




 下载:
下载:


-
果胶是植物中的一种酸性多糖物质,主要存在于植物的细胞壁和细胞内层,为内部细胞的支撑物质。果胶类多糖包括三大类:同型半乳糖醛酸聚糖(HG),鼠李糖半乳糖醛酸聚糖Ⅰ(RG-Ⅰ)和鼠李糖半乳糖醛酸聚糖Ⅱ(RG-Ⅱ)[1-2]。其中RG-Ⅱ至少有12种不同成分的糖基残基组成,而2-酮-3-脱氧辛糖酸(KDO)是RG-Ⅱ成分中很少见的八碳糖[3],其在进化过程中也很保守,对花粉管的生长和伸长具有一定的作用[4-5]。KDO的合成部位为细胞溶胶质,合成过程中涉及到5种酶[6]。脱氧辛糖酸-8-磷酸合酶(3-Deoxy-D-manno-octulosonate 8-phosphate synthase,KDO8PS,EC)是参与KDO合成代谢的关键酶之一,催化浓缩磷酸烯醇式丙酮酸与α-阿拉伯糖5磷酸产生2-酮-3-脱氧辛糖8磷酸(3-Deoxy-D-manno-octulosonate8-phosphate),并释放无机磷酸(Pi)。产物2-酮-3-脱氧辛糖-8磷酸是2-酮-3-脱氧辛糖酸的前体。最初研究发现,在所有的革兰氏阴性菌中(Gˉ),2-酮-3-脱氧辛糖酸是脂多糖组分中必需的八碳糖,它连接类脂A与O-特异侧链共同嵌入细胞壁的外膜上,其中类脂A是内毒素的物质基础,O-抗原的多样性决定了Gˉ表面抗原决定簇的多样性,脂多糖(LPS)具有控制着某些物质进出细胞的部分选择性屏障功能[7]。此后又发现,2-酮-3-脱氧辛糖酸也是高等植物细胞壁以及一些绿色藻类植物细胞壁多糖的成分[3, 8]。大肠埃希菌Escherichia coli中,KDO8PS晶体结构已经被解析,它是一个不对称同源四聚体,其中每个单体都含有(β/α)8桶状折叠[9],在拟南芥Arabidopsis thaliana中分离到2个编码KDO8PS的旁系同源基因AtkdsA1和AtkdsA2[10],相对于茎干、幼嫩的叶、成熟的叶,成熟花中AtkdsA2基因的表达量明显增高,对花粉管的生长和伸长具有一定的作用,并发现KDO8PS在水溶液中主要是以二聚体形式存在[11]。但是由于在植物中KDO8PS晶体结构尚未解析,酶功能及催化作用机制、不同生物来源KDO8PS酶的催化特性等科学问题仍需深入研究。中国是竹子大国,竹产业的发展蕴藏着巨大的经济效益,其中食用竹笋拥有丰富的营养价值,被公认为是最佳的绿色食品、经济价值高[12]。在众多竹类中,毛竹Phyllostachys edulis是中国竹类植物中分布范围最广、栽培面积最大、蓄积量最多、经济价值最高的一个材用和笋用竹种,在中国林业生产中占有非常重要的地位。本研究拟克隆毛竹脱氧辛糖酸-8-磷酸合酶基因,分析其在毛竹不同组织表达特异性,进行进化系统分析和在原核细胞中过量表达,经过亲和层析和分子筛层析纯化得到高纯度蛋白,用于培养晶体。实验结果显示:该蛋白在溶液中是以二聚体形式存在,并筛选出微晶,为解析其晶体结构和揭示其酶催化机制奠定基础。
-
克隆基因所用材料取自浙江农林大学亚热带森林培育国家重点实验室培育基地培养室内半年生毛竹Phyllostachys edulis实生苗;分别取新鲜、无病虫害的根、茎和叶,液氮速冻-80 ℃保存备用。
-
核糖核酸(RNA)提取试剂/ Trizol购自北京鼎国昌盛生物技术有限公司;AMV酶反转录试剂盒,BamHI,EcoRI,T4DNA ligase酶和pMD18-T simple vector购自宝生物工程有限公司;DNA胶回收试剂盒、质粒提取试剂盒购自上海生工生物工程有限公司;大肠埃希菌菌株DH5α,RosettaTM(DE3),BL21(DE3)和pET-28a质粒(浙江农林大学亚热带森林培育国家重点实验室培育基地改造后的质粒)载体为浙江农林大学亚热带森林培育国家重点实验室培育基地所保存。
-
取新鲜、无病虫害的根、茎和叶,液氮中迅速研磨成粉末,采用RNA提取试剂盒提取RNA,用DNaseI酶消除RNA中的DNA。分别以毛竹根、茎、叶以及总RNA为模板,利用逆转录试剂盒合成cDNA的第1条链。
-
通过美国生物技术信息中心(NCBI)数据库获得双子叶植物拟南芥,水稻Oryza sativa的KDO8PS基因序列,根据该基因的开放阅读框(ORF)区以及氨基酸序列比对获得聚合酶链式反应(PCR)引物:上游BamHI酶切位点引物A 5′-CGGGATCCATGGATGCTTCGTCC-3′和下游EcoRI酶切位点引物B 5′-GGAATTCTCATTCCTGGAATGGAGTGAGGT -3′(酶切位点用下划线显示),由上海生工生物工程技术服务有限公司合成。以毛竹实生苗总RNA反转录成cDNA为模板,利用A和B特异性引物进行扩增。PCR产物电泳分析后经DNA胶回收试剂盒回收,利用EcoHI和EcoRI限制性内酶切双酶切PCR产物,获得的目的DNA片段插入经同样酶切后的pET-28a(+)表达载体,得到1个N-末端含有6个His亲和标签的重组子。将重组质粒经热击转化到大肠埃希菌菌株DH5α感受态细胞中,经菌液PCR鉴定阳性克隆,同时提取阳性克隆质粒进行双酶切和单酶切验证,经生工生物工程有限公司测序。
-
采用RT-PCR法确定PeKDO8PS基因在不同器官中的表达量。以毛竹Actin基因作为内参,PCR反应条件:94 ℃ 5 min 1个循坏;94 ℃ 30 s, 58 ℃ 30 s, 72 ℃ 45 s,28个循环;72 ℃ 8 min,可获得150 bp左右的DNA片段。目的基因(上游引物5′-ATGGATGCTTCGTCCGTGG-3′和下游引物5′-CTAGACCAGGACCACGGAAGG-3′)经RT-PCR可获得300 bp左右的目的产物,PCR反应条件选择循环次数在线性范围内,且电泳效果好的次数作为最佳循环次数。
-
重组表达质粒分别转化表达菌株RosettaTM(DE3)和BL21(DE3),从抗卡那氯霉素养平板上挑取单克隆,接入2 mL LB液体培养基(K+)37 ℃培养,当菌体OD600达到0.7时,其中一部分加入0.4 mmol·L-1 IPTG诱导,在37 ℃摇床上诱导3 h,收集菌体。另一部分加入0.4 mmol·L-1 IPTG诱导,在20 ℃摇床上诱导14 h,收集菌体。超声(开/关5 s /6 s,总时间1 min,功率20%)破碎,SDS-PAGE检测蛋白表达及可溶性情况。
-
将收集的菌体重悬在25 mL裂解缓冲液(30 mmol·L-1 Tris-HCl pH 8.0, 200 mmol·L-1氯化钠)中,加入500 g·kg-1甘油至终质量分数50 g·kg-1,超生破碎(开/关8 s /8 s,总时间15 min,功率40%),4 ℃,17 000 r·min-1离心30 min,将上清转移至镍柱(预先用超声buffer平衡),在冰上结合0.5~1.0 h后,用Buffer A(30 mmol·L-1 Tris-HCl pH 8.0, 200 mmol·L-1氯化钠, 50 mmol·L-1咪唑)洗杂蛋白,用Bradford试剂粗略检测至无颜色变化,Buffer B(30 mmol·L-1 Tris-HCl pH 8.0, 200 mmol·L-1氯化钠, 200 mmol·L-1咪唑)洗脱目的蛋白。将洗脱的目的蛋白在10 kDa浓缩柱中浓缩至500 μL左右。分子筛层析柱Superose 12 10/300 GL用Buffer C(30 mmol·L-1 Tris-HCl pH 8.0, 200 mmol·L-1氯化钠)平衡,将浓缩的蛋白上样,根据出峰的顺序,在目标蛋白峰处每500 μL收集1管,SDS-PAGE检测每管的纯度,将纯度高的蛋白组份浓缩到10 g·L-1(紫外280 nm分光光度检测),-80 ℃保存。
-
采用悬滴法初筛晶体的生长条件。初次筛选的条件是用Emerald Biosystems公司的WizardTMⅠ,WizardTMⅡ,WizardTMⅢ和WizardTMⅣ试剂盒,共有192个筛选条件。采用24孔蛋白结晶培养板,池液350 μL,蛋白液和池液以1:1比例混合,点样,密封盖玻片,置于20 ℃进行晶体培养。晶体生长情况可以在显微镜下观察。
-
经克隆后重组测序,获得开放阅读框为876 bp的目的片段,应用Blast在线软件与已报道的基因进行同源比较,发现插入片段与脱氧辛糖酸-8-磷酸合酶基因家族NeuB超家族的保守区有较高的相似性,其中与水稻的一致性达95%,与拟南芥脱氧辛糖酸-8-磷酸合酶基因一致性达84%,初步确定为毛竹脱氧辛糖酸-8-磷酸合酶基因的序列,此基因编码291个氨基酸(图 1),第260~270位为脱氧辛糖酸-8-磷酸合酶保守区,等电点为7.7,分子量为31 656.45 Da。

图 1 毛竹PeKD08PS基因ORF区及预测的氨基酸序列
Figure 1. ORF of PeKD08PS gene and the corresponding amino acid sequence
-
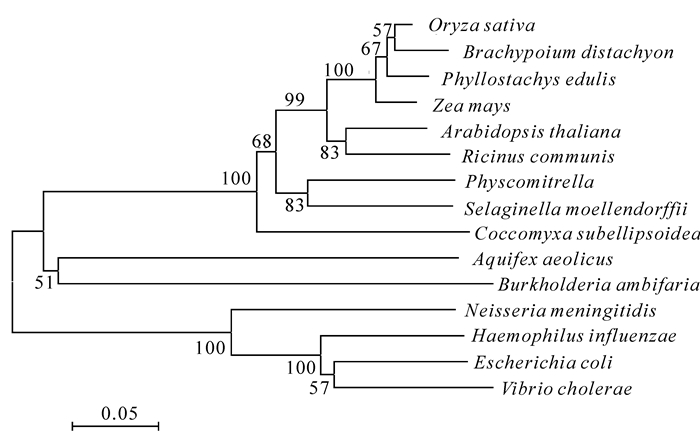
通过Blast软件在线分析,结果显示:PeKDO8PS编码的氨基酸序列与单子叶植物的KDSA具有较高的一致性,尤其与同是禾本科Poaceae的水稻\玉米Zea mays和二穗短柄草Brachypodium distachyon的一致性较高,其中与水稻的一致性达95%,与小立碗藓属Physcomitrella的一致性为76%。由此可见:KDO8PS在进化系统中是比较保守的。采用MEGA5软件构建了基于KDSA编码氨基酸序列的系统进化树(图 2)。分析表明:系统树明显分为三大支,其中竹子与水稻、玉米位置较近,位于同一个分支上;与拟南芥位于同一大分支上,与大肠埃希菌和风产杆菌分支最远。

图 2 基于不同植物KD08PS氨基酸序列构建的系统树
Figure 2. Phylogenetic tree of KDO8PS from different plants
-
分别提取毛竹根、茎和幼叶的RNA,经分光光度法检测,D(260)/D(280)为1.8~2.0,D(260)/D(280)大于2.3,表明RNA质量高,可用于进一步分析。反转录cDNA时,用毛竹actin基因作为内参,调整cDNA用量和循环数,使目的基因扩增时内参基因的表达丰度一致。半定量PCR结果表明:PeKDO8PS基因在根、茎和幼叶中均有表达,其中在根中的表达丰度最高,茎次之,在幼叶中的表达丰度最低(图 3)。

图 3 毛竹KD08PS基因在根、茎、叶中的表达
Figure 3. Tissue specific expression analysis of PeKD08PS in root. stem, and leaf
-
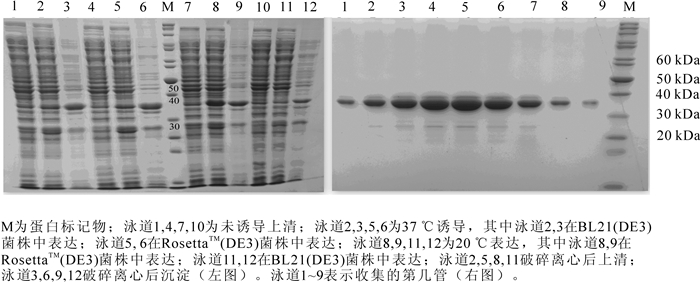
重组载体转化表达菌株RosettaTM(DE3)和BL21(DE3)感受态细胞,诱导表达后,取10 mL诱导菌体和未诱导菌体,超生破碎后,4 ℃, 17 000 r·min-1离心30 min,分别取上清、沉淀,加入上标缓冲液,95 ℃变性后,SDS-PAGE变性胶电泳分析,在电泳图谱发现1条明显的表达条带,接近理论分子量33 kDa,并且对比分析发现表达菌株RosettaTM(DE3)在20 ℃培养温度下表达量相对高些,有助于减少重组蛋白包涵体的形成,故在后续试验中确定表达菌株RosettaTM(DE3)在20 ℃为重组蛋白诱导表达的最佳条件(图 4左图)。

图 4 PeKD08PS分别在表达菌株RosettaTM(DE3)和BL21 (DE3)中的表达(左图)和PeKD08PS分子筛层析后,分管收集蛋白SDS-PAGE电泳图(右图)
Figure 4. Prokaryotic expression of PeKDOPS in RosettaTM(DE3) and BL21(DE3) cells (left) and SDS-PAGE analysis of PeKD08PS(right)
-
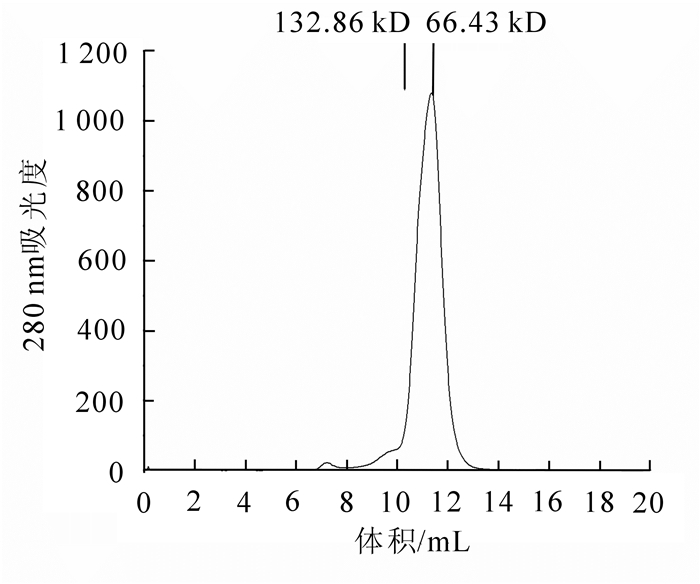
因水溶液中BSA分子之间快速可逆自聚可形成少量的BSA二聚体,所以BSA是一个很好的标记物。经Superose 12 10/300 GL层析后,对比BSA洗脱体积和分子量关系可知, 目的蛋白峰位置所对应的分子量大小在BSA单体(66.43 kDa)位置,初步确定其为二聚体(63.3 kDa,图 5),并且重组蛋白只出现明显的单峰,说明蛋白的均一性很好,收集目的蛋白,500 μL为1管,用于电泳检测和晶体生长。

图 5 PeKD08PS分子筛层析图
Figure 5. Purification of PeKD08PS by size exclusion chromatography
-
分子筛层析后分管收集的蛋白变性后用120 g·kg-1聚丙烯酰胺凝胶变性电泳检测,结果显示蛋白分子量为33 kDa(图 4右图),纯度为95%以上,可用于蛋白晶体生长。
-
收集的蛋白浓缩至30 g·L-1,可用于蛋白晶体生长。2 d后蛋白2种条件中长出了长条状晶体(图 6),2种条件分别是100 g·L-1 PEG 3000,0.1 mol·L-1 Cacodylate(pH 6.5),0.2 ·L-1氯化镁;2.5 ·L-1氯化钠, Na/K phosphate(pH 6.2)。但形状并不相同。经考马斯亮蓝试剂染色,晶体为淡蓝色,说明是蛋白晶体。优化工作正在进行中。

图 6 PeKD08PS蛋白晶体
Figure 6. Protein crystallization of PeKD08PS
-
毛竹KDO8PS编码的蛋白序列与拟南芥KDO8PS的一致性达84%,与大肠埃希菌KDO8PS一致性达49%,同属于NeuB超家族。系统进化分析显示,毛竹与水稻和玉米位置较近,位于同一个分支上;与拟南芥位于同一大分支上,与大肠埃希菌和风产杆菌Aquifex aeolicus分支最远。KDO8PS基因在植物组织中的表达具有组织特异性。本研究中,PeKDO8PS基因在根、茎和幼叶中均有表达,在根中的表达丰度较高,但在幼叶中的表达丰度最低,这与拟南芥中KDSA2表达模式相同[10],进一步推测竹子KDO8PS基因与拟南芥中KDSA2基因可能具有相似功能。本研究目前得到的KDO8PS晶体较小,结晶条件有待进一步优化。N端His标签的引入也可能增加蛋白分子柔性,不利于晶体生长[12]。后续实验将继续优化结晶条件,并通过酶法切除His标签,以提高晶体质量。
自2000年大肠埃希菌的KDO8PS结构公布后,又陆续有其他微生物的KDO8PS结构得到报道。但是植物来源KDO8PS结构至今仍未被解析,仅在拟南芥相关研究中表明KDO8PS在溶液中呈现二聚体的形式[11]。在本研究中出现一个有趣的现象:我们实验室首次获得拟南芥脱氧辛糖酸-8-磷酸合酶(KDO8PS)晶体,并解析其衍射精度2.1×10-10m的三维空间结构,其三维结构为不对称同源四聚体(文章待发表),这与大肠埃希菌的KDO8PS的不对称同源四聚体结构相似,虽与C-末端存在一定差异。但从分子筛以及戊二醛交联结果可知,拟南芥KDO8PS在溶液中以二聚体形式存在,这种情况可能是两两环抱的二体聚合,在X射线照射下呈现四聚体。而对于纯化过程中的毛竹KDO8PS,从分子筛的图形中估计也是以二聚体形式存在。因此,为了进一步了解单子叶和双子叶植物KDO8PS结构上的差异,还需获得毛竹KDO8PS较高精度的三维结构,并与拟南芥KDO8PS的三维结构进行比较。
Expression, purification, and crystallization of KDO8PS from Phyllostachys edulis
-
摘要: 3-deoxy-D-manno-octulosonate(KDO)8-phosphate synthase(KDO8PS)[EC 4.1.2.16]是KDO生物合成途径中的关键酶。采用逆转录聚合酶链式反应(RT-PCR)方法, 以毛竹Phyllostachys edulis新鲜实生苗为材料, 分离得到竹子KDO8PS基因。通过对不同物种来源的KDO8PS进行氨基酸序列比对分析和半定量RT-PCR技术对该基因进行组织特异性表达分析。将PeKDO8PS基因构建入原核表达载体pET-28a, 并在大肠埃希菌Escherichia coli原核表达系统中获得了KDO8PS的可溶性高表达。经nickel亲和层析和分子筛纯化2种方法对表达蛋白进行纯化。结果分析表明:该基因编码区全长876 bp, 可编码291个氨基酸。PeKDO8PS与拟南芥Arabidopsis thaliana等植物来源的KDSA2的基因有很高序列一致性, 而与微生物来源的KDO8PS序列一致性较低。分析毛竹各组织中KDO8PS基因的表达结果表明, 该基因在根、茎和叶片中均有表达, 但在根中相对表达量较高, 此表达模式亦同拟南芥AtKDSA2类似。另外, 发现KDO8PS在溶液(30 mmol·L-1 Tris-HCl pH 8.0, 200 mmol·L-1氯化钠)中以二聚体的形式存在, 并且初步筛选出该酶晶体生长条件。此结果为PeKDO8PS蛋白的最终结构分析奠定了基础。Abstract: 3-deoxy-D-manno-octulosonate (KDO) 8-phosphate synthase (KDO8PS)[EC 4.1.2.16] is the rate-limiting enzyme in the KDO biosynthetic pathway. In this study, the KDO8PS gene (3-deoxy-d-manno-octulosonate-8-phosphate synthase) was cloned from fresh Phyllostachys edulis seedlings using Reverse transcription polymerase chain reaction (RT-PCR). Amino acid sequence alignment analysis of the KDO-8P synthase from different organisms and tissue specific expression analysis by semi-quantitative RT-PCR were conducted. After, PeKDO8PS was recombined into the expressed pET-28a vector and over expressed in Escherichia coli. Then the fusion protein was purified through using a two-steps purification strategy including nickel affinity and size exclusion chromatography (SEC). Results showed that the open reading frame was 876 bp encoding a protein consisting of 291 amino acid residues. The PeKDO-8P synthase had high similarity with KDO-8P synthase from plants such as AtkdsA2 from Arabidopsis thaliana, while low identities from microorganism. The tissue specific expression analysis showed much higher gene expression in roots than in stems and leaves which was similar to AtkdsA2 from Arabidopsis thaliana. Finally, the chromatography analysis showed that the PeKDO8PS protein mainly existed as dimmer in 30 mmmol·L-1 Tris-HCl pH 8.0, 200 mmol·L-1 NaCl, from which the growth of preliminary screening protein crystallization condition was obtained. These works provide the first step for its structure determination.
-
图 1 毛竹PeKD08PS基因ORF区及预测的氨基酸序列
Figure 1 ORF of PeKD08PS gene and the corresponding amino acid sequence
图 2 基于不同植物KD08PS氨基酸序列构建的系统树
Figure 2 Phylogenetic tree of KDO8PS from different plants
图 3 毛竹KD08PS基因在根、茎、叶中的表达
Figure 3 Tissue specific expression analysis of PeKD08PS in root. stem, and leaf
图 4 PeKD08PS分别在表达菌株RosettaTM(DE3)和BL21 (DE3)中的表达(左图)和PeKD08PS分子筛层析后,分管收集蛋白SDS-PAGE电泳图(右图)
Figure 4 Prokaryotic expression of PeKDOPS in RosettaTM(DE3) and BL21(DE3) cells (left) and SDS-PAGE analysis of PeKD08PS(right)
-
[1] SCHOLS H A, BAKX E J, SCHIPPER D, et al. A xylogalacturonan subunit present in the modified hairy regions of apple pectin[J]. Carbohydrate Res, 1995, 279:265-279. [2] WILLATS W G T, McCARTNEY L, MACKIE W, et al. Pectin:cell biology and prospects for functional analysis[J]. Plant Mol Biol, 2001, 47:9-27. [3] YORK W S, DARVILL A G, MCNEIL M, et al. 3-deoxy-d-manno-2-octulosonic acid (KDO) is a component of rhamnogalacturonan(Ⅱ) a pectic polysaccharide in the primary cell walls of plants[J]. Carbohydrate Res, 1985, 138(1):109-126. [4] SÉVENO M, SÉVENO-CARPENTIER E, VOXEUR A, et al. Characterization of a putative 3-deoxy-D-manno-2-octulosonic acid (Kdo) transferase gene from Arabidopsis thaliana[J]. Glycobiology, 2010, 20(5):617-628. [5] LI Yi. Studies of 3-Deoxy-D-manno-Octulosonate 8-Phosphate Phosphatase:Mechanistic Insights and a Gene Fusion Example[M]. Ann Arbor:ProQuest, 2009. [6] DELMAS F, SÉVENO M, NORTHEY J G B, et al. The synthesis of the rhamnogalacturonan(Ⅱ)component 3-deoxy-D-manno-2-octulosonic acid (KDO) is required for pollen tube growth and elongation[J]. J Exp Bot, 2008, 59(10):2639-2647. [7] RAETZ C R H. Biochemistry of endotoxins[J]. Ann Rev Biochem, 1990, 59(1):129-170. [8] BECKER B, LOMMERSE J P M, MELKONIAN M, et al. The structure of an acidic trisaccharide component from a cell wall polysaccharide preparation of the green alga Tetraselmis striata Butcher[J]. Carbohydrate Res, 1995, 267(2):313-321. [9] RADAEV S, DASTIDAR P, PATEL M, et al. Structure and mechanism of 3-deoxy-D-manno-octulosonate 8-phosphate synthase[J]. J Biol Chem, 2000, 275(13):9476-9484. [10] LASKIN A I, BENNETT J W, GADD G M. Advances in Applied Microbiology[M]. San Diego:Elsevier Academic Press, 2003. [11] WU Jing, PATEL M, SUNDARAM A, et al. Functional and biochemical characterization of a recombinant Arabidopsis thaliana 3-deoxy-D-manno-octulosonate 8-phosphate synthase[J]. Biochem J, 2004, 381:185-193. [12] BERGFORS T. Seeds to crystals[J]. J Struct Biol, 2003, 142(1):66-76. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2014.04.004
 点击查看大图
点击查看大图

计量
- 文章访问数: 3818
- HTML全文浏览量: 784
- PDF下载量: 555
- 被引次数: 0
