity: a critique and alternative parameters[J]. Ecology, 1971, 52(4): 577−586. DOI: 10.2307/1934145. [30] PEET R K. The measurement of species diversity[J]. Annual Review of Ecology and Systematics, 1974, 5: 285−307. DOI: 10.1146/annurev.es.05.110174.001441. [31] 米畅, 彭羽. 植物物种网络特征及其应用研究进展[J]. 应用与环境生物学报, 2025, 31(11): 1821−1833. MI Chang, PENG Yu. Advances in plant species network traits and their applications[J]. Chinese Journal of Applied and Environmental Biology, 2025, 31(11): 1821−1833. DOI: 10.19675/j.cnki.1006-687x.2024.09028. [32] JIN Yi, QIAN Hong. V. PhyloMaker2: an updated and enlarged R package that can generate very large phylogenies for vascular plants[J]. Plant Diversity, 2022, 44(4): 335−339. DOI: 10.1016/j.pld.2022.05.005. [33] WEBB C O. Exploring the phylogenetic structure of ecological communities: an example for rain forest trees[J]. The American Naturalist, 2000, 156(2): 145−155. DOI: 10.1086/303378. [34] WANG Han, HARRISON S P, LI Meng, et al. The China plant trait database version 2[J]. Scientific Data, 2022, 9: 769. DOI: 10.1038/s41597-022-01884-4. [35] KATTGE J, BÖNISCH G, DÍAZ S, et al. TRY plant trait database–enhanced coverage and open access[J]. Global Change Biology, 2020, 26(1): 119−188. DOI: 10.1111/gcb.14904. [36] LE Haichuan, MAO Jiangtao, CAVENDER-BARES J, et al. Non-native plants tend to be phylogenetically distant but functionally similar to native plants under intense disturbance at the Three Gorges Reservoir Area[J]. New Phytologist, 2024, 244(5): 2078−2088. DOI: 10.1111/nph.20126. [37] PETCHEY O L, GASTON K J. Functional diversity (FD), species richness and community composition[J]. Ecology Letters, 2002, 5(3): 402−411. DOI: 10.1046/j.1461-0248.2002.00339.x. [38] BLOMBERG S P, GARLAND T, IVES A R. Testing for phylogenetic signal in comparative data: behavioral traits are more labile[J]. Evolution, 2003, 57(4): 717. DOI: 10.1554/0014-3820. [39] 王诗韵, 吕光辉, 蒋腊梅, 等. 不同尺度下艾比湖典型植物群落功能多样性和系统发育多样性研究[J]. 生态环境学报, 2020, 29(5): 889−900. WANG Shiyun, LÜ Guanghui, JIANG Lamei, et al. Multi-scale analysis on functional diversity and phylogenetic diversity of typical plant community in Ebinur Lake[J]. Ecology and Environmental Sciences, 2020, 29(5): 889−900. DOI: 10.16258/j.cnki.1674-5906.2020.05.004. [40] 林阳. 百山祖国家公园海拔梯度上植物群落结构和物种多样性变化[D]. 金华: 浙江师范大学, 2024. LIN Yang. Changes in Plant Community Structure and Species Diversity along the Elevation Gradient in Baishanzu National Park[D]. Jinhua: Zhejiang Normal University, 2024. [41] 刘啸林, 吴友贵, 张敏华, 等. 浙江百山祖25 ha亚热带森林动态监测样地群落组成与结构特征[J]. 生物多样性, 2024, 32(2): 21−31. LIU Xiaolin, WU Yougui, ZHANG Minhua, et al. Community composition and structure of a 25-ha forest dynamics plot of subtropical forest in Baishanzu, Zhejiang Province[J]. Biodiversity Science, 2024, 32(2): 21−31. DOI: 10.17520/biods.2023294. [42] 王冠舜, 刘西, 张沛然, 等. 中国特有植物泰顺杜鹃所处群落特征与物种多样性分析[J]. 植物资源与环境学报, 2025, 34(1): 52−60. WANG Guanshun, LIU Xi, ZHANG Peiran, et al. Analyses on characteristics and species diversity of Chinese endemic species Rhododendron taishunense located communities[J]. Journal of Plant Resources and Environment, 2025, 34(1): 52−60. DOI: 10.3969/j.issn.1674-7895.2025.01.06. [43] SHANG Naiyan, HE Shaocui, QI Dongling, et al. Structure and stability in the community with the endangered plant Heritiera parvifolia Merr. (Malvaceae) in lowland rainforest in Diaoluo Mountain of Hainan Island, China[J]. Frontiers in Plant Science, 2025, 16: 1578361. DOI: 10.3389/fpls.2025.1578361. [44] BLANCHET F G, CAZELLES K, GRAVEL D. Co-occurrence is not evidence of ecological interactions[J]. Ecology Letters, 2020, 23(7): 1050−1063. DOI: 10.1111/ele.1352. [45] CHILLO V, VÁZQUEZ D P, TAVELLA J, et al. Plant–plant co-occurrences under a complex land-use gradient in a temperate forest[J]. Oecologia, 2021, 196(3): 815−824. DOI: 10.1007/s00442-021-04953-4. [46] CADOTTE M W, CARBONI M, SI Xingfeng, et al. Do traits and phylogeny support congruent community diversity patterns and assembly inferences?[J]. Journal of Ecology, 2019, 107(5): 2065−2077. DOI: 10.1111/1365-2745.13247. [47] 樊晓玲, 刘彬, 孙庆振, 等. 库车北山植物群落物种系统发育多样性及功能多样性沿海拔梯度的分布格局[J]. 生态学报, 2025, 45(18): 8862−8875. FAN Xiaoling, LIU Bin, SUN Qingzhen, et al. Phylogenetic and functional diversity patterns of plant communities along the altitude gradient in the Beishan Mountains of Kuqa[J]. Acta Ecologica Sinica, 2025, 45(18): 8862−8875. DOI: 10.20103/j.stxb.202409232305. [48] MASON N W H, MOUILLOT D, LEE W G, et al. Functional richness, functional evenness and functional divergence: the primary components of functional diversity[J]. Oikos, 2005, 111(1): 112−118. DOI: 10.1111/j.0030-1299.2005.13886.x. [49] SUDING K N, LAVOREL S, CHAPIN F S III, et al. Scaling environmental change through the community-level: a trait-based response-and-effect framework for plants[J]. Global Change Biology, 2008, 14(5): 1125−1140. DOI: 10.1111/j.1365-2486.2008.01557.x. [50] SWENSON N G, ENQUIST B J, THOMPSON J, et al. The influence of spatial and size scale on phylogenetic relatedness in tropical forest communities[J]. Ecology, 2007, 88(7): 1770−1780. DOI: 10.1890/06-1499.1. [51] 姜晓燕, 高圣杰, 蒋燕, 等. 毛乌素沙地植被不同恢复阶段植物群落物种多样性、功能多样性和系统发育多样性[J]. 生物多样性, 2022, 30(5): 15−25. JIANG Xiaoyan, GAO Shengjie, JIANG Yan, et al. Species diversity, functional diversity, and phylogenetic diversity in plant communities at different phases of vegetation restoration in the Mu Us sandy grassland[J]. Biodiversity Science, 2022, 30(5): 15−25. DOI: 10.17520/biods.2021387. [52] KRAFT N J B, VALENCIA R, ACKERLY D D. Functional traits and niche-based tree community assembly in an Amazonian forest[J]. Science, 2008, 322(5901): 580−582. DOI: 10.1126/science.1160662. [53] 陈博, 江蓝, 谢子扬, 等. 格氏栲天然林林窗植物物种多样性与系统发育多样性[J]. 生物多样性, 2021, 29(4): 439−448. CHEN Bo, JIANG Lan, XIE Ziyang, et al. Taxonomic and phylogenetic diversity of plants in a Castanopsis kawakamii natural forest[J]. Biodiversity Science, 2021, 29(4): 439−448. DOI: 10.17520/biods.2020399. [54] FOREST F, GRENYER R, ROUGET M, et al. Preserving the evolutionary potential of floras in biodiversity hotspots[J]. Nature, 2007, 445(7129): 757−760. DOI: 10.1038/nature05587. [55] DONG Shikui, TANG Lin, ZHANG Xiangfeng, et al. Relationship between plant species diversity and functional diversity in Alpine grasslands[J]. Acta Ecologica Sinica, 2017, 37(5): 1472−1483. DOI: 10.5846/stxb201509281981. [56] XUE Qianni, YAN Ming, BI Runcheng. Functional diversity research of tree and shrub layers in forest communities of the Wulu Mountains Nature Reserve in Shanxi, China[J]. Acta Ecologica Sinica, 2015, 35(21): 7023−7032. DOI: 10.5846/stxb201401280206. [57] JUNKER R R, BLÜTHGEN N, KELLER A. Functional and phylogenetic diversity of plant communities differently affect the structure of flower-visitor interactions and reveal convergences in floral traits[J]. Evolutionary Ecology, 2015, 29(3): 437−450. DOI: 10.1007/s10682-014-9747-2. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20250611
 点击查看大图
点击查看大图
计量
- 文章访问数: 112
- HTML全文浏览量: 50
- PDF下载量: 25
- 被引次数: 0
