







-
竹材是一种高效再生的绿色资源,具有生长面积广、生长速度快、经济价值高等优势,是木材的优质替代品,目前已广泛应用在车厢底板、制浆造纸、工程家具等不同领域[1−2]。第九次全国森林资源清查结果表明:中国竹材年产量占世界总量的1/3,位居全球之首[3]。近20 a来竹资源的利用及人造板行业的发展,使得竹资源的利用集中在以毛竹Phyllostachys edulis为主的大径级竹材,使毛竹成为工业应用中最普遍的竹种[4−6]。然而毛竹材具有梯度特性,各个部位的纤维形态不相同,在生产中利用率低。如何优质高效的利用竹材,已成为行业内亟待解决的突出问题。深入了解毛竹的梯度结构,分析各部位纤维形态差异,对毛竹的高效利用具有重要价值。
蔡燚等[7]对3~5年生毛竹纤维形态及化学成分进行测试分析发现:毛竹的纤维形态优于一般的阔叶材,属于长纤维类,是优良的制浆造纸原料。李荣荣等[8]研究认为:毛竹不同部位的纤维形态和部分物理性能存在一定差异,毛竹竹黄、竹肉与竹青的纤维长度和宽度存在显著差异,且不同长度的纤维占比和纤维长宽比亦存在差异。夏旭光等[9]对不同部位的竹材进行力学测试表明:单层竹青与单层竹黄的弹性模量有极大差异,两者模量比值最大可达201%,最小为173%。综上所述,梯度结构决定了毛竹材各部位纤维形态及力学特性,直接影响竹纤维的生产工艺及成品性能。鉴于此,本研究选用不同竹龄的毛竹为研究对象,探究不同竹龄不同取样部位的纤维形态和结晶度差异,为毛竹材的合理高效利用提供理论依据。
-
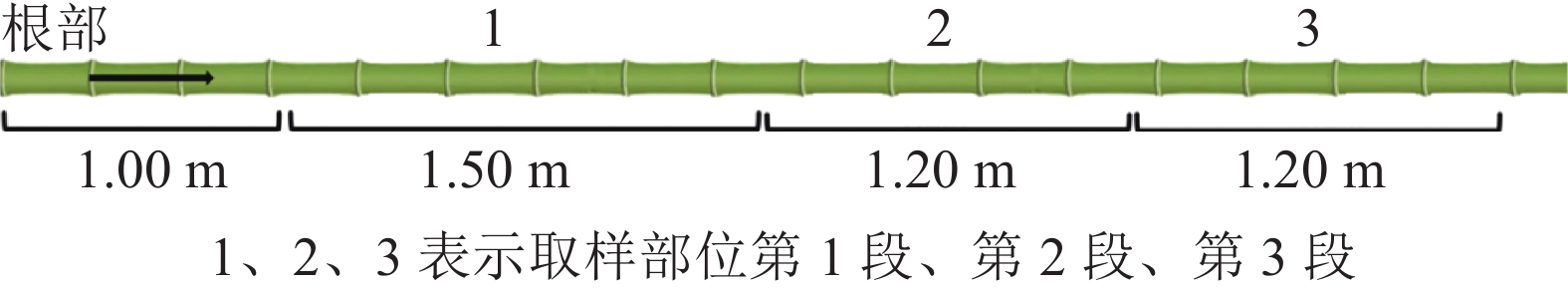
在安徽省六安市金寨县青山镇分别选取竹龄为2、4、6 a的毛竹各3株。根据GB/T 15780—1995《竹材物理力学性质试验方法》选取适合的毛竹材。如图1所示:将选好的毛竹材沿根部截断,去枝备用。在距离地约1.00 m的整竹节处向上截取1.50 m为第1段(离地1.75 m),标记为1,后每1.20 m截取第2段(离地3.25 m)、第3段(离地4.30 m),标记为2、3,去除竹节,气干至含水率为12%~15%,备用。样株取样信息见表1。
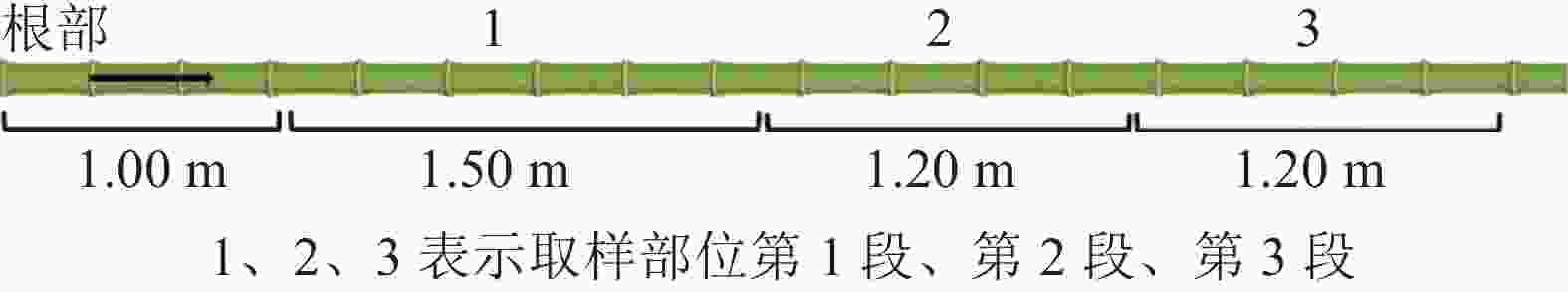
图 1 取样部位示意图
Figure 1. Schematic diagram of sampling position
表 1 采集试样的基本情况
Table 1. Basic information of sample collection
取样部位 不同竹龄取样位置/竹节 不同竹龄取样壁厚/mm 2 a 4 a 6 a 2 a 4 a 6 a 1 6~12 6~11 6~12 11.07 11.29 12.02 2 13~17 12~16 13~17 10.47 9.06 10.32 3 18~21 17~20 18~21 8.91 8.85 9.58 说明:取样部位1、2、3表示竹材由下至上不同部位,具体见图1 冰乙酸购于江苏省强盛功能化学股份有限公司,质量分数为30%的过氧化氢溶液购于上海振企精细化学品有限公司。
-
制样流程如图2所示:将气干状态的竹材用砂盘砂带机砂去除试样的竹黄和竹青部分,随后用破篾机将竹材径向均分为5等份,依次标记为A (近竹黄)、B、C、D、E (近竹青)。再将分好类的竹片切成小火柴棒状,大小为1 mm (长)×1 mm (宽)×30 mm (高)。

图 2 纤维形态测试取样流程
Figure 2. Sampling process for fiber morphology test
采用富兰克林离析法获取不同竹龄和不同部位的竹纤维。具体步骤:将不同竹龄和部位的竹片劈成火柴棒状,取制备好的试样3根放入小试管中,倒入配好的冰乙酸和过氧化氢(体积比1∶1)溶液10 mL,用水浴锅加热,温度为60~70 ℃;加热至试样呈白色后,用蒸馏水反复冲洗至小木棒上的药品完全干净,在试管中到入适量蒸馏水,用玻璃棒搅拌至纤维分离,保鲜膜封闭试管口备用。利用正置荧光显微镜测量纤维形态特征,测定毛竹材的纤维长度、宽度、腔径和壁厚,并计算纤维长宽比及壁腔比[10]。每组样品随机测量50根纤维。
-
取不同竹龄和部位的毛竹试样,研磨,筛选大于100目的粉末,置于烘箱(103±2) ℃烘至绝干备用。采用XD6多晶X射线衍射仪进行测定。测试参数:Cu-Ka靶,管电压40 kV,管电流30 mA,波长λ为1.540 566 nm,样品扫描角度为5°~40°,扫描速率为4°·min−1。通过Segal法计算相对结晶度[11]。每组样品制备15个结晶度样品,各扫描3次,取平均值。相对结晶度(Cr)计算公式为Cr=(Im−Ix)/Im×100%。其中:Im为晶体衍射角的极大强度;Ix为2
$ \theta $ =18°时非结晶背景衍射时的强度。 -
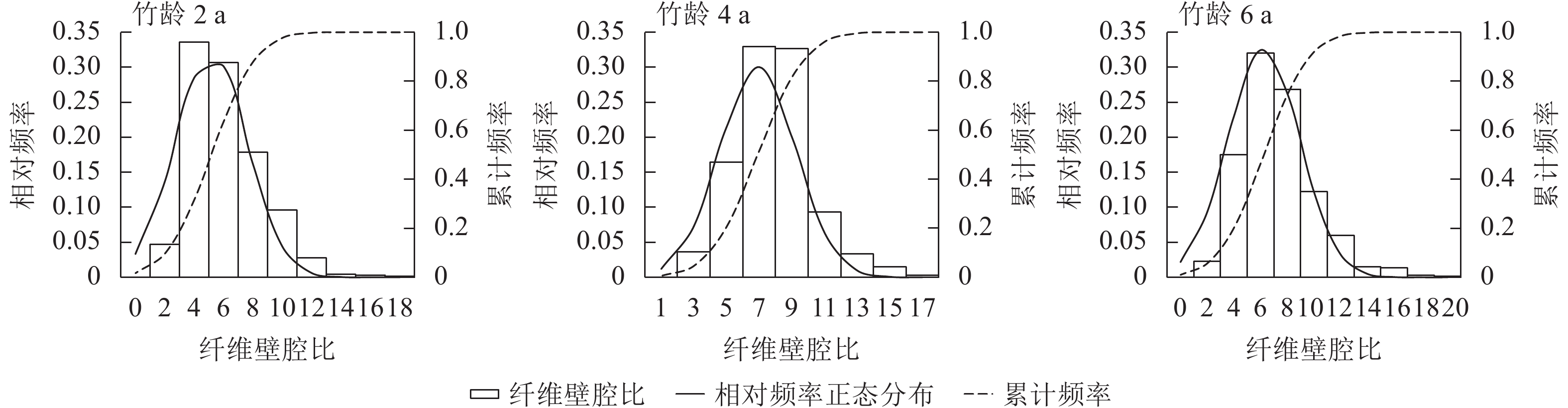
由图3可知:竹材纤维壁腔比随竹龄增长而增加,在竹龄6 a时达到峰值。竹龄2 a的毛竹82.1%纤维壁腔比集中在6.00,仅有不到0.8%的纤维壁腔比大于12.00;竹龄4和6 a的毛竹纤维壁腔比均在7.00附近达到顶峰,其中竹龄4 a的毛竹纤维壁腔比极值为16.97,而竹龄6 a的毛竹壁腔比极值为20.33。纤维细胞壁厚是影响纤维壁腔比的1个重要因素,竹纤维细胞和基本薄壁组织的细胞存在多层结构,由厚层与薄层相互交织而成,其中次生壁在细胞壁结构中占比最多[12−13]。毛竹纤维细胞通常在出笋至1 a内,壁层数量明显增加,形成3~6层次生壁;1~6 a间,壁层增加速度减缓,6 a时达到顶峰,形成6~11层次生壁;随后壁层数量随竹龄的增长逐渐回落[14]。因此,竹龄6 a的毛竹壁腔比明显大于其他年份。

图 3 不同竹龄毛竹的纤维壁腔比
Figure 3. Wall cavity ratio of Ph. edulis in different ages
纤维长度主要影响纤维的交织能力,纤维长度越长,其纠缠其他纤维的节点就越多,纤维交织能力越好,纤维间结合力越强。纤维长度是衡量纸浆造纸品质的重要指标,在一定范围内,纤维越长,纸张撕裂度、抗张强度、耐破度和耐折度就越大[15−16]。而对竹纤维水泥基材料而言,短纤维在水泥基材料中较长纤维分布更均匀,密实性高,孔隙率低,干表观密度更大,则更宜选取壁腔比小、纤维长度较短的原竹纤维[17]。纤维可以分为长度≤900 µm的短纤维,长度为900~1 600 µm的中等纤维,长度>1 600 µm的长纤维[18]。在工业生产中应选择相对应的纤维长度,以提高产品质量与材料利用率。
由图4可知:毛竹纤维长度在竹龄为2~6 a时相对稳定,随着竹龄的增长,纤维细胞缓慢发育,纤维长度呈现小幅增长。其中,竹龄2 a的毛竹纤维长度分布在1 600 µm附近最多,占比22.13%;竹龄4 和6 a 的毛竹纤维长度分别在1 700、2 000 µm达到峰值;竹龄2 a的毛竹纤维有3%的样本纤维长度小于1 000 µm,而竹龄4 和6 a 的仅为1.8%与0.8%。

图 4 不同竹龄毛竹的纤维长度
Figure 4. Fiber length of Ph. edulis in different ages
细而长的纤维能增加纸张的强度,而长宽比低于45则不适用于造纸原材料[19]。竹龄2和6 a的毛竹纤维长宽比频数峰值均在120 (图5),分布相对集中稳定,而竹龄4 a的毛竹纤维长宽比多集中于160,这表明竹龄为2~6 a的毛竹纤维均可用于制浆造纸行业。
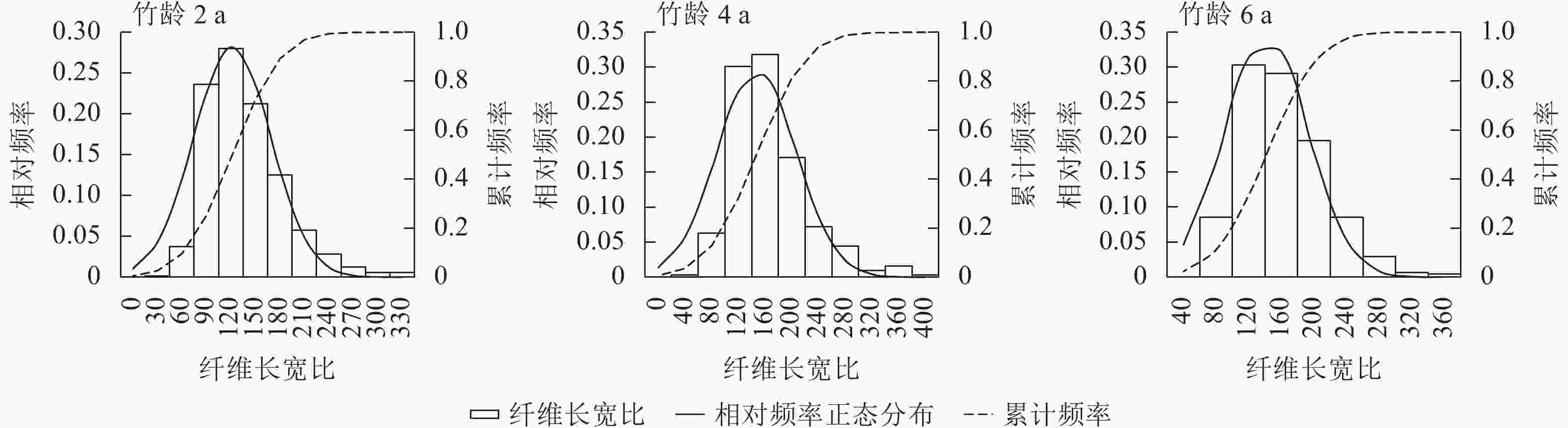
图 5 不同竹龄毛竹材的纤维长宽比
Figure 5. Fiber length width ratio of Ph. edulis in different ages
-
由表2可知:竹龄2 a毛竹的第3段纤维形态与第1段、第2段有明显差异。其中,第3段的纤维长度更长,长宽比更大,但其纤维壁腔比也随之增加,综合比较并无明显的制浆优势。竹龄4和6 a的毛竹纤维壁腔比均在第2段取样处最大,竹龄4 a毛竹的第2段纤维长度最短,竹龄6 a的毛竹第2段纤维长宽比最小,总体并未呈现明显变化规律。马灵飞等[20]研究表明:竹材纤维长宽比在轴向上并未存在显著关系,且竹龄的影响很小。蔡燚等[7]认为毛竹的纤维长宽比在轴向高度上呈现先减小后增加再减小的趋势,并在5.5 m处达最大值。以上研究与本研究在取样重合区间和测定数据等方面相符。
表 2 毛竹轴向纤维形态差异
Table 2. Differences of Ph. edulis in axial fiber morphology
取样
部位竹龄2 a 竹龄4 a 竹龄6 a 纤维壁腔比 纤维长度/µm 纤维长宽比 纤维壁腔比 纤维长度/µm 纤维长宽比 纤维壁腔比 纤维长度/µm 纤维长宽比 1 4.90±0.15 a 1 787.04±34.14 AB 114.29±2.82 a 6.30±0.16 a 1 810.21±28.98 a 133.62±2.98 A 7.91±0.18 A 1 854.00±27.22 a 146.71±3.05 a 2 4.90±0.16 a 1 754.41±29.25 A 117.58±2.99 a 7.31±0.13 b 1 769.81±33.86 a 149.43±3.83 B 8.70±0.18 B 1 884.01±29.88 a 134.47±3.00 b 3 5.69±0.14 b 1 864.93±29.80 B 133.02±3.01 b 7.11±0.14 b 1 852.12±32.62 a 158.67±3.69 B 8.10±0.14 A 1 894.12±31.17 a 143.75±3.35 ab 平均值 5.16 1 802.13 121.63 6.91 1 810.71 147.24 8.24 1 877.38 141.64 说明:数据为平均值±标准误。取样部位1、2、3所表示的具体位置见图1。小写字母表示数据符合方差齐性检验,两两比较为邦弗伦尼法;大写字母表示数据不符合方差齐性检验,两两比较为塔姆黑尼法。同列不同字母表示不同取样部位差异显著(P<0.05) -
影响纤维强度即纤维力学性能的主要因素是纤维壁腔比。纤维壁腔比越小,表示纤维细胞壁越薄,胞腔越大,纤维柔韧性越好,但过小的纤维壁腔比会导致纤维的力学性能变差,在工业加工过程中,纤维细胞易产生塌陷或弯折,降低力学强度与交织能力。
由表3可知:随着竹龄的增加,毛竹径向上各部位的纤维壁腔比均有所增加;各竹龄毛竹径向部位由A~E纤维壁腔比变化不显著。除竹龄4 a毛竹A、B的2个取样部位之间,其他竹龄相邻取样部位的纤维长度之间差异均不显著,而纤维长度在径向上的变化明显,但趋势相对缓和;竹材纤维长度在径向上存在显著变异规律(P<0.05),竹龄4和6 a毛竹均在B~D部位最大,这与王鹏程等[21]的研究结论一致。竹龄2 a毛竹的纤维长度总体呈增加趋势,在D部位处小幅下降。竹龄4和6 a竹材的各部位纤维长宽比均为130~180,符合纺织行业对于日用纤维制品选材的标准。
表 3 毛竹径向纤维形态差异
Table 3. Differences of Ph. edulis in radial fiber morphology
取样部位
(竹黄~竹青)竹龄2 a 竹龄4 a 竹龄6 a 纤维壁腔比 纤维长度/µm 纤维长宽比 纤维壁腔比 纤维长度/µm 纤维长宽比 纤维壁腔比 纤维长度/µm 纤维长宽比 A 5.07±0.18 A 1 694.74±36.84 a 123.87±3.61 ab 6.93±0.20 a 1 714.09±43.21 a 142.49±4.29 a 9.09±0.23 a 1 863.92±36.82 ab 147.69±4.27 ac B 6.49±0.19 B 1 712.21±40.33 a 112.47±3.94 a 7.02±0.18 a 1 935.47±40.85 b 153.03±4.97 a 7.63±0.19 b 1 884.41±38.23 ab 144.30±3.84 abc C 4.84±0.22 A 1 852.63±41.17 ab 129.04±3.79 b 7.11±0.21 a 1 822.00±42.29 ab 153.07±5.00 a 7.84±0.22 b 1 973.08±36.44 a 151.02±4.28 c D 4.49±0.16 A 1 824.75±39.29 ab 125.15±4.20 ab 6.81±0.16 a 1 765.08±38.28 ab 145.83±4.59 a 7.83±0.20 b 1 865.71±38.70 ab 133.26±3.57 ab E 4.93±0.19 A 1 926.29±41.19 b 117.60±3.60 ab 6.67±0.19 a 1 817.60±39.88 ab 141.76±4.11 a 8.13±0.23 a 1 799.77±38.56 b 132.32±4.09 b 说明:数据为平均值±标准误。取样部位A~E表示的具体位置见图2。小写字母表示数据符合方差齐性检验,两两比较为邦弗伦尼法;大写字母表示数据不符合方差齐性检验,两两比较为塔姆黑尼法。同列不同字母表示不同取样部位差异显著(P<0.05) 由表4可知:在影响毛竹纤维形态的不同因子中,竹龄占比最大,贡献率为34.004%。因此,在工业生产过程中建议优先考虑竹龄因素,以达到高效合理的原竹利用。
表 4 主成分各因子提取载荷平方和
Table 4. Each factor extracts the sum of squares of the load
成分 方差百分比/% 累计贡献率/% 竹龄 30.004 30.004 轴向取样部位 22.431 52.435 径向取样部位 16.757 69.192 -
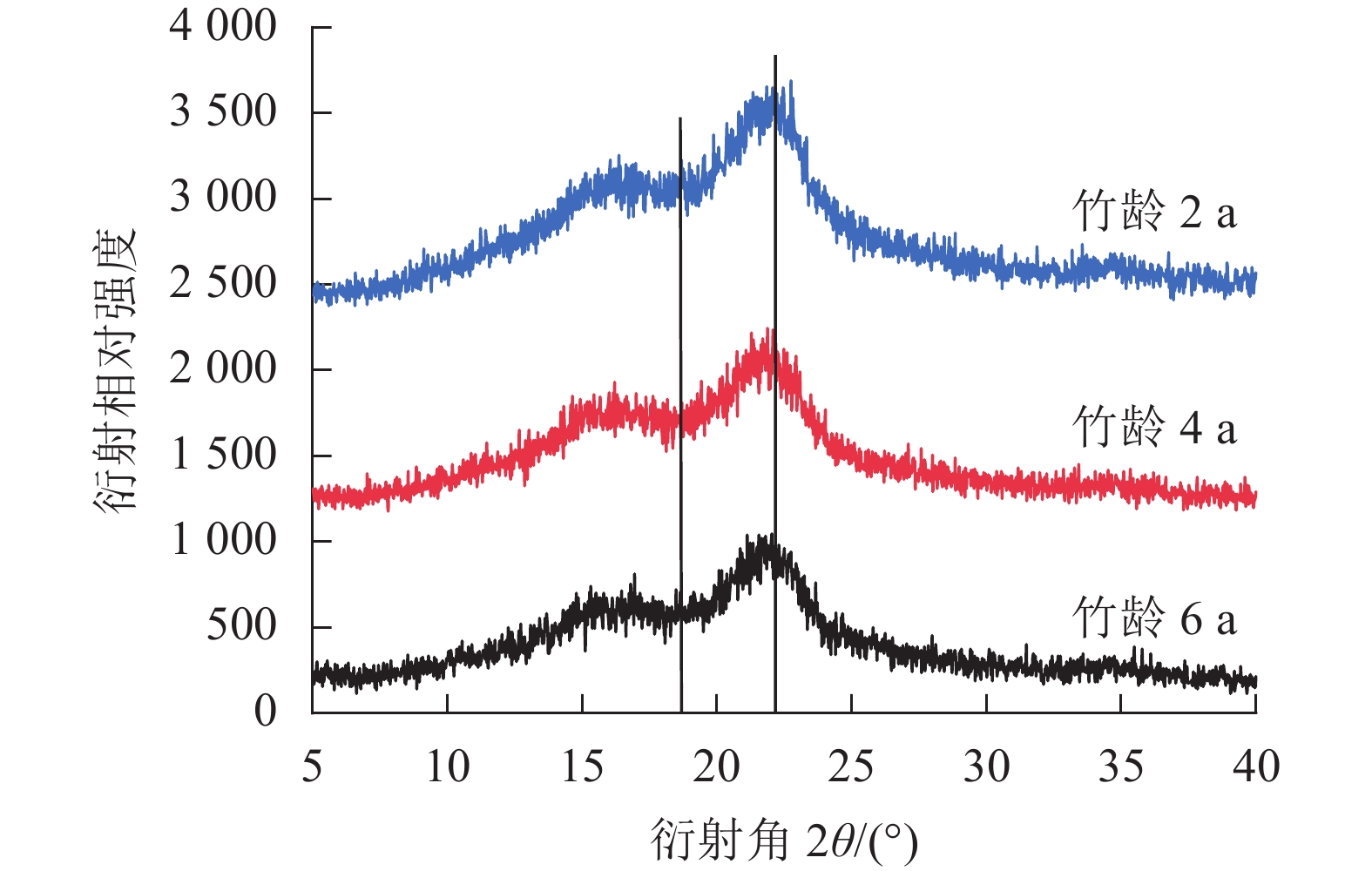
结晶度作为描述纤维素分子结构的重要参数,与木质素、纤维素和半纤维素(简称三大素)密切相关。在毛竹生长过程中结晶度的变化规律和三大素含量的变化是一致的,随竹龄增加呈现波动趋势[22]。由图6可知:不同竹龄毛竹的纤维素晶型一致,无明显差异。竹龄2、4、6 a毛竹的结晶度相似,分别为39.48%、39.72%、39.59%(表5)。
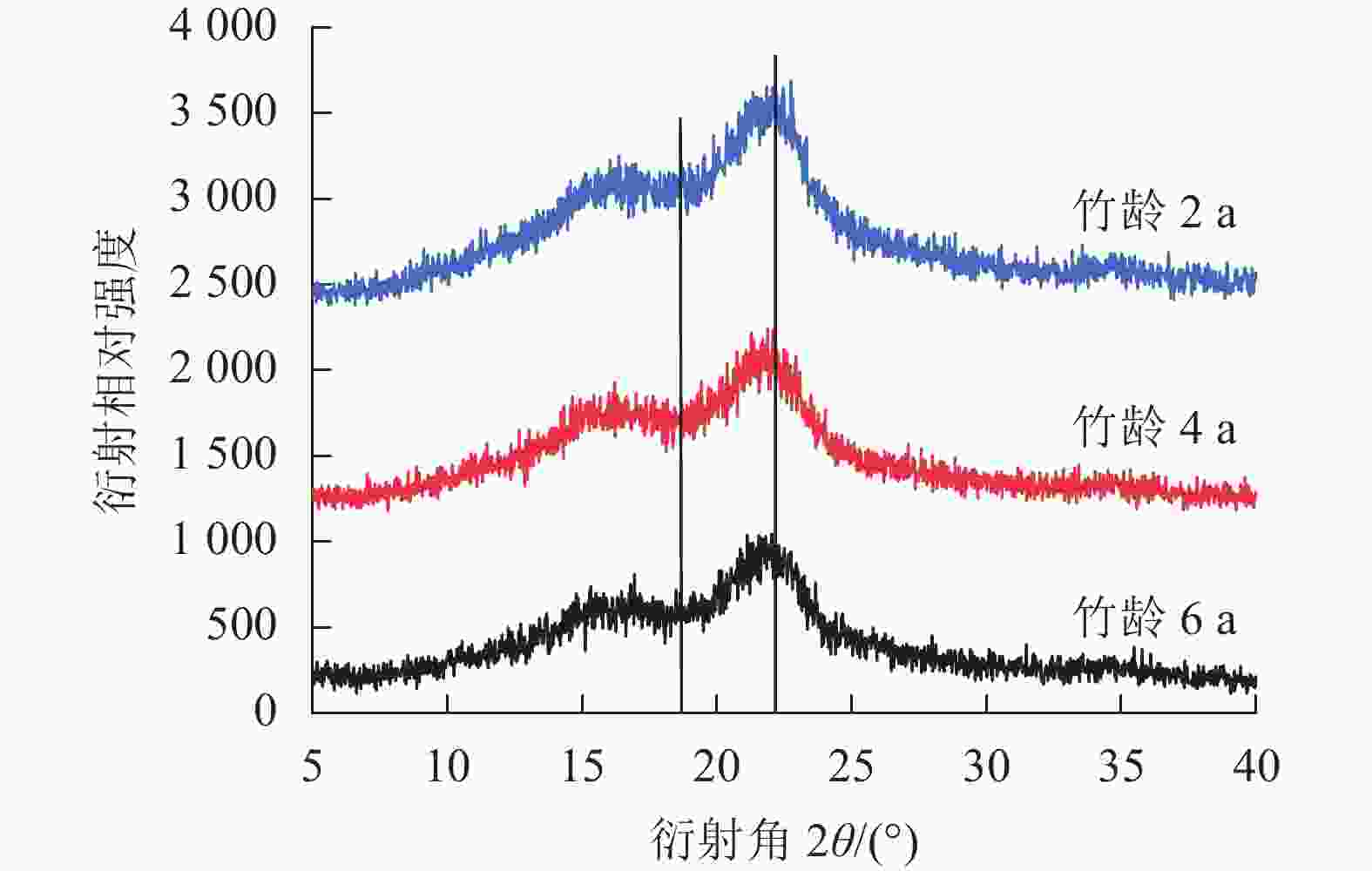
图 6 不同竹龄毛竹的X射线衍射图谱
Figure 6. X-ray diffraction patterns of Ph. edulis in different radial directions
表 5 不同竹龄毛竹轴向纤维结晶度差异
Table 5. Relative crystallinity of Ph. edulis in the axial direction under different ages
竹龄/a 不同轴向高度纤维素结晶度/% 1 2 3 平均值 2 38.75 40.23 39.46 39.48 4 38.66 39.99 40.51 39.72 6 38.52 40.52 39.68 39.59 说明:1、2、3表示取样部位,具体位置见图1 -
不同竹龄毛竹纤维结晶度在轴向上的变化规律也不相同。由表5可知:竹龄2和6 a毛竹的纤维结晶度从大到小依次为第2段、第3段、第1段,而竹龄2 a毛竹的纤维结晶度从大到小依次为第3段、第2段、第1段。
-
由表6可知:3个竹龄毛竹的径向纤维结晶度的变化均为由A到E递增。这是由于纤维细胞数量从竹黄到竹青呈增加趋势,形成纤维素结晶区的比例也随之增加;近竹黄A部位比近竹青E部位的细胞壁更薄且胞间层较多,细胞壁主要由纤维素构成,而胞间层主要由木质素构成。因此,近竹青E处比近竹黄A处有更多的纤维素和更少的木质素,故毛竹从近竹黄A到近竹青E纤维结晶度呈增大趋势。
表 6 不同竹龄毛竹径向纤维结晶度差异
Table 6. Relative crystallinity of Ph. edulis in radial direction under different ages
竹龄/a 竹壁径向位置纤维素结晶度/% A B C D E 2 36.69 38.32 38.81 40.07 42.50 4 33.47 37.89 39.46 42.91 44.88 6 35.70 37.39 39.07 42.42 42.84 说明: A~E表示取样部位,具体位置见图2 -
毛竹材纤维长度随竹龄的增加呈增大趋势,纤维形态的各项指标在轴向上无显著变化规律,径向上,4和6 a毛竹近竹黄处纤维最短,竹肉纤维最长,近竹青处纤维长度介于两者之间;纤维壁腔比和长宽比径向上变化规律不明显,竹龄对纤维结晶度的影响不显著,取样部位对纤维结晶度的影响显著。这说明在不同领域的应用过程中,竹龄是毛竹材纤维形态最显著的影响因素,在筛选原材料时,应优先考虑竹龄。
Effects of different ages and positions on fiber morphology and crystallinity of Phyllostachys edulis
-
摘要:
目的 研究竹龄与部位对毛竹Phyllostachys edulis纤维形态及结晶度的影响,为实现毛竹在制浆造纸、竹纺织品等工业生产中的高效选材利用提供基础数据。 方法 采用纤维离析法,借助普通光学显微镜,测定纤维形态;通过Segal法计算相对结晶度。 结果 竹龄主要影响竹材的纤维长度,纤维长度随竹龄的增长而增大,且80%的纤维长度为1 000~2 500 µm,属长纤维。轴向高度对毛竹材纤维形态的影响较小,纤维壁腔比、长宽比在3个取样部位间差异显著(P<0.05),但未有明显变化规律;轴向上,不同位置纤维长度未见显著差异。径向纤维长度从大到小依次为竹肉、近竹青、近竹黄;结晶度与竹龄无明显关系,径向上由近竹黄到近竹青呈现递增趋势。在影响竹材纤维形态的因子中,竹龄贡献率最大,影响最为明显。 结论 毛竹纤维形态受竹龄影响最大,受径向取样部位影响明显,轴向高度影响较小,所有部位纤维可用于工业生产,建议将竹龄作为原材料筛选的优先指标。图6表6参22 Abstract:Objective This study, with an investigation of the influence of bamboo age and position on the morphology and crystallinity of Phyllostachys edulis fiber, is aimed to provide basic data for the the reasonable selection and efficient utilization of Ph. edulis in the production of pulp, paper, bamboo textile and other industrial products. Method With fiber segregation method and common optical microscope measurement method employed, the fiber morphology was determined whereas the relative crystallinity was calculated by Segal method. Result Bamboo age was postively correlated with the fiber length of bamboo, 80% of which was between 1 000 and 2 500 µm, belonging with long fiber. The shape of bamboo fiber was less affected by the axial height of bamboo, and the fiber wall cavity ratio and length width ratio had significant differences among the three sampling positions (P<0.05), but there was no obvious change rule, and the fiber length had no significant difference. In radial direction, the fiber length displayed a significant change rule, which was generally reflected as bamboo medium>bamboo green>bamboo yellow. There was no significant relationship between crystallinity and bamboo age, and there was an increasing trend from near yellow to near green in radial direction. Among the factors affecting bamboo fiber morphology, bamboo age had the largest contribution rate and the most obvious influence. Conclusion The fiber morphology of Ph. edulis was most affected by the bamboo age, significantly affected by the radial sampling position, and less affected by the axial height. All samples could be used in industrial production, and it is recommended to take the bamboo age as the priority index for raw material screening. [Ch, 6 fig. 6 tab. 22 ref.] -
Key words:
- Phyllostachys edulis /
- fiber morphology /
- wall cavity ratio /
- fiber length /
- fiber length width ratio /
- crystallinity
-
图 6 不同竹龄毛竹的X射线衍射图谱
Figure 6 X-ray diffraction patterns of Ph. edulis in different radial directions
表 1 采集试样的基本情况
Table 1. Basic information of sample collection
取样部位 不同竹龄取样位置/竹节 不同竹龄取样壁厚/mm 2 a 4 a 6 a 2 a 4 a 6 a 1 6~12 6~11 6~12 11.07 11.29 12.02 2 13~17 12~16 13~17 10.47 9.06 10.32 3 18~21 17~20 18~21 8.91 8.85 9.58 说明:取样部位1、2、3表示竹材由下至上不同部位,具体见图1  下载: 导出CSV
下载: 导出CSV
表 2 毛竹轴向纤维形态差异
Table 2. Differences of Ph. edulis in axial fiber morphology
取样
部位竹龄2 a 竹龄4 a 竹龄6 a 纤维壁腔比 纤维长度/µm 纤维长宽比 纤维壁腔比 纤维长度/µm 纤维长宽比 纤维壁腔比 纤维长度/µm 纤维长宽比 1 4.90±0.15 a 1 787.04±34.14 AB 114.29±2.82 a 6.30±0.16 a 1 810.21±28.98 a 133.62±2.98 A 7.91±0.18 A 1 854.00±27.22 a 146.71±3.05 a 2 4.90±0.16 a 1 754.41±29.25 A 117.58±2.99 a 7.31±0.13 b 1 769.81±33.86 a 149.43±3.83 B 8.70±0.18 B 1 884.01±29.88 a 134.47±3.00 b 3 5.69±0.14 b 1 864.93±29.80 B 133.02±3.01 b 7.11±0.14 b 1 852.12±32.62 a 158.67±3.69 B 8.10±0.14 A 1 894.12±31.17 a 143.75±3.35 ab 平均值 5.16 1 802.13 121.63 6.91 1 810.71 147.24 8.24 1 877.38 141.64 说明:数据为平均值±标准误。取样部位1、2、3所表示的具体位置见图1。小写字母表示数据符合方差齐性检验,两两比较为邦弗伦尼法;大写字母表示数据不符合方差齐性检验,两两比较为塔姆黑尼法。同列不同字母表示不同取样部位差异显著(P<0.05)
下载: 导出CSV
表 3 毛竹径向纤维形态差异
Table 3. Differences of Ph. edulis in radial fiber morphology
取样部位
(竹黄~竹青)竹龄2 a 竹龄4 a 竹龄6 a 纤维壁腔比 纤维长度/µm 纤维长宽比 纤维壁腔比 纤维长度/µm 纤维长宽比 纤维壁腔比 纤维长度/µm 纤维长宽比 A 5.07±0.18 A 1 694.74±36.84 a 123.87±3.61 ab 6.93±0.20 a 1 714.09±43.21 a 142.49±4.29 a 9.09±0.23 a 1 863.92±36.82 ab 147.69±4.27 ac B 6.49±0.19 B 1 712.21±40.33 a 112.47±3.94 a 7.02±0.18 a 1 935.47±40.85 b 153.03±4.97 a 7.63±0.19 b 1 884.41±38.23 ab 144.30±3.84 abc C 4.84±0.22 A 1 852.63±41.17 ab 129.04±3.79 b 7.11±0.21 a 1 822.00±42.29 ab 153.07±5.00 a 7.84±0.22 b 1 973.08±36.44 a 151.02±4.28 c D 4.49±0.16 A 1 824.75±39.29 ab 125.15±4.20 ab 6.81±0.16 a 1 765.08±38.28 ab 145.83±4.59 a 7.83±0.20 b 1 865.71±38.70 ab 133.26±3.57 ab E 4.93±0.19 A 1 926.29±41.19 b 117.60±3.60 ab 6.67±0.19 a 1 817.60±39.88 ab 141.76±4.11 a 8.13±0.23 a 1 799.77±38.56 b 132.32±4.09 b 说明:数据为平均值±标准误。取样部位A~E表示的具体位置见图2。小写字母表示数据符合方差齐性检验,两两比较为邦弗伦尼法;大写字母表示数据不符合方差齐性检验,两两比较为塔姆黑尼法。同列不同字母表示不同取样部位差异显著(P<0.05)
下载: 导出CSV
表 4 主成分各因子提取载荷平方和
Table 4. Each factor extracts the sum of squares of the load
成分 方差百分比/% 累计贡献率/% 竹龄 30.004 30.004 轴向取样部位 22.431 52.435 径向取样部位 16.757 69.192
下载: 导出CSV
-
[1] 王晓明, 王建和. 我国竹资源综合开发利用的现状分析[J]. 浙江林学院学报, 1993, 10(1): 86 − 92. WANG Xiaoming, WANG Jianhe. Status analysis of comprehensive developing and utilization of bamboo resources in recent years in China [J]. Journal of Zhejiang Forestry College, 1993, 10(1): 86 − 92. [2] 刘娇, 周爱萍, 盛宝璐, 等. 温度对重组竹短期受压蠕变性能的影响[J]. 林业工程学报, 2021, 6(2): 64 − 69. LIU Jiao, ZHOU Aiping, SHENG Baolu, et al. Effect of temperature on short-term compression creep property of bamboo scrimber [J]. Journal of Forestry Engineering, 2021, 6(2): 64 − 69. [3] 国家林业和草原局. 中国森林资源报告(2014—2018)[M]. 北京: 中国林业出版社, 2019. State Forestry and Grassland Administration. China Forest Resources Report (2014−2018) [M]. Beijing: China Forestry Press, 2019. [4] 李媛媛, 张双燕, 王传贵, 等. 毛竹采伐剩余物的化学成分、纤维形态及纸浆性能[J]. 浙江农林大学学报, 2019, 36(2): 219 − 226. LI Yuanyuan, ZHANG Shuangyan, WANG Chuangui, et al. Chemical composition, fiber morphology, and pulping properties of logging residues in Phyllostachys edulis [J]. Journal of Zhejiang A&F University, 2019, 36(2): 219 − 226. [5] 吴杉杉, 吴婧怡. 新形势下如何更好对竹文化体育资源进行保护性开发[J]. 林产工业, 2020, 57(10): 97 − 99. WU Shanshan, WU Jingyi. How to take better protective development of bamboo culture and sports resources under the new situation [J]. Forest Products Industry, 2020, 57(10): 97 − 99. [6] WANG Xinzhou, CHENG Dali, HUANG Xianai, et al. Effect of high-temperature saturated steam treatment on the physical, chemical, and mechanical properties of Moso bamboo [J/OL]. Journal of Wood Science, 2020, 66: 52[2022-11-15]. doi: 10.1186/s10086-020-01899-8. [7] 蔡燚, 王宝金, 官洁茹, 等. 金寨毛竹纤维形态及化学成分[J]. 东北林业大学学报, 2020, 48(2): 81 − 86. CAI Yi, WANG Baojin, GUAN Jieru, et al. Fiber morphology and chemical composition of moso bamboo from Jinzhai [J]. Journal of Northeast Forestry University, 2020, 48(2): 81 − 86. [8] 李荣荣, 贺楚君, 彭博, 等. 毛竹材不同部位纤维形态及部分物理性能差异[J]. 浙江农林大学学报, 2021, 38(4): 854 − 860. LI Rongrong, HE Chujun, PENG Bo, et al. Differences in fiber morphology and partial physical properties in different parts of Phyllostachys edulis [J]. Journal of Zhejiang A&F University, 2021, 38(4): 854 − 860. [9] 夏旭光, 姚文斌, 俞伟鹏, 等. 非均匀竹材各单层顺纹抗压弹性模量的测定[J]. 竹子学报, 2018, 37(4): 49 − 55. XIA Xuguang, YAO Wenbin, YU Weipeng, et al. The measurement of the parallel grain compression MOE of each layer of non-uniform bamboo [J]. Journal of Bamboo Research, 2018, 37(4): 49 − 55. [10] 彭博, 王传贵, 张双燕. 四川两种竹材理化性质及纤维形态分析[J]. 世界竹藤通讯, 2018, 16(3): 15 − 19. PENG Bo, WANG Chuangui, ZHANG Shuangyan. Analysis of physical & chemical properties and fiber configuration of 2 species of bamboos from Sichuan Province [J]. World Bamboo and Rattan, 2018, 16(3): 15 − 19. [11] 杨淑敏, 江泽慧, 任海青, 等. 利用X-射线衍射法测定竹材纤维素结晶度[J]. 东北林业大学学报, 2010, 38(8): 75 − 77. YANG Shumin, JIANG Zehui, REN Haiqing, et al. Determination of crystallinity of bamboo cellulose by X-ray diffraction [J]. Journal of Northeast Forestry University, 2010, 38(8): 75 − 77. [12] MURPHY R J, ALVIN K L. Varation in fiber wall structure in bamboo [J]. IAWA Bulletin, 1992, 13(4): 403 − 410. [13] ITOH T. Lignification of bamboo (Phyllostachys heterocycla Mitf. ) during its growth [J]. Holzforschung, 1990, 44(3): 191 − 200. [14] 刘波. 毛竹发育过程中细胞壁形成的研究[D]. 北京: 中国林业科学研究院, 2008. LIU Bo. Formation of Cell Wall in Developmental Culms of Phyllostachys pubescens [D]. Beijing: Chinese Academy of Forestry, 2008. [15] 吴金凤, 周学政, 黄治, 等. 竹纤维润胀性能的测定方法及其应用[J]. 造纸科学及技术, 2021, 40(6): 1 − 5. WU Jinfeng, ZHOU Xuezheng, HUANG Zhi, et al. Test method of bamboo fiber swelling characteristic and its application [J]. Paper Science and Technology, 2021, 40(6): 1 − 5. [16] 杨金燕. 几种病虫害对杨树木材材质的影响[D]. 哈尔滨: 东北林业大学, 2002. YANG Jinyan. The Influence of Several Disease and Insect on Wood Quality of Populus [D]. Harbin: Northeast Forest University, 2002. [17] 王倩, 李建成. 竹纤维水泥基材料的物理力学性能研究[J]. 建材世界, 2022, 43(5): 9 − 13. WANG Qian, LI Jiancheng. Study on physical and mechanical properties of bamboo fiber cement-based materials [J]. Building Materials World, 2022, 43(5): 9 − 13. [18] 朱海龙. 美洲黑杨NL351和2-2的KP法蒸煮、氧脱木质素特性研究[D]. 南京: 南京林业大学, 2014. ZHU Hailong. Study on Delignification Properties of Populus deltoides NL351 and 2-2 in KP Pulping and Oxygen Delignification [D]. Nanjing: Nanjing Forest University, 2014. [19] 王曙光, 普晓兰, 丁雨龙, 等. 云南箭竹纤维形态变异规律[J]. 浙江林学院学报, 2009, 26(4): 528 − 532. WANG Shuguang, PU Xiaolan, DING Yulong, et al. Morphological differences of Fargesia yunnanensis fibers [J]. Journal of Zhejiang Forestry College, 2009, 26(4): 528 − 532. [20] 马灵飞, 马乃训. 毛竹材材性变异的研究[J]. 林业科学, 1997, 33(4): 356 − 364. MA Lingfei, MA Naixun. Study on the variation of wood properties of Phyllostachys pubescens [J]. Scientia Silvae Sinicae, 1997, 33(4): 356 − 364. [21] 王鹏程, 代永刚, 汪佑宏, 等. 竹龄对梁山慈竹纤维形态特征的影响[J]. 安徽农业大学学报, 2018, 45(5): 853 − 860. WANG Pengcheng, DAI Yonggang, WANG Youhong, et al. Effect of age on the fiber morphological characteristics of Dendrocalamus farinosus [J]. Journal of Anhui Agricultural University, 2018, 45(5): 853 − 860. [22] 黄建辉, 陈灵芝. 北京百花山附近杂灌丛的化学元素含量特征[J]. 植物生态学与地植物学学报, 1991, 15(3): 224 − 233. HUANG Jianhui, CHEN Lingzhi. Chemical element content characteristics of the shrubs near Baihua Mountain in Beijing [J]. Acta Phytoecologica et Geobotanica Sinica, 1991, 15(3): 224 − 233. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220749
 点击查看大图
点击查看大图
计量
- 文章访问数: 2682
- HTML全文浏览量: 493
- PDF下载量: 105
- 被引次数: 0

