





-
城市森林是一种与城市发展密切相关的森林类型[1]。城镇化的快速推进加速了植物群落的更新速度,对城市森林提供的生态服务功能提出了更高的要求。由于城市森林直接服务于当地社区和民众,景观美学方面的考量显得尤为重要[2-3],但如果只考虑眼前的景观美学特征,不重视植物群落长期稳定的更替,城市森林将永远无法满足城市可持续发展的需求。植物群落是城市森林生态系统的基础,为人们提供了诸多生态服务功能[2]。每个物种对于构建城市森林植物群落和完善生态系统的功能都至关重要,各个物种在贡献途径和功能性质方面存在差异。在物种多样性指数中,物种在构建植物群落,维持植物生态系统等方面的作用并没有得到完整的呈现。生态系统功能除了与物种的分布格局有关,还与物种具备的功能特征有更为紧密的联系[4]。与物种的多样性相比,植物的功能性状以及系统发育组成等方面的信息对生态功能系统方面出现变化的解释相对更多[5-6]。2个有相同物种多样性的群落,可能会有相似的系统发育信息,但也可能会大不相同[7]。某些群落在一定范围内还会呈现出较低的功能多样性,但是谱系多样性以及物种多样性却会很高[8-9]。为更加全面地认识植物群落的结构变化、演替过程,并预判其未来的变化趋势,需要深入分析多个层面的植物多样性。本研究通过调查海口城市主建成区范围内的植物样方数据集,分析海口城市森林植物群落的结构特征、基调物种等方面的信息,评价城市森林植物群落的整体质量,并探讨植物物种多样性、功能多样性和谱系多样性3个层面多样性指标的相关性。
-
海口市地处海南岛北部(19°31′~20°04′N,110°07′~110°42′E),总面积3 134.84 km2,其中陆地面积2 304.84 km2。海口市属热带海洋性气候。年日照时间相对较长,年平均日照时数2 000 h以上。年平均气温约24.4 ℃,其中平均最低气温为18.0 ℃,最高气温为28.0 ℃。年平均降水量1 696.6 mm,平均年蒸发量为1 834.0 mm,年平均相对湿度为85%。截至2017年年末,海口市已经建成城市园林绿地面积5 736 hm2,建成区绿化覆盖率达40.8%,人均公共绿地面积12.3 m2[10]。
-
当在城市中随机建立的样方数量超过200个(样方面积为400 m2),对植被的数量及结构特征估计的标准差约12%,对一座城市的植物群落调查相对比较充分[11]。本研究采用样方调查期内的海口市卫星遥感影像,划定城市主建成区,面积约123.6 km2。在实地调查过程中,考虑到若干样方的落点位置无法进入,总共随机设立了235个样方。
在地面调查中,采用卫星影像以及手持式全球定位系统(GPS)对样方的中心进行定位,12 m作为半径,建立圆形样方,收集植物群落数据。数据内容包括样方中心坐标,乔木(胸径≥3 cm)种类、胸径、株数、树高、冠幅等,灌木的种类、高度和面积,草本的种类、高度和面积。以《海南植物名录》作为物种来源的参考依据进行判别和确定[12]。记录的植物种类不含栽培的农作物、果蔬花卉品种以及温室引种的种类。
-
物种在群落中的优势水平用重要值表示,以反映海口建成区内基调植物种类。公式如下[13]:乔木层重要值=(相对密度+相对显著度+相对频度)/3,灌木层重要值=(相对高度+相对盖度+相对频度)/3,草本层重要值=(相对高度+相对盖度+相对频度)/3,相对重要值=该物种的重要值/该样地内所有物种的重要值总和。划分群落类型能够有效地表示城市森林中重要的优势群落外貌结构、群落种类和特征。以海口市建成区为例,将数据集中的乔木相对重要值作为分析基础,运用SPSS 20.0的组间联结聚类方法进行系统聚类[14],以群落的外貌特征和聚类树状图作为划分依据,进行群落类型的划分。
-
选取4类普遍应用的α物种多样性指数。Patrick物种丰富度指数:R=S,Shannon-Wiener指数:$H=-\sum\limits_{i=1}^{s}\left(P_{i} \ln P_{i}\right) $,Simpson指数:$ D=1-\sum\limits_{i=1}^{s}\left(\frac{n_{i}}{N}\right)^{2}$,Pielou均匀度指数:$ J=\frac{H}{\ln S}$。其中:ni表示物种的个体数量,S表示物种的数量用,N物种个体数的总和,Pi=ni/N。
-
针对获取数据的便利性和城市环境的特殊性,选择和城市环境有关的13种功能性状指标,其内容包括植物的生活类型、株高、传粉方法、花期、植物来源,种子的扩散方式、体积、数量;植株能否常绿,能否进行有性繁殖,植物是否有毒,茎秆是否带刺等。选取的功能性状指标根据WILLIAMS等[15]以及KNAPP等[16]研究中城市植物功能性状研究体系。通过《中国植物志》《海南植物志》及《广东植物志》查找植物功能性状数据。选取4种功能多样性指数来测度研究区域内的群落功能多样性:功能均匀度指数(FEve)、功能丰富度指数(FRic)、功能离散度指数(FDis)和功能分散度指数(FDiv)。通过R.3.2.2的FD软件包中的dbFD函数完成功能多样性指数计算。
-
将调查数据中的物种和对应的科属信息按照APG Ⅲ(被子植物分类系统Ⅲ)输入在线植物系统发育库Phylomatic v3中[17],导出系统发育树文件。选取3种类型的谱系多样性指数:谱系多样性指数(PD)、谱系变异性指数(PSV)及谱系均匀度指数(PSE)。PD指数反映的是数据集中所有物种在系统发育进化树上的枝长之和[18],值越大表示所包含物种的系统发育信息越丰富。PSV指数能够把群落中全部物种都共同拥有的一个随机性状产生的变异水平进行量化,并反映出是如何通过系统发育亲缘关系进行变化的。PSE指数是对PSV指数进行改进的指标,通过改进能够融合更多的物种信息[19]。为分析α多样性水平上物种多样性、功能多样性和谱系多样性指数之间的相关性,通过Pearson线性相关进行分析。相关系数和显著性通过R.3.2.2的psych软件包中的corr. test函数分析完成。
-
如表 1所示:海口市建成区植物调查中,记录了72科218属,共283种物种。其中,乔木87种,灌木77种,草本119种。植物来源分析中发现,乡土种114种,外来种169种,栽培种131种,外来种和乡土种的数量比为3:2,栽培种在外来种当中所占的比例最高,达77.51%。
表 1 海口建成区城市森林植物来源数据汇总表
Table 1. Statistics of plant origin in the built-up area of Haikou City
生活型 总种数/种 乡土种 外来种 栽培种占外来种数比例/% 种数/种 占总种数比例/% 种数/种 占总种数比例/% 乔木 87 32 36.78 55 63.21 94.54 灌木 77 24 31.17 53 68.83 90.57 草本 119 56 47.06 63 52.94 49.21 总计 283 114 40.28 169 59.71 77.51 -
如图 1所示:优势乔木包含椰子Cocos nucifera,印度紫檀Pterocarpus indicus,王棕Roystonea regia和垂叶榕Ficus benjamina等;优势灌木包含龙船花Ixora chinensis,黄金榕Ficus microcarpa ‘GoldenLeaves’和扶桑Hibiscus rosa-sinensis等;优势草本包含蟛蜞菊Sphagneticola calendulacea,牛筋草Eleusine indica,细叶结缕草Zoysia tenuifolia,结缕草Zoysia japonica等。在所有乔木物种重要值总数中,乔木层排序前10位的乔木物种,其重要值之和占比为48.74%,灌木层前10位重要值之和占比为54.15%,草本层前10位重要值之和占比为47.37%。

图 1 海口市建成区重要值前10位的植物种类
Figure 1. Important values of the top 10 plants in the built-up area of Haikou City
-
以聚类分析的结果为依据,把植物群落划分成20个主要类型,各类型用乔木层优势种命名,如表 2所示。印度紫檀群落、椰子群落和垂叶榕群落是出现频率最高的群落类型,约占样方总数1/3。
表 2 海口市建成区城市森林主要群落类型
Table 2. Urban forest main community types in the built-up area of Haikou City
编号 群落名 C1 印度紫檀群落 C2 椰子群落 C3 垂叶榕群落 C4 小叶榄仁Terminalia boivinii+鸡蛋花Plumeria rubra‘Acutifolia’群落 C5 王棕群落 C6 王棕+小叶榕Ficus microcarpa群落 C7 高山榕Ficus altissima群落 C8 凤凰木Delonix regia群落 C9 苦楝Melia azedarach+番木瓜Carica papaya群落 C10 樟树Cinnamomum camphora群落 C11 秋枫Bischofia javanica群落 C12 非洲楝Khaya senegalensis群落 C13 黄葛榕Ficus virens群落 C14 阳桃Averrhoa carambola群落 C15 木麻黄Casuarina equisetifolia群落 C16 苦楝+糖胶树Alstonia scholaris群落 C17 苦楝群落 C18 黄槐Senna surattensis群落 C19 潺槁木姜子Litsea glutinosa群落 C20 乌桕Triadica sebifera群落 -
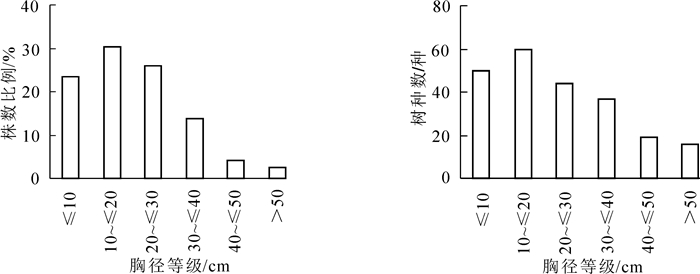
如图 2所示:在径级结构中,占比最高的树木胸径等级为10~20 cm,为30.25%,胸径<20 cm的树木胸径占比53.6%;胸径<20 cm的物种数量较多,胸径等级在<10 cm和10~20 cm的物种数量分别是50种和60种。随着胸径等级的增加,株数所占的比例和植株数量都逐步下降,胸径>50 cm的物种仅为16种,占总数比例的2.45%,主要物种包括秋枫,红花天料木Homalium hainanense,樟树,小叶榕,高山榕,非洲楝等。乔木层所有植株的平均胸径为21.7 cm。

图 2 不同胸径等级上的乔木树种数量和种数分布
Figure 2. Distribution of tree individuals and tree species in different DBH classes
如图 3所示:在立木层方面,植株高度在<5 m以及5~10 m的株数占比分别是34.56%和50.47%,树种分别为63种和70种,为主要立木垂直分布层。>15 m的乔木仅有7种物种,株数占总数比例的1.42%,包括糖胶树,榄仁,王棕,大叶桃花心木Swietenia macrophylla,非洲楝,木麻黄,白兰Michelia alba等树种。乔木层所有植株的平均高度只有6.8 m。

图 3 不同立木层次上的乔木树种数量和种数分布
Figure 3. Distribution of tree individuals and tree species in different tree stratums
如表 3所示:FRic指数与J指数在各种生活型中均不存在显著相关。在各生活型中,FEve指数和FDis指数都与物种多样性指数在0.1%统计水平上极显著正相关(P<0.001)。在灌木层和草本层中,FDiv指数与J指数不存在显著相关,但与其他物种多样性指数的相关系数至少在5%的统计水平上显著。
表 3 物种多样性指数与功能多样性指数的Pearson相关系数
Table 3. Pearson correlation coefficients between species diversity indices and functional diversity indices
功能多样性指数 乔木层 灌木层 草本层 R D H J R D H J R D H J FRic 0.77*** 0.62*** 0.72*** 0.11 0.86*** 0.56*** 0.71*** 0.18 0.85*** 0.54*** 0.69*** 0.14 FEve 0.47*** 0.78*** 0.67*** 0.44*** 0.43*** 0.73*** 0.70*** 0.52*** 0.45*** 0.74*** 0.72*** 0.52*** FDiv 0.47*** 0.74*** 0.65*** 0.36*** 0.34* 0.24* 0.31** -0.04 0.34*** 0.24** 0.31*** -0.04 FDis 0.44*** 0.64*** 0.56*** 0.47*** 0.40*** 0.85*** 0.78*** 0.78*** 0.37*** 0.85*** 0.78*** 0.78*** 说明:*P<0.05,**P<0.01,***P<0.001 如表 4所示:PD指数在灌木层中与各个物种多样性指数均不存在显著相关;在草木层中,与J不存在显著相关。PSV指数仅在乔木层中与D显著正相关(P<0.05)。PSE指数在灌木层中与各个谱系多样性指数均不存在显著的相关性。
表 4 物种多样性指数和谱系多样性指数的Pearson相关系数
Table 4. Pearson correlation coefficients between species diversity indices and phylogenetic diversity indices
谱系多样性指数 乔木层 灌木层 草本层 R D H J R D H J R D H J PD 0.93*** 0.77*** 0.89*** 0.19* -0.15 -0.08 -0.09 -0.01 0.90*** 0.53*** 0.70*** 0.13 PSV 0.11 0.18* 0.15 0.11 -0.16 -0.15 -0.17 -0.10 0.11 0.03 0.06 -0.07 PSE 0.27** 0.55*** 0.42*** 0.58*** -0.19 -0.10 -0.13 -0.03 0.20* 0.78*** 0.66*** 0.84*** 说明:*P<0.05,**P<0.01,***P<0.001 如表 5所示:PD指数与灌木层中与各个功能多样性指数均不存在显著相关。PSV指数与各功能多样性指数的相关性在各个生活型中均有差异,在乔木层中与FEve指数和FDiv指数显著正相关(P<0.05),在灌木层中仅与FDiv指数显著负相关(P<0.05),在草本层中与FDiv指数和FDis指数显著正相关(P<0.01,P<0.05)。PSE指数在乔木层中与FRic指数不存在显著相关,灌木层中仅与FRic指数显著正相关(P<0.05),草本层中仅与FDiv指数不存在显著相关。
表 5 功能多样性指数与谱系多样性指数的Pearson相关系数
Table 5. Pearson correlation coefficients between functional diversity indices and phylogenetic diversity indices
谱系多样性指数 乔木层 灌木层 草本层 FRic FEve FDiv FDis FRic FEve FDiv FDis FRic FEve FDiv FDis PD 0.66*** 0.49*** 0.48*** 0.41*** -0.15 -0.12 -0.13 -0.05 0.82*** 0.50*** 0.44*** 0.42*** PSV 0.02 0.18* 0.18* 0.15 -0.08 -0.15 -0.22* -0.11 0.13 0.16 0.25** 0.20* PSE 0.16 0.43*** 0.38*** 0.42*** -0.24* -0.06 -0.11 -0.11 0.24** 0.58*** 0.07 0.84*** 说明:*P<0.05,**P<0.01,***P<0.001 -
本研究记录了海口市城市森林植物种类283种。有研究统计,海口市行政区范围内有1 588种植物种类(不包括果蔬、花卉、农作物、经济植物以及温室引种的各类植物),其中有40种裸子植物,归属于8科14种,有28种蕨类植物,归属于15科20种,有1 520种被子植物,归属于149科814属[20]。城市调查区域和取样方法的不同,都会使结果产生差异[21-22]。物种普查方法在收集植物种类上比样方调查效率更高,但会缺失对物种多度信息的采集量化。
外来物种在城市中的扩张已经是众多研究的统一结果[23-24]。植物来源分析显示,乡土种和外来种的比例为2:3。其主要原因是外来物种在城市环境中使用的成本效益等同于或甚至优于乡土物种,且对城市生境有较高的适应能力[25],因此外来物种成为更加有效率的选择[26]。尽管外来物种的引入在一定时期内可能会提升生物多样性水平,但是外来物种的大量使用和逐步归化会加剧城市森林中群落水平上的同质化现象[27]。选择乡土物种进行城市森林群落营造,不仅能突出地域景观特色,有效缓解城市植物群落同质化现象,而且有助于提高城市森林的物种丰富度[24]。
-
海口城市森林优势树种与广东省湛江市[28]、深圳市[29]极为相似,均包括椰子、王棕、小叶榕、垂叶榕、高山榕、黄葛榕等物种。这反映出地理区位邻近的城市可能会导致优势物种选择趋同。海口市的优势灌木大多是像龙船花、黄金榕、朱蕉Cordyline fruticosa、扶桑等花型花色上颇有特点的栽培物种。由于城市绿化建设过程中的人为选择偏好,栽培种通常都会具有这些特征[30]。优势草本植物多为草坪草(如结缕草、地毯草Axonopus compressus)和热带杂草(牛筋草、蟛蜞菊),其成因主要是由于杂草和1年生草本形成的种群在城市中能够很好地抵抗干扰[31],修剪或者随意踩踏等人为干扰很难对此类植物种群带来有效控制[32]。
聚类分析结果显示:海口市的植物群落主要由垂叶榕群落、椰子群落和印度紫檀群落构成。需要关注的是在海口市常有台风发生,印度紫檀抗风能力较弱[33],可适当减少将其作为在海口城市景观基调树种,从而减少台风侵袭带来的不必要损失。乔木的空间结构分析表明,海口城市森林整体立木层不高,分布比较单调,多数植株还有较大物理生长空间。
-
物种多样性、功能多样性和谱系多样性的相关指数目前已用于城市化对城市植物多样性的影响研究中,如城市化进程与树种功能多样性的关系[34],城市空间格局对本地和外来植物群落物种丰富度及组成的影响等[35],但这3类指数在不同的研究尺度、格局和地域条件下表现出不同程度的关联性,因此需要更多的实际案例来探讨三者之间的内在关联和成因。
根据本研究结果,城市环境的功能多样性指数和物种多样性指数有显著相关性,同样,在自然环境的案例中也有类似的结果[36-37]。但在自然环境中部分不相关的指数,在本研究中存在显著正相关,比如在长白山植物群落的研究中显示,H指数和功能多样性指数没有相关性[38],这表明城市环境会对物种多样性指数和功能多样性指数的关联程度造成影响,人为选择与观赏有关的功能性状(如高度、花期、常绿与否等)可能是造成这一结果的重要原因。值得一提的是,物种多样性、功能多样性的多数指数与PSV指数均无相关性,但却多与PSE指数显著相关。PSE指数是融合了相对物种多度信息的PSV指数的改进指数[19],表明计算公式中是否考虑多度信息会影响这些指数间的相关性。此外,仅在灌木层中,多数物种、功能多样性指数与谱系多样性指数之间无相关性。不同生活型植物的系统发育结构具有差异,从而会使植物群落构建的主导因素产生区别[38],在研究指数相关性和影响因素时,有必要划分群落中的植物生活型进行针对性研究。
物种、功能和谱系方面的指数对城市植物群落的评估和理解各有用途。虽然直接决定群落构建的是物种的功能性状,但谱系多样性也常用于反映群落的功能多样性[39-40]。在城市环境中,主观选择与特定环境相关的功能性状对植物群落的功能性进行解释,可能比谱系多样性更具有针对性。但谱系多样性避免了在测定功能多样性时人为选择功能性状的主观影响,当关键的功能性状数据缺失时,其适用性可能优于功能多样性[41]。
-
海口市森林植物结构仍然有较大的提升空间,城市植物群落反映出明显的物种选择偏好。此外,与临近城市的物种同质化现象是在未来海口城市森林建设中亟待解决的问题。筛选城市基调树种时,提倡采用与城市文化相关或气候环境相适应的乡土物种,使城市景观风貌区别于周边城市,并能够持续健康的生长发育。因多样性指数和生活型的选取差别,指数间的相关性也有所差异,因此,需要进一步验证城市森林各多样性指标之间的关联性和有效性,以更为全面而准确地为城市森林生态服务功能提升提供决策依据。
Urban forest community structure and correlation analysis on the plant diversity index in Haikou City, China
-
摘要: 选取海口市主建成区随机建立的235个植物样方进行调查,评价其结构特征,分析其植物多样性指数相关性。结果表明:①主建成区常见植物共283种隶属于72科218属,其中,本地种114种,外来种169种;②优势植物主要有椰子Cocos nucifera、印度紫檀Pterocarpus indicus、王棕Roystonea regia、垂叶榕Ficus benjamina等;③树木径级结构呈现"整体偏小"的分布格局,整体立木层次较低,平均树高仅6.8 m;④物种多样性指数与功能多样性指数之间多数存在显著相关(P < 0.05);⑤谱系多样性指数(PD)与乔木层、草木层中各物种多样性指数多数存在显著相关(P < 0.05);谱系变异性指数(PSV)与多数物种多样性指数、功能多样性指数均不相关,但谱系均匀度指数(PSE)与多数物种多样性指数、功能多样性指数显著相关(P < 0.05)。为避免物种同质化现象,提倡运用乡土物种进行城市森林建设,同时选择多层次且具有针对性的多样性指数和功能性状参数,以保证对城市森林植物多样性评估的全面性和准确性。Abstract: In order to have a more comprehensive understanding of the structural change and succession process of plant community in Haikou City, and predict the composition of its future change trend. A total of 235 randomly selected urban forest plots were studied in a built-up area of Haikou City. Structural characteristics and their corresponding plant diversity indexes were determined with a correlation analysis. Results indicated that (1) The presence of 283 species of common plants in the built-up area belonging to 218 genera of 72 families with 114 native species and 169 exotic species. (2) The dominant plants included Cocos nucifera, Pterocarpus indicus, Roystonea regia, and Ficus benjamina. (3) The diameter presented a distribution pattern of "overall small", and the overall tree height was low with an average height of 6.8 m. (4) There was a significant correlation between species diversity and functional diversity indexes (P < 0.05). (5) Also, the phylogenetic diversity index (PD) was significantly correlated (P < 0.05) with species diversity indexes in the tree layer and herb layer. The phylogenetic species variability index (PSV) was not correlated with most species diversity indexes and functional diversity indexes, but the phylogenetic species variability evenness (PSE) index was significantly correlated with most of them (P < 0.05). Haikou City having a large improvement potential in forest development, should avoid the homogeneity phenomenon where native species have been adopted in urban forest development; to ensure comprehensiveness and accuracy, Haikou should consider multi-level and targeted diversity indexes and functional trait parameters when evaluating urban plant diversity.
-
Key words:
- forest ecology /
- community type /
- dominant species /
- species diversity /
- functional diversity /
- phylogenetic diversity
-
图 1 海口市建成区重要值前10位的植物种类
Figure 1 Important values of the top 10 plants in the built-up area of Haikou City
图 2 不同胸径等级上的乔木树种数量和种数分布
Figure 2 Distribution of tree individuals and tree species in different DBH classes
图 3 不同立木层次上的乔木树种数量和种数分布
Figure 3 Distribution of tree individuals and tree species in different tree stratums
表 1 海口建成区城市森林植物来源数据汇总表
Table 1. Statistics of plant origin in the built-up area of Haikou City
生活型 总种数/种 乡土种 外来种 栽培种占外来种数比例/% 种数/种 占总种数比例/% 种数/种 占总种数比例/% 乔木 87 32 36.78 55 63.21 94.54 灌木 77 24 31.17 53 68.83 90.57 草本 119 56 47.06 63 52.94 49.21 总计 283 114 40.28 169 59.71 77.51  下载: 导出CSV
下载: 导出CSV
表 2 海口市建成区城市森林主要群落类型
Table 2. Urban forest main community types in the built-up area of Haikou City
编号 群落名 C1 印度紫檀群落 C2 椰子群落 C3 垂叶榕群落 C4 小叶榄仁Terminalia boivinii+鸡蛋花Plumeria rubra‘Acutifolia’群落 C5 王棕群落 C6 王棕+小叶榕Ficus microcarpa群落 C7 高山榕Ficus altissima群落 C8 凤凰木Delonix regia群落 C9 苦楝Melia azedarach+番木瓜Carica papaya群落 C10 樟树Cinnamomum camphora群落 C11 秋枫Bischofia javanica群落 C12 非洲楝Khaya senegalensis群落 C13 黄葛榕Ficus virens群落 C14 阳桃Averrhoa carambola群落 C15 木麻黄Casuarina equisetifolia群落 C16 苦楝+糖胶树Alstonia scholaris群落 C17 苦楝群落 C18 黄槐Senna surattensis群落 C19 潺槁木姜子Litsea glutinosa群落 C20 乌桕Triadica sebifera群落
下载: 导出CSV
表 3 物种多样性指数与功能多样性指数的Pearson相关系数
Table 3. Pearson correlation coefficients between species diversity indices and functional diversity indices
功能多样性指数 乔木层 灌木层 草本层 R D H J R D H J R D H J FRic 0.77*** 0.62*** 0.72*** 0.11 0.86*** 0.56*** 0.71*** 0.18 0.85*** 0.54*** 0.69*** 0.14 FEve 0.47*** 0.78*** 0.67*** 0.44*** 0.43*** 0.73*** 0.70*** 0.52*** 0.45*** 0.74*** 0.72*** 0.52*** FDiv 0.47*** 0.74*** 0.65*** 0.36*** 0.34* 0.24* 0.31** -0.04 0.34*** 0.24** 0.31*** -0.04 FDis 0.44*** 0.64*** 0.56*** 0.47*** 0.40*** 0.85*** 0.78*** 0.78*** 0.37*** 0.85*** 0.78*** 0.78*** 说明:*P<0.05,**P<0.01,***P<0.001
下载: 导出CSV
表 4 物种多样性指数和谱系多样性指数的Pearson相关系数
Table 4. Pearson correlation coefficients between species diversity indices and phylogenetic diversity indices
谱系多样性指数 乔木层 灌木层 草本层 R D H J R D H J R D H J PD 0.93*** 0.77*** 0.89*** 0.19* -0.15 -0.08 -0.09 -0.01 0.90*** 0.53*** 0.70*** 0.13 PSV 0.11 0.18* 0.15 0.11 -0.16 -0.15 -0.17 -0.10 0.11 0.03 0.06 -0.07 PSE 0.27** 0.55*** 0.42*** 0.58*** -0.19 -0.10 -0.13 -0.03 0.20* 0.78*** 0.66*** 0.84*** 说明:*P<0.05,**P<0.01,***P<0.001
下载: 导出CSV
表 5 功能多样性指数与谱系多样性指数的Pearson相关系数
Table 5. Pearson correlation coefficients between functional diversity indices and phylogenetic diversity indices
谱系多样性指数 乔木层 灌木层 草本层 FRic FEve FDiv FDis FRic FEve FDiv FDis FRic FEve FDiv FDis PD 0.66*** 0.49*** 0.48*** 0.41*** -0.15 -0.12 -0.13 -0.05 0.82*** 0.50*** 0.44*** 0.42*** PSV 0.02 0.18* 0.18* 0.15 -0.08 -0.15 -0.22* -0.11 0.13 0.16 0.25** 0.20* PSE 0.16 0.43*** 0.38*** 0.42*** -0.24* -0.06 -0.11 -0.11 0.24** 0.58*** 0.07 0.84*** 说明:*P<0.05,**P<0.01,***P<0.001
下载: 导出CSV
-
[1] 王成, 彭镇华, 陶康华.中国城市森林的特点及发展思考[J].生态学杂志, 2004, 23(3):88-92. WANG Cheng, PENG Zhenhua, TAO Kanghua. Characteristics and development of urban forest in China[J]. Chin J Ecol, 2004, 23(3):88-92. [2] 赵煜, 赵千钧, 崔胜辉, 等.城市森林生态服务价值评估研究进展[J].生态学报, 2009, 29(12):6723-6732. ZHAO Yu, ZHAO Qianjun, CUI Shenghui, et al. Progress in ecological services evaluation of urban forest[J]. Acta Ecol Sin, 2009, 29(12):6723-6732. [3] 马冰倩, 徐程扬, 刘江, 等.城镇森林视觉景观异质性对美学质量的影响[J].浙江农林大学学报, 2019, 36(2):366-374. MA Bingqian, XU Chengyang, LIU Jiang, et al. Visual heterogeneity and visual landscape quality of urban forests as an architectural backdrop[J]. J Zhejiang A&F Univ, 2019, 36(2):366-374. [4] 陈文静.厦门城市森林的结构特征与效益评价研究[D].福州: 福建农林大学, 2007. CHEN Wenjing. The Study of Structure Characteristics and Effectiveness Evaluation of Urban Forest in Xiamen City[D]. Fuzhou: Fujian Agriculture and Forestry University, 2007. [5] DÍAZ S, CABIDO M. Vive la difference:plant functional diversity matters to ecosystem processes[J]. Trends Ecol Evol, 2001, 16(11):646-655. [6] CADOTTE M W. Experimental evidence that evolutionarily diverse assemblages result in higher productivity[J]. Proc Natl Acad Sci, 2013, 110(22):8996-9000. [7] LAVOREL S, GARNIER E. Predicting changes in community composition and ecosystem functioning from plant traits:revisiting the Holy Grail[J]. Funct Ecol, 2002, 16(5):545-556. [8] FOREST F, GRENYER R, ROUGET M, et al. Preserving the evolutionary potential of floras in biodiversity hotspots[J]. Nature, 2007, 445(7129):757-760. [9] CUMMING G S, CHILD M F. Contrasting spatial patterns of taxonomic and functional richness offer insights into potential loss of ecosystem services[J]. Philos Trans Royal Soc London B Biol Sci, 2009, 364(1524):1683-1692. [10] 海口市统计局, 国家统计局海口调查队.海口市统计年鉴2018[M].北京:中国统计出版社, 2018. [11] NOWAK D J, WALTON J T, STEVEN J C, et al. Effect of plot and sample size on timing and precision of urban forest assessments[J]. Arboric Urban For, 2008, 34(6):386-390. [12] 杨小波.海南植物名录[M].北京:科学出版社, 2013. [13] 雷金睿, 宋希强, 陈宗铸.海口城市公园植物群落多样性研究[J].西南林业大学学报, 2017, 37(1):88-93. LEI Jinrui, SONG Xiqiang, CHEN Zongzhu. Analysis on plant community diversity in Haikou City parks[J]. J Southwest For Univ, 2017, 37(1):88-93. [14] 张艳丽, 李智勇, 杨军, 等.杭州城市绿地群落结构及植物多样性[J].东北林业大学学报, 2013, 41(11):25-30. ZHANG Yanli, LI Zhiyong, YANG Jun, et al. Community structure and species diversity of urban green space in Hangzhou, China[J]. J Northeast For Univ, 2013, 41(11):25-30. [15] WILLIAMS N S G, HAHS A K, VESK P A. Urbanisation, plant traits and the composition of urban floras[J]. Perspect Plant Ecol Evol Syst, 2015, 17(1):78-86. [16] KNAPP S, KCOHN I, BAKKER J P. How species traits and affinity to urban land use control large-scale species frequency[J]. Diversity Distrib, 2009, 15(3):533-546. [17] WEBB C O, DONOGHUE M J. Phylomatic:tree assembly for applied phylogenetics[J]. Mol Ecol Note, 2005, 5(1):181-183. [18] FAITH D P. Conservation evaluation and phylogenetic diversity[J]. Biol Conserv, 1992, 61(1):1-10. [19] HELMUS M R, BLAND T J, WILLIAMS C K, et al. Phylogenetic measures of biodiversity[J]. Am Nat, 2007, 169(3):E68-E83. doi:10. 1086/511334. [20] 杨小波.城市植物多样性[M].北京:中国农业出版社, 2009. [21] PYŠEK P. Factors affecting the diversity of flora and vegetation in central European settlements[J]. Vegetatio, 1993, 106(1):89-100. [22] HE Rongxiao, YANG Jun, SONG Xiqiang. Quantifying the impact of different ways to delimit study areas on the assessment of species diversity of an urban forest[J]. Forests, 2016, 7(2):42. doi:10.3390/f7020042. [23] MCKINNEY M L. Measuring floristic homogenization by non-native plants in North America[J]. Global Ecol Biogeogr, 2004, 13(1):47-53. [24] KÜHN I, BRANDL R, KLOTZ S. The flora of German cities is naturally species rich[J]. Evol Ecol Res, 2004, 6(5):749-764. [25] MCKINNEY M L. Urbanization as a major cause of biotic homogenization[J]. Biol Conserv, 2006, 127(3):247-260. [26] DEARBORN D C, KARK S. Motivations for conserving urban biodiversity[J]. Conserv Biol, 2010, 24(2):432-440. [27] ZDEŇKA L, CHYTRY M, LUBOMÍR Tichy, et al. Biotic homogenization of central European urban floras depends on residence time of alien species and habitat types[J]. Biol Conserv, 2012, 145(1):179-184. [28] 吴刘萍.湛江市城市森林树种结构分析[J].亚热带植物科学, 2007, 36(2):31-35. WU Liuping. The analysis of species composition of urban forest in Zhanjiang City[J]. Subtrop Plant Sci, 2007, 36(2):31-35. [29] 宋艳暾, 史志华, 余世孝, 等.城市化对绿地植物组成特征的影响:以深圳为例[J].生态环境学报, 2010, 19(3):615-620. SONG Yantun, SHI Zhihua, YU Shixiao, et al. Influence of urbanization on the characters of plants composition in urban greenspace in Shenzhen, China[J]. Ecol Environ Sci, 2010, 19(3):615-620. [30] KENDAL D, WILLIAMS K J H, WILLIAMS N S G. Plant traits link people's plant preferences to the composition of their gardens[J]. Landscape Urban Plann, 2012, 105(1):34-42. [31] 彭羽, 刘雪华.城市化对植物多样性影响的研究进展[J].生物多样性, 2007, 15(5):558-562. PENG Yu, LIU Xuehua. Research progress in effects of urbanization on plant biodiversity[J]. Biodiversity Sci, 2007, 15(5):558-562. [32] 陈晓双, 梁红, 宋坤, 等.哈尔滨城区杂草群落多样性及其分类体系[J].应用生态学报, 2014, 25(8):2221-2228. CHEN Xiaoshuang, LIANG Hong, SONG Kun, et al. Diversity and classification system of weed community in Harbin City, China[J]. Chin J Appl Ecol, 2014, 25(8):2221-2228. [33] 罗冠勇, 宋希强, 杨冬华, 等.海南10种园林乔木生物学特性与抗风性关联性分析[J].热带作物学报, 2013, 34(2):263-267. LUO Guanyong, SONG Xiqiang, YANG Donghua, et al. Correlation analysis on the relationship between the biological characteristic of ten ornamental tree species and the wind-resistance ability in Hainan Island[J]. Chin J Trop Crops, 2013, 34(2):263-267. [34] NOCK C A, PAQUETTE A, FOLLETT M, et al. Effects of urbanization on tree species functional diversity in Eastern North America[J]. Ecosystems, 2013, 16(8):1487-1497. [35] ŇEPLOVÁN, LOSOSOVÁZ, ZELENŇ D, et al. Phylogenetic diversity of central-European urban plant communities:effects of alien species and habitat types[J]. Preslia, 2015, 87(1):1-16. [36] 么旭阳, 胡耀升, 刘艳红.长白山阔叶红松林不同群落类型的植物功能性状与功能多样性[J].西北农林科技大学学报(自然科学版), 2014, 42(3):77-84. YAO Xuyang, HU Yaosheng, LIU Yanhong. Plant functional traits and functional diversities of different communities in broad-leaved Korean pine forests in the Changbai Mountain[J]. J Northwest A&F Univ Nat Sci Ed, 2014, 42(3):77-84. [37] 薛倩妮, 闫明, 毕润成.山西五鹿山森林群落木本植物功能多样性[J].生态学报, 2015, 35(21):7023-7032. XUE Qianni, YAN Ming, BI Runcheng. Functional diversity research of tree and shrub layers in forest communities of the Wulu Mountains Nature Reserve in Shanxi, China[J]. Acta Ecol Sin, 2015, 35(21):7023-7032. [38] 丁洪波, 吴兆录, 吕东蓬, 等.云南东部山区不同类型次生林群落谱系结构特征[J].生态学杂志, 2015, 34(10):2720-2726. DING Hongbo, WU Zhaolu, LÜ Dongpeng, et al. Community phylogenetic structural characteristics of various secondary forests in mountainous eastern Yunnan[J]. Chin J Ecol, 2015, 34(10):2720-2726. [39] WEBB C O, ACKERLY D D, MCPEEK M A, et al. Phylogenies and community ecology[J]. Annual Rev Ecol Syst, 2002, 8(33):475-505. [40] SILVERTOWN J, DODD M E, GOWING D J G. Phylogeny and the niche structure of meadow plant communities[J]. J Ecol, 2001, 89(3):428-435. [41] 贾鹏, 杜国祯.生态学的多样性指数:功能与系统发育[J].生命科学, 2014, 26(2):153-157. JIA Peng, DU Guozhen. Measuring functional and phylogenetic diversity in community ecology[J]. Chin Bull Life Sci, 2014, 26(2):153-157. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2019.06.011
 点击查看大图
点击查看大图

计量
- 文章访问数: 5189
- HTML全文浏览量: 1383
- PDF下载量: 130
- 被引次数: 0
