





-
木材作为环境友好型的绿色原材料,是国民经济发展的基础材料。为坚持生态发展战略,中国林业相关部门启动了一批重点生态工程,在天然林保护的同时,大力发展人工林种植。随着国内民众生态意识的增强和国家森林保护力度的加大,木材刚性需求增加,木材的对外依赖度达到了近50%。据测算,到2020年,中国木材年需求量将达到8亿m3,对大径材和珍贵树种用材需求也会大幅增加[1]。引进国外优良树种是提高林业生产力、填补国内名特优新经济林、满足人们对林产品需求的有效途径,对于在短期内实现有效供给、缓解木材及林产品的供需矛盾,实现可持续发展,具有十分重要的意义。中国土壤气候条件与北美相似,引种美国红橡树种具有得天独厚的优势。引种树种能在短时间内增加当地的植物种类,树种的合理布局和应用,对弥补优质用材资源不足有重要意义。橡树又称栎树,是壳斗科Fagaceae栎属Quercus树种的统称,用途广泛,材质优异,是优质商品材和燃料的重要来源[2]。中国自1998年开展北美栎树种质资源引种与栽培研究,对引种试验、引种树苗抗逆性[3]研究较多,为栎类树种引进开发利用提供了理论基础。纤维形态特征与木材生长状况以及强度、密度以及物理、力学性质有密切关系,可用来进行木材的材性预测。微纤丝角在一定程度上可以反映木材的物理和力学性质,如木材的尺寸稳定性、干缩湿胀及弹性模量、强度和蠕变特性等,是评价木材品质性能和良种选育的重要依据[4]。木材的结晶度与木材密度、强度、尺寸稳定性存在着正相关关系[5],研究结晶度有助于了解木材解剖结构与其性质之间联系。本研究以纳塔栎Quercus nuttallii、水栎Quercus nigra和舒玛栎Quercus shumardii为对象,比较了3种美国红橡纤维形态、微纤丝角和结晶度的异同,以期为引种栎木的科学利用提供理论依据,为更大范围引种栽培提供理论基础。
-
根据GB/T 1927-2009《木材物理力学试材采集方法》采集试样[6]。选取14年生纳塔栎、水栎、舒玛栎各4株;其中纳塔栎、水栎采自江苏省林业科学研究院句容苗圃(年平均气温为15.2 ℃,年降水量为1 058.8 mm)。纳塔栎平均树高为8.48 m,平均胸径为19.28 cm;水栎平均树高为10.68 m,平均胸径为15.38 cm;舒玛栎采自于南京彩树种植有限公司江宁区种植基地(年平均气温为15.4 ℃,年降水量为1 200.0 mm),平均树高为11.70 m,平均胸径为15.20 cm。
-
将样木自胸高处圆盘锯解,立即沿南北向自髓心向外逐年切取样块并依次编号,由于第1,2年轮宽度较窄,合并为1个样品。将各标号木块切成火柴棍大小,每个年轮制4组平行样。用富兰克林离析法[7]离析细胞,制成临时切片,置于普通光学显微镜下;随机测量50个细胞长度、宽度和腔径,计算双壁厚、壁腔比。结果取平均数。
-
将3种样木按1.2.1制成厚度约为2 mm的试样,采用Ultima Ⅳ组合型多功能水平X-射线衍射仪(XRD)按Cave 0.6T法[8]测量试材微纤丝角。结果取平均数。
-
将3种样木按1.2.1制成火柴棍大小试样,磨成80~100目木粉,室温下压成薄片后测定木材结晶度,同时分析木材2θ衍射强度曲线。计算结果取平均数。木材纤维的衍射强度在2θ=22°是极大值,其积分强度用I002表示;在2θ=18°附近出现波谷,是木材纤维中无定形区衍射的散射强度,其积分强度用Iam表示。相对结晶度(ICr)公式[9]如下:ICr=100×(I002-Iam)/I002。
-
试验数据采用Excel 2013进行方差分析及回归分析,Origin软件作图。
-
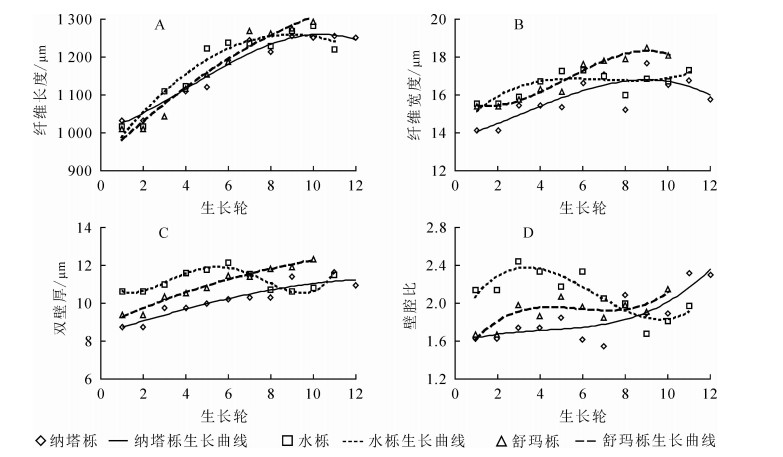
由表 1可知:纳塔栎纤维长度径向变化范围为1 032.46~1 255.46 μm,平均值为(1 172.14±86.88)μm;水栎纤维长度径向变化范围为1 018.04~1 281.27 μm,平均值为(1 178.68±95.54)μm;舒玛栎纤维长度径向变化范围为1 010.34~1 292.56 μm,平均值为(1 162.45±112.67)μm。其中,舒玛栎的平均纤维长度最短,纳塔栎和水栎之间差异较小,三者栎木纤维长度种间不具有显著差异(P>0.05)。按照国际木材解剖学家协会理事会发布的纤维长度分级标准(<0.9 mm的属于短纤维,0.9~1.6 mm的属于中等纤维,>1.6 mm的属于长纤维),3种栎木纤维均属于中等纤维,大于62年生蒙古栎Q. mongolica纤维平均长度(1 001.20 μm)[10],与40年生欧洲栓皮栎Q. suber(960.00~1 220.00 μm)[11]、33年生冬青栎Q. ilex(1 103.00 μm)及24年生深红栎Q. coccifera纤维平均长度(1 116.00 μm)相近[12]。由图 1A可见:3种栎木纤维长度从髓心至树皮方向总体呈上升趋势,但径向变化尚未趋于稳定,说明树木仍处于幼龄期。
表 1 纤维形态的比较分析
Table 1. Comparative analysis of fiber morphology
树种 纤维长度 纤维宽度 双壁厚 壁腔比 变化范围/μm 平均值/μm 变异系数/% P 变化范围/μm 平均值/μm 变异系数/% P 变化范围/μm 平均值/μm 变异系数/% P 变化范围/μm 平均值/μm 变异系数/% P 纳塔栎 1 032.46~1 255.46 1 172.14±86.88 7.41 14.14~17.69 15.86 ± 1.11 7.02 8.77~11.67 10.23 ± 0.91 8.89 1.55~2.32 1.85 ± 0.26 13.97 水栎 1 018.04~1 281.27 1 178.68 ± 95.54 8.11 0.941 560 15.53~17.34 16.56 ± 0.70 4.24 0.062 224 10.65~12.15 11.19 ± 0.54 4.85 0.028 812* 1.68~2.44 2.10 ± 0.23 10.97 0.035 464* 舒玛栎 1010.34~1 292.56 1 162.45 ± 112.67 9.69 15.43~18.51 16.91 ± 1.20 7.08 舒玛栎 9.40~12.34 10.96 ± 1.03 9.37 1.67~2.15 1.91 ± 0.16 8.22 说明:P为各指标平均值的种间差异显著性分析;表示0.05水平上具有显著性差异 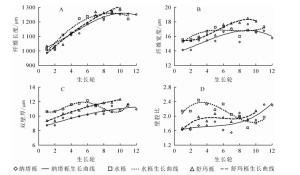
图 1 纤维形态随生长轮的径向变化规律
Figure 1. Radial variation of fiber morphology and growth ring from pith to bark
3种栎木纤维宽度为14.14~18.51 μm,舒玛栎纤维宽度最大(16.91 μm±1.20 μm),水栎(16.56 μm±0.70 μm)和纳塔栎(15.86 μm±1.11 μm)略小,变异系数均小于10%,变异较小。纤维宽度种间差异不显著(P>0.05)。3种栎木纤维宽度均小于40年生欧洲栓皮栎(18.40~21.49 μm)。由图 1B可知:3种栎木纤维宽度的径向变化均随生长轮增加而增加,但存在着一定的波动。
纤维双壁厚是木材质量、基本密度和强度的物质基础[13],受生态环境条件和气候因子的影响较大[14]。研究发现:水栎双壁厚最大(11.19 μm±0.54 μm),纳塔栎(10.23 μm±0.91 μm)和舒玛栎(10.96 μm±1.03 μm)略小,3种栎木双壁厚种间具有显著差异(P<0.05)。由图 1C可知:随生长轮增加,3种栎木纤维双壁厚径向均呈增长趋势。较其他2种栎,水栎更易受生态环境条件和气候因子影响,因而纤维双壁厚变化波动也较大。
纤维壁腔比是纤维双壁厚与纤维胞腔直径的比值,是衡量木材纤维强度的重要指标。纤维壁腔比越小,则木材密度越大,材质越坚硬[15]。木材纤维工业原料中通常要求纤维为中级长度(900.00~1 600.00 μm),长宽比不低于35,壁腔比小于1[16]。本研究发现:3种栎木壁腔比为1.55~2.44,水栎壁腔比平均值最大(2.10±0.23),纳塔栎(1.85±0.26)和舒玛栎(1.91±0.16)略低,纤维壁腔比种间具有显著差异(P<0.05)。3种栎木壁腔比均大于1,未达到纤维工业原料要求。从髓心向外,3种栎木壁腔比径向均略微上升,在一定范围内存在波动(图 1D)。
-
微纤丝角是影响木材强度的主要因素,通常作为材质评定及早期良种选育的重要指标。本研究发现:纳塔栎微纤丝角径向变化范围为30.02°~38.73°,平均值为33.79°。水栎为26.12°~33.52°,平均值为30.48°。舒玛栎为37.65°~30.93°,平均值为34.10°。3种栎木微纤丝角大小依次为舒玛栎(34.10°)、纳塔栎(33.79°)和水栎(30.48°),微纤丝角种间具有极显著差异(P<0.01)。3种栎木微纤丝角平均值为32.79°,略大于采用细胞壁高分辨率显微照片测定的夏栎Q. robur(22°~31.6°)[17]的微纤丝角,可能与栎木未达到成熟状态有关,也可能是因为测量方法不同,XRD法测量时采用的是细胞壁各壁层微纤丝角平均值。
从图 2可以看出:3种栎木微纤丝角随生长轮增加总体呈下降趋势。原因可能是试材越靠近髓心,树木年龄越小,形成层原始细胞产生的子细胞胞壁就越薄,各层的平均微纤丝角就越大[18]。
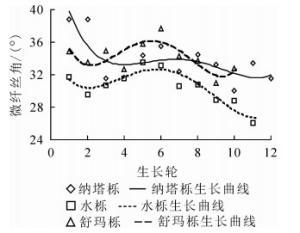
图 2 微纤丝角随生长轮的径向变化规律
Figure 2. Radial variation of microfibril angle and growth ring from pith to bark
-
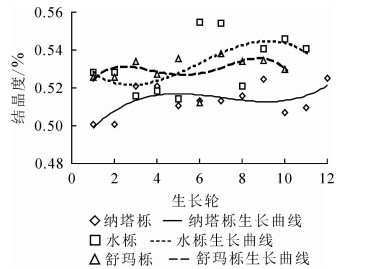
纳塔栎结晶度径向变化范围为50.07%~52.52%,平均值为51.35%。水栎结晶度径向变化范围为51.45%~55.47%,平均值为53.30%。舒玛栎结晶度径向变化范围为51.23%~53.83%,平均值为52.97%。结晶度大小依次为水栎(53.30%)、舒玛栎(52.97%)和纳塔栎(51.35%),均大于加拿大栎Q. canariensis(38.93%)[19],3种栎木种间具有极显著差异(P<0.01)。纳塔栎、水栎和舒玛栎结晶度的变异系数分别为1.63%、2.80%和1.43%,变异较小。如图 3所示:结晶度随生长轮增加总体呈上升趋势,还未处于平稳状态,可能与其处于生长时期有关。结晶度径向变化趋势与其他学者研究一致:在树木生长初期形成层细胞分裂速度较快,结晶度较小,随着树龄的增加,结晶度也逐渐增大最后趋于稳定[20]。
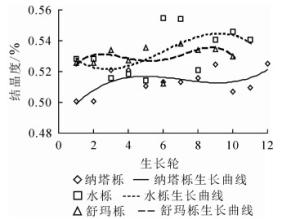
图 3 结晶度随生长轮的径向变化规律
Figure 3. Radial variation of the crystallinity and growth ring from pith to bark
-
纤维形态、微纤丝角与生长轮间关系密切。对生长轮与纤维形态、微纤丝角进行拟合分析,建立适当的模型,对人工林材质早期预测及林木的定向培育、优化利用具有十分重要理论意义和应用价值。以生长轮为自变量(x),纤维长度、纤维宽度、纤维双壁厚、微纤丝角分别为因变量(y),进行回归拟合。
如表 2所示:3种引种栎木纤维长度与生长轮的关系与2次曲线拟合度较好,相关系数(R2)均在0.900以上,回归效果理想,与大多数研究结果一致[21],符合生物学生长规律。3种引种栎木纤维宽度与生长轮的3次曲线拟合度高,其中舒玛栎拟合效果最佳,相关系数达到0.960。纳塔栎和舒玛栎纤维双壁厚与生长轮的2次曲线有较好的拟合关系,水栎纤维双壁厚与生长轮的3次曲线拟合关系较好,其中舒玛栎拟合效果最佳,相关系数达到0.972。纳塔栎微纤丝角与生长轮的关系与对数函数有较好的拟合,水栎及舒玛栎与2次曲线有较好的拟合,其中水栎拟合效果最佳,相关系数为0.748。
表 2 纤维形态、微纤丝角与生长轮的回归分析
Table 2. Regression analysis of fiber morphology, microfibril angle, and growth ring
拟合类型 样木 回归方程 R2 纤维长度与生长轮 纳塔栎 y=-2.004 3x2+48.641 0x+964.540 0 0.943 水栎 y=-4.235 4x2+76.000 0x+917.510 0 0.933 舒玛栎 y=-1.673 2x2+54.666 0x+926.210 0 0.962 纤维宽度与生长轮 纳塔栎 y=-0.004 0x3+0.040 0x2+0.330 0x+13.710 0 0.689 水栎 y=0.889 0x3—0.190 0x2+1.280 0x+14.310 0 0.656 舒玛栎 y=-0.010 0x3+0.220 0x2-0.590 0x+15.832 0 0.960 双壁厚与生长轮 纳塔栎 y=-0.120 0x2+0.400 0x+8.340 0 0.884 水栎 y=-0.116 0x3+0.790 0x2-1.630 0x+l 1.580 0 0.922 舒玛栎 y=-0.020 0x2+0.510 0x+8.780 0 0.972 微纤丝角与生长轮 纳塔栎 y=-2.529 0lnx+38.007 0 0.481 水栎 y=-0.143 9x2+1.356 4x+28.960 0 0.748 舒玛栎 y=-0.110 0x2+0.988 3x+32.896 0 0.337 -
对纳塔栎、水栎和舒玛栎纤维形态测量得知:3种引种栎木纤维长度为1 010.34~1 292.56 μm,纤维宽度为为14.14~18.51 μm,纤维长度、宽度种间不具有显著差异(P>0.05),其径向变化均为从髓心至树皮方向呈上升趋势,但径向变化尚未趋于稳定;3种栎木纤维双壁厚为8.77~12.34 μm,壁腔比为1.55~2.44,纤维双壁厚、壁腔比种间具有显著差异(P<0.05),其径向变化均为从髓心至树皮方向略微上升,在一定范围内存在波动。
3种栎木中舒玛栎微纤丝角最大(34.10°),纳塔栎(33.79°)和水栎(30.48°)略小,微纤丝角种间具有极显著差异(P<0.01)。径向变化为微纤丝角随着生长轮增加而降低;结晶度大小依次为水栎(53.30%)、舒玛栎(52.97%)和纳塔栎(51.35%),结晶度种间具有极显著差异(P<0.01),径向变化为结晶度随生长轮增加总体呈平缓上升趋势。
对生长轮与各指标进行拟合分析发现:3种栎木纤维长度与生长轮关系用2次曲线拟合效果理想,相关系数均在0.900以上。纤维宽度与生长轮关系:舒玛栎用3次曲线拟合效果理想,相关系数在0.900以上。双壁厚与生长轮的关系:舒玛栎用2次曲线拟合效果理想,相关系数在0.900以上。微纤丝角与生长轮的关系:水栎用2次曲线拟合效果较好,相关系数在0.700以上。
由此认为:14年生引种美国红橡仍处于幼龄期,纤维形态、微纤丝角与结晶度径向变化趋势尚未趋于稳定。
Fiber morphology, microfibril angle and crystallinity of American red oak introduced
-
摘要:
目的 为引种栎木的科学利用提供理论依据。 方法 以14年生美国红橡纳塔栎Quercus nuttallii、水栎Quercus nigra、舒玛栎Quercus shumardii为对象,采用富兰克林解离法及X-射线衍射法(XRD)研究其纤维形态、微纤丝角及结晶度,并对数据进行方差分析及回归分析。 结果 3种美国红橡纤维长度分别为1 172.14、1 178.68和1 162.45 μm,纤维宽度分别为15.86、16.56和16.91 μm,种间差异均不显著(P>0.05);双壁厚度分别为10.23、11.19和10.96 μm,壁腔比分别为1.85、2.10和1.91,种间差异均显著(P < 0.05);3种美国红橡的微纤丝角分别为33.79°、30.48°和34.10°,结晶度分别为51.35%、53.30%和52.97%,种间均具有极显著差异(P < 0.01)。3种美国红橡纤维长度、宽度、双壁厚、壁腔比以及结晶度均随生长轮的增加呈增长趋势,微纤丝角随生长轮的增加呈下降趋势,并且年轮间存在着一定的波动。 结论 14年生纳塔栎、水栎和舒玛栎纤维尺寸、微纤丝角、结晶度径向变化尚未稳定,仍处于幼龄期。纤维长度、宽度、双壁厚及微纤丝角与生长轮有较好的拟合关系。 Abstract:Objective Measuring and analyzingthe fiber morphology, microfibril angle and crystallinity can provide a theoretical basis for its quality prediction and utilization. Method Taking 14-year-old Quercus nuttallii, Q. nigra and Q. shumardii as the research object, with the maceration method and X-ray diffractometer (XRD) employed, a study is conducted of their fiber morphology, microfibril angle and crystallinity with a regression and variance analysis made of the data. Result (1) The fiber lengths of the three species of oak are 1 172.14, 1 178.68 and 1 162.45 μm, the fiber widths of the three species of oak are 15.86, 16.56 and 16.91 μm with no significant differences among species (P>0.05). The double-wall thickness of three species of oak are 10.23, 11.19 and 10.96 μm, and the wall thickness-lumen diameter ratios of the three species of oak are 1.85, 2.10 and 1.91 with significant differences among species (P < 0.05). The microfibril angles of three species of oak are 33.79°, 30.48° and 34.10° and the crystallinities of the three oak species are 51.35%, 53.30% and 52.97% with extremely significant differences among species (P < 0.01). (2) The fiber length, the fiber width, the double-wall thickness, the wall thickness-lumen diameter ratio and the crystallinity of three species of oak displayed an ascending trend from pith to bark. The microfibril angle displayed a descending trend from pith to bark with some fluctuations between the rings. Conclusion The radial change of the fiber morphology, the microfibril angle and the crystallinity of 14-years-old Q. nuttallii, Q. nigra and Q. shumardii was not stable but stayed in the juvenile stage. Also, the fiber length, fiber width, double wall thickness and microfibril angle demonstrate favorable fitting with growth rings. -
Key words:
- wood science and technology /
- American red oak /
- fiber morphology /
- microfibril angle /
- crystallinity
-
图 1 纤维形态随生长轮的径向变化规律
Figure 1 Radial variation of fiber morphology and growth ring from pith to bark
图 2 微纤丝角随生长轮的径向变化规律
Figure 2 Radial variation of microfibril angle and growth ring from pith to bark
图 3 结晶度随生长轮的径向变化规律
Figure 3 Radial variation of the crystallinity and growth ring from pith to bark
表 1 纤维形态的比较分析
Table 1. Comparative analysis of fiber morphology
树种 纤维长度 纤维宽度 双壁厚 壁腔比 变化范围/μm 平均值/μm 变异系数/% P 变化范围/μm 平均值/μm 变异系数/% P 变化范围/μm 平均值/μm 变异系数/% P 变化范围/μm 平均值/μm 变异系数/% P 纳塔栎 1 032.46~1 255.46 1 172.14±86.88 7.41 14.14~17.69 15.86 ± 1.11 7.02 8.77~11.67 10.23 ± 0.91 8.89 1.55~2.32 1.85 ± 0.26 13.97 水栎 1 018.04~1 281.27 1 178.68 ± 95.54 8.11 0.941 560 15.53~17.34 16.56 ± 0.70 4.24 0.062 224 10.65~12.15 11.19 ± 0.54 4.85 0.028 812* 1.68~2.44 2.10 ± 0.23 10.97 0.035 464* 舒玛栎 1010.34~1 292.56 1 162.45 ± 112.67 9.69 15.43~18.51 16.91 ± 1.20 7.08 舒玛栎 9.40~12.34 10.96 ± 1.03 9.37 1.67~2.15 1.91 ± 0.16 8.22 说明:P为各指标平均值的种间差异显著性分析;表示0.05水平上具有显著性差异  下载: 导出CSV
下载: 导出CSV
表 2 纤维形态、微纤丝角与生长轮的回归分析
Table 2. Regression analysis of fiber morphology, microfibril angle, and growth ring
拟合类型 样木 回归方程 R2 纤维长度与生长轮 纳塔栎 y=-2.004 3x2+48.641 0x+964.540 0 0.943 水栎 y=-4.235 4x2+76.000 0x+917.510 0 0.933 舒玛栎 y=-1.673 2x2+54.666 0x+926.210 0 0.962 纤维宽度与生长轮 纳塔栎 y=-0.004 0x3+0.040 0x2+0.330 0x+13.710 0 0.689 水栎 y=0.889 0x3—0.190 0x2+1.280 0x+14.310 0 0.656 舒玛栎 y=-0.010 0x3+0.220 0x2-0.590 0x+15.832 0 0.960 双壁厚与生长轮 纳塔栎 y=-0.120 0x2+0.400 0x+8.340 0 0.884 水栎 y=-0.116 0x3+0.790 0x2-1.630 0x+l 1.580 0 0.922 舒玛栎 y=-0.020 0x2+0.510 0x+8.780 0 0.972 微纤丝角与生长轮 纳塔栎 y=-2.529 0lnx+38.007 0 0.481 水栎 y=-0.143 9x2+1.356 4x+28.960 0 0.748 舒玛栎 y=-0.110 0x2+0.988 3x+32.896 0 0.337
下载: 导出CSV
-
[1] 刘珉.多角度解读第8次全国森林资源清查结果[J].林业经济, 2014, 36(5):3-9, 15. LIU Min. Study on the 8th national forest inventory from multiple perspective[J]. For Econ, 2014, 36(5):3-9, 15. [2] 陈益泰, 孙海菁, 王树凤, 等. 5种北美栎树在我国长三角地区的引种生长表现[J].林业科学研究, 2013, 26(3):344-351. CHEN Yitai, SUN Haijing, WANG Shufeng, et al. Growth performances of five north Ameirican oak species in Yangzi river Delta of China[J]. For Res, 2013, 26(3):344-351. [3] 兰士波.中国北方落叶栎类研究进展及前景[J].中国林副特产, 2018(4):71-76. LAN Shibo. Present research progress and prospect on deciduous oak of northern China[J]. For By-Prod Spec China, 2018(4):71-76. [4] ZHANG Bo, FEI Benhua, YU Yan, et al. Microfibril angle variability in masson pine (Pinus massoniana Lamb.) using X-ray diffraction[J]. For Studies China, 2007, 9(1):33-38. [5] 胡梦霄, 杭芸, 黄华宏, 等.杉木木材结晶度的近红外预测模型建立及变异分析[J].浙江农林大学学报, 2017, 34(2):361-368. HU Mengxiao, HANG Yun, HUANG Huahong, et al. A near infrared prediction model and variation analysis of wood crystallinity in Cunninghamia lanceolata[J]. J Zhejiang A&F Univ, 2017, 34(2):361-368. [6] 全国木材标准化技术委员会.木材物理力学试材采集方法: GB/T 1927-2009[S].北京: 中国标准出版社, 2009. [7] 郑明朝, 黄金, 韦鹏练, 等. 15年生巨尾桉纤维特性分析[J].江西农业学报, 2018, 30(9):31-34. ZHENG Mingchao, HUANG Jin, WEI Penglian, et al. Fiber characteristics of 15-year-old Eucalyptus grandis×E. urophylla[J]. Acta Agric Jiangxi, 2018, 30(9):31-34. [8] 杨振亚. 4个引种栎类树种苗期抗逆性初步研究[D].泰安: 山东农业大学, 2016. YANG Zhenya. Studies on Stress-resistance of Four Introduced Oaks Seedlings[D]. Taian: Shangdong Agricutral University, 2016. [9] 方忠艳, 廖薇, 余丽萍, 等.贵州不同种源山棕纤维结晶度的分析与比较[J].西部林业科学, 2018, 47(1):92-96, 103. FANG Zhongyan, LIAO Wei, YU Liping, et al. Comparative study on crystallinity of mountain palm fiber from different provenances of Guizhou[J]. J West China For Sci, 2018, 47(1):92-96, 103. [10] 赵西平, 张超男, 刘高均, 等.蒙古栎次生木质部解剖结构分析[J].西南林业大学学报, 2013, 33(6):76-79. ZHAO Xiping, ZHANG Chaonan, LIU Gaojun, et al. Anatomical structure analyses of secondary xylem of Quercus mongolica[J]. J Southwest For Univ, 2013, 33(6):76-79. [11] LEAL S, SOUSA V B, PEREIRA H. Within and between-tree variation in the biometry of wood rays and fibres in cork oak (Quercus suber L.)[J]. Wood Sci Technol, 2006, 40(7):585-597. [12] VOULGARIDIS E. Wood cell morphology characteristics of some oak species and mediterranean shrubs[J]. Holz als Roh-und Werkstoff, 1990, 48(7/8):261-267. [13] 梅爱君, 徐卫南, 童再康, 等.不同地区光皮桦木材纤维形态特征的研究[J].林业科技, 2017, 42(3):20-24. MEI Aijun, XU Weinan, TONG Zaikang, et al. Study on morphological properties of Betula luminifera wood fiber for different regions[J]. For Sci Technol, 2017, 42(3):20-24. [14] 刘晓玲, 符韵林.人工林观光木主要解剖特性及基本密度研究[J].浙江农林大学学报, 2013, 30(5):769-776. LIU Xiaoling, FU Yunlin. Anatomy and basic density of Tsoongiodendron odorum[J]. J Zhejiang A&F Univ, 2013, 30(5):769-776. [15] 于朝阳, 苌姗姗, 胡进波, 等.莎丽格木材解剖学特征及物理力学性能研究[J].林产工业, 2018, 45(11):17-21. YU Zhaoyang, CHANG Shanshan, HU Jinbo, et al. Study on the anatomical features and physical-mechanical properties of salicaceae wood[J]. China For Prod Ind, 2018, 45(11):17-21. [16] 周维, 卢翠香, 杨中宁, 等. 6年生大花序桉不同种源木材纤维特性的差异分析[J].西部林业科学, 2016, 45(2):29-34. ZHOU Wei, LU Cuixiang, YANG Zhongning, et al. Genetic variation in fiber characteristics of 9 provenances of Eucalyptus cloeziana[J]. J West China For Sci, 2016, 45(2):29-34. [17] LEHRINGER C, DANIEL G, SCHMITT U. TEM/FE-SEM studies on tension wood fibres of Acer spp. Fagus sylvatica L. and Quercus robur L.[J]. Wood Sci Technol, 2009, 43(7/8):691-702. [18] 杨燕, 张青.西南桦木材微纤丝角的径向变异规律[J].木材加工机械, 2010, 21(4):16-18, 24. YANG Yan, ZHANG Qing. The radial variation patterns of microfibril angle of alder birch[J]. Wood Proc Mach, 2010, 21(4):16-18, 24. [19] HADDADOU I, ALIOUCHE D, BROSSE N, et al. Characterization of cellulose prepared from some Algerian lignocellulosic materials (zeen oak wood, Aleppo pine wood and date palm rachis)[J]. Eur J Wood Prod, 2015, 73(3):419-421. [20] 廖声熙, 杨振寅, 崔凯, 等.翠柏木材管胞特性及结晶度的径向变异分析[J].南京林业大学学报(自然科学版), 2013, 37(1):87-90. LIAO Shengxi, YANG Zhenyin, CUI Kai, et al. Radical variation of wood tracheid character and crystallinity of precious Calocedrus macrolepis[J]. J Nanjing For Univ Nat Sci Ed, 2013, 37(1):87-90. [21] 郭东强, 叶露, 周维, 等. 2个种源邓恩桉木材纤维特性及变异[J].浙江农林大学学报, 2014, 31(4):502-507. GUO Dongqiang, YE Lu, ZHOU Wei, et al. Wood fiber features for two provenances of Eucalyptus dunnii[J]. J Zhejiang A&F Univ, 2014, 31(4):502-507. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2020.01.021
 点击查看大图
点击查看大图
计量
- 文章访问数: 2614
- HTML全文浏览量: 597
- PDF下载量: 40
- 被引次数: 0
