





-
毛竹Phyllostachys edulis属于禾本科Poaceae竹亚科Bambusoideae刚竹属Phyllostachys,在中国分布范围广,被广泛应用于观赏、木材加工、食用等方面,具有很高的经济和生态价值[1]。毛竹喜湿润环境[2],干旱、盐碱、低温等非生物胁迫将会严重限制毛竹的推广与应用[3]。据报道,持续的高温少雨导致了咸宁市毛竹林大批量死亡[4],湖州市严重干旱导致大量竹笋退笋不能成竹[5],非生物胁迫已经严重影响产地竹产业及社会经济发展。目前已经展开了对毛竹逆境胁迫机理的系列研究,有利于实现毛竹可持续经营,推动竹产业经济高质量可持续发展。
bHLH (basic helix-loop-helix)转录因子在真核生物中广泛存在,主要参与生物和非生物胁迫响应[6−7]、激素信号转导途径[8]、植物的胚胎[9]、雌蕊发育[10]及开花调控[11]。在毛竹中存在153个bHLH[3]家族成员,在拟南芥Arabidopsis thaliana和水稻Oryza sativa中分别有162个[12]和167个[13],是植物中仅次于MYB的第二大基因家族[14−16]。HEIM 等[17]根据基因结构,又将拟南芥bHLH家族划分为 12 个亚家族。大多数bHLH转录因子在植物非生物胁迫信号通路中起着重要的调控作用。FAN等[18]分析了高粱Sorghum bicolor在胁迫处理下bHLH家族的表达模式,鉴定出12个可能和非生物胁迫相关的SbbHLH基因。本研究通过对毛竹实生苗进行非生物胁迫处理,利用转录组数据筛选出1条差异表达基因,命名为PhebHLH6,并对其进行生物信息学分析及表达模式探究,初步讨论了其潜在基因功能,以期为毛竹抗逆基因功能研究提供一定的理论基础。
-
毛竹26个组织转录组数据来自美国国家生物技术信息中心 (NCBI)数据库(GSE169067)[19]。毛竹种子采自广西桂林,在浙江农林大学玻璃温室(温度为26 ℃;湿度为80%)培养1个月,采集幼嫩叶片置于−80 ℃保存,用于后续研究。对生长至2月龄的毛竹实生苗进行处理,设置对照(ck)组观察取样,并在处理后的0、3、24 h分别取样保存至−80 ℃液氮。
-
对筛选得到的差异表达基因PhebHLH6进行克隆及表达模式分析。使用Oligo 7软件[20]设计蛋白编码区(CDS)全长引物,引物由杭州有康生物科技有限公司合成,引物序列参见表1。提取毛竹幼叶RNA并反转录为cDNA作为模板,参考表1中引物进行PCR扩增。反应体系为50.0 μL体系:2×Mix为25.0 μL,10.0 μmol·L−1上下游引物各1.0 μL,cDNA为1.0 μL ,ddH2O为22.0 μL。PCR反应程序:95 ℃预变性 5 min,95 ℃变性30 s,60 ℃退火30 s,72 ℃延伸60 s,34个循环,72 ℃延伸5 min。将回收产物连接pMD18-T克隆载体并转化大肠埃希菌Escherichia coli DH5α,菌检选取阳性克隆送杭州有康生物技术有限公司测序。
表 1 基因克隆及表达所用引物序列
Table 1. Primers used in gene clone and quantitative real-time PCR
用途 引物名称 引物序列(5′→3′) 基因克隆 PhebHLH6-F ATGGACGCGGACATGGGCGACA PhebHLH6-R CTAATAGCTCATCGAGCTCGGG
GGGCTTC实时荧光定量
PCR (RT-qPCR)Q-PhebHLH6-F CGAGAAGCTATACGCGATCC Q-PhebHLH6-F CTGCAGCTGCTGGATGTAAT Q-NTB-F TCTTGTTTGACACCGAAGAGGAG Q-NTB-F AATAGCTGTCCCTGGAGGAGTTT -
将克隆获取的PhebHLH6蛋白序列,利用ExPASy的在线软件Protparam (
http://web.expasy.org/protparam/ )和ProtScale (https://web.expasy.org/protscale/ )分别分析氨基酸理化性质和蛋白的亲/疏水性。分别利用PRABI的SOPMA (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.Html )和Swiss-Model (http://swissmodel. expasy.org/interactive )进行蛋白二级、三级结构分析。蛋白保守结构域使用NCBI Conserved Domain Database (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi )进行分析。根据拟南芥、水稻、玉米Zea mays、二穗短柄草Brachypodium distachyon、芸香竹Bonia amplexicaulis、瓜多竹Guadua angustifolia、Olyra latifolia和Raddia guianensis [21]蛋白数据库,进行BLAST序列比对,进一步比对得到PhebHLH6的同源基因氨基酸序列。利用MEGA11.0软件Neighbor-Joining算法,自检举1 000次,构建系统进化树。 -
使用TBtools[22]软件提取基因5′UTR上游2 000 bp序列,并使用 PlantCARE在线软件对其进行分析,预测启动子核心元件和基因结合位点。
-
参考毛竹26个组织转录组数据,使用转录数据中的FPKM值并利用TBtools软件绘制热图,对PhebHLH6基因进行不同组织表达量分析。
-
毛竹种子采自广西桂林。将发芽的种子置于温度为(25±2) ℃、相对湿度为(70±10)%的玻璃温室中,用1/4霍格兰营养液水培,每周更换1次,培养至2月龄。分别设置对照、氯化钠(NaCl)、聚乙二醇(PEG)、水杨酸(SA)、脱落酸(ABA) 5个处理。分别使用200 mmol·L−1 NaCl和质量分数为25% 的PEG营养液进行盐和干旱胁迫。此外,分别采用1 mmol·L−1和1 μmol·L−1的SA和ABA营养液进行处理。在处理0、3、24 h分别采集毛竹叶片,每个时间点采集3个生物学重复,液氮速冻,保存至−80 ℃冰箱以便后续分析使用。
使用Primer 3 version 0.4.0设计定量引物,将其在NCBI (Primer designing tool)进行引物特异性分析。实时荧光PCR (RT-qPCR)反应体系如下:cDNA(已稀释)为1.0 μL,上、下游引物各0.5 μL (10.0 μmol·L−1),NovoStart®SYBR qPCR SuperMix plus (Novoprotein)为5.0 μL,ddH2O为3.0 μL。反应程序为:95 ℃ 5 min;95 ℃ 5 s, 60 ℃ 30 s, 72 ℃ 60 s,30 循环;72 ℃ 10 min。39个循环,3次重复。数据分析采用$2^{-\Delta\Delta Ct}$方法[23]。仪器为CFX96TM Real-Time PCR Detection System (Bio-Rad)。内参基因为NTB基因[24]。数据结果采用TBtools软件绘制热图分析,以10为底取对数值。
-
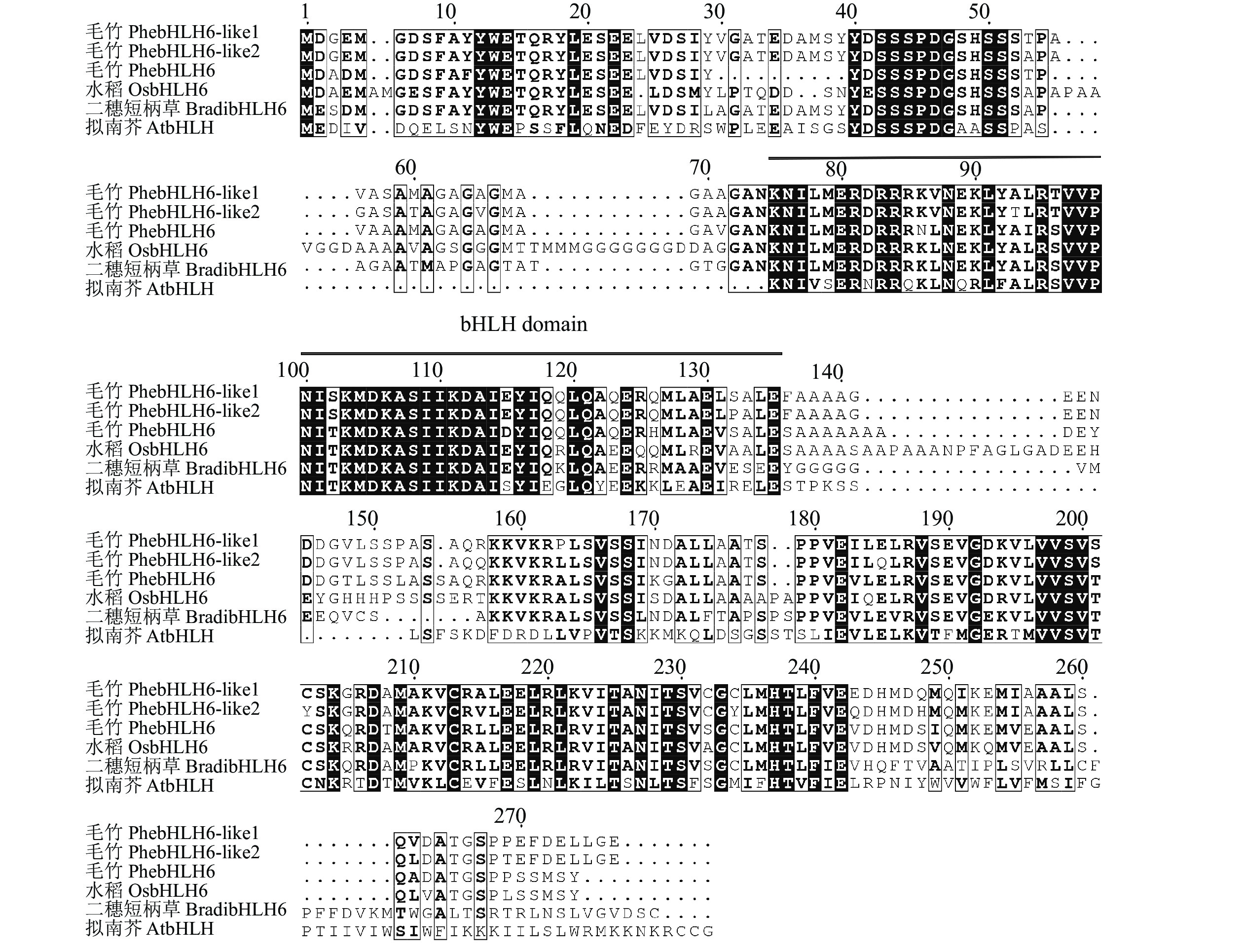
利用特异性引物进行PCR扩增,获得PhebHLH6编码氨基酸序列如图1所示。测序结果显示:编码区序列长801 bp,编码266个氨基酸。保守结构域分析结果表明:PhebHLH6蛋白在64~125氨基酸位置具有完整的bHLH保守结构域,说明该基因是典型的bHLH家族成员。

图 1 不同物种bHLH6基因氨基酸序列比对
Figure 1. Alignment of bHLH6 amino acid sequences from different species
-
蛋白序列分析结果表明:PhebHLH6的蛋白质分子量为28.651 kDa,等电点为5.04,平均疏水性为−0.102,蛋白质不稳定指数55.58,脂肪指数为88.83。上述结果表明:PhebHLH6蛋白为亲水性不稳定蛋白。二级蛋白结构预测结果显示:PhebHLH6的蛋白二级结构占比分别为α-螺旋53.76%,β-转角2.63%,延伸链9.02%,无规则卷曲结构34.59%。使用SWISS-MODEL在线比对蛋白序列,预测PhebHLH6蛋白的三级结构并建立空间模型,并与水稻OsbHLH6比较分析,发现其三级结构相似,功能可能较为保守。
-
顺式作用元件分析结果表明(表2):PhebHLH6启动子上游2 000 bp除包含TATA-box和CAAT-box等核心启动元件外,还包含ABRE元件(响应ABA)、TGACG-motif [响应茉莉酸甲酯(MEJA)]、CGTCA-motif (响应MEJA)、TGA-element (响应生长素)、AAGAA-motif (响应ABA)等一系列非生物胁迫及激素响应元件。上述结果表明:PhebHLH6可能参与到了多种非生物胁迫及激素响应途径中。
表 2 PhebHLH6基因启动子顺式作用元件分析
Table 2. Cis-element analysis of PhebHLH6 gene promoter
作用元件 序列 数量 功能 作用元件 序列 数量 功能 ABRE CACGTG 9 脱落酸响应元件 Sp1 GGGCGG 1 光响应元件 ARE AAACCA 1 厌氧诱导顺式作用元件 chs-CMA2a TCACTTGA 1 光响应元件 CAAT-box CCAAT 15 启动子和增强子区域调控元件 TGA-element AACGAC 2 生长素响应元件 CGTCA-motif CGTCA 5 茉莉酸甲酯响应元件 O2-site GATGATGTGG 1 玉米醇溶蛋白代谢调节元件 MRE AACCTAA 1 光响应元件 TATA-box TATA 35 核心启动子元件 G-Box TACGTG 3 光响应元件 TGACG-motif TGACG 5 茉莉酸甲酯响应元件 -
蛋白比对结果显示:PhebHLH6与毛竹PhebHLH6-like基因及水稻、拟南芥等物种同源基因的氨基酸序列相似,均具有完整的bHLH结构域,推测其可能拥有相似的基因功能。为进一步探究PhebHLH6功能,本研究利用毛竹、水稻、拟南芥等9个物种的bHLH6同源蛋白序列构建生物进化树。如图2所示:毛竹PhebHLH6与其他2种木本竹同源基因的蛋白聚在一起,2种草本竹聚在一起,推测木本竹和草本竹在进化过程中功能出现了一定分化。此外,竹子中的bHLH蛋白与水稻、玉米的蛋白以较高的置信度聚在一起,说明bHLH基因序列在进化方面较为保守,其功能可能具有一定相似性。
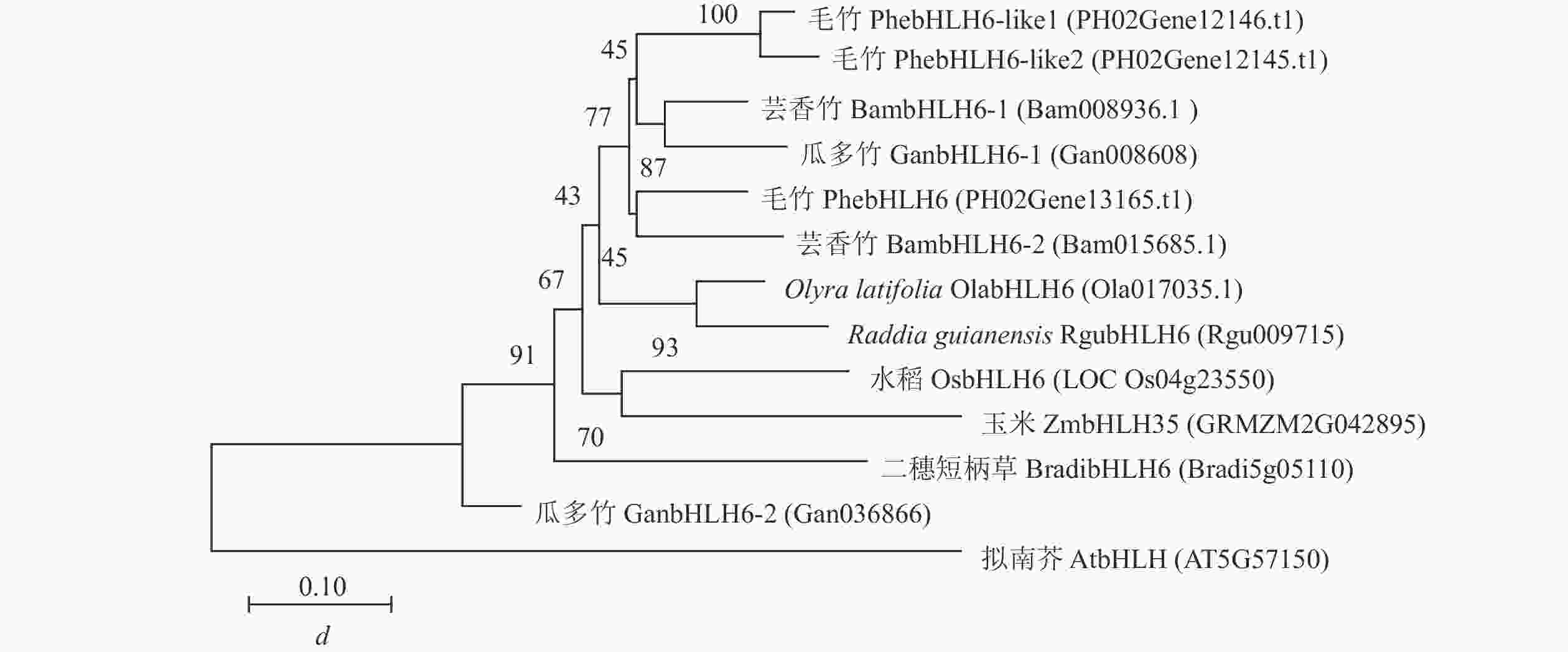
图 2 不同物种bHLH6-like氨基酸序列进化树构建
Figure 2. Phylogenetic tree construction of BHLH6-like amino acid sequences
-
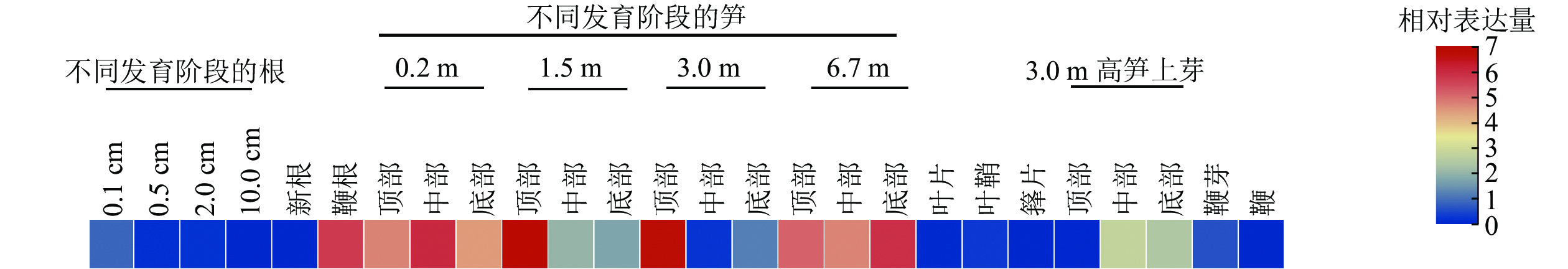
分析了PhebHLH6在毛竹26个组织中的表达水平(图3)。该基因在毛竹不同发育阶段的根、叶片、芽和鞭中相对表达量都很低,在笋中相对表达量相对较高,其中1.5和3.0 m的笋顶部相对表达量最高,6.7 m的笋各部位次之。此外,在鞭根和3.0 m高笋的侧芽中下部,PhebHLH6也有一定的表达。
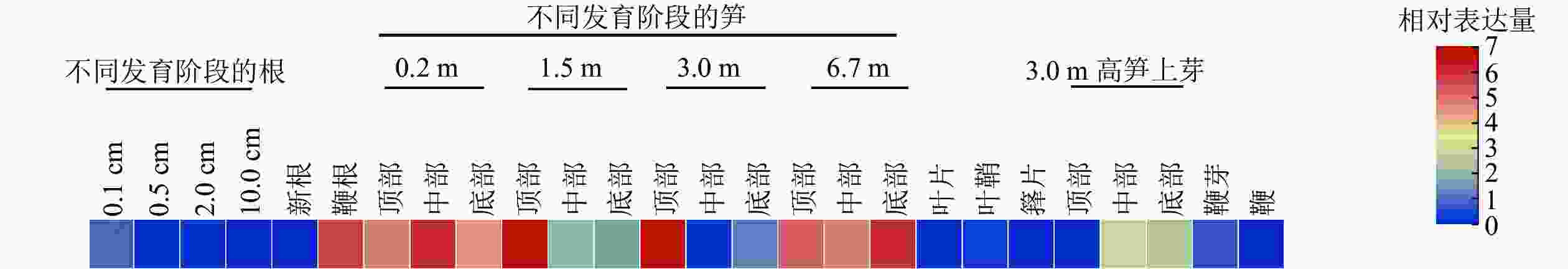
图 3 毛竹不同组织中PhebHLH6基因的表达
Figure 3. Expression level of PhebHLH6 gene in different tissues
-
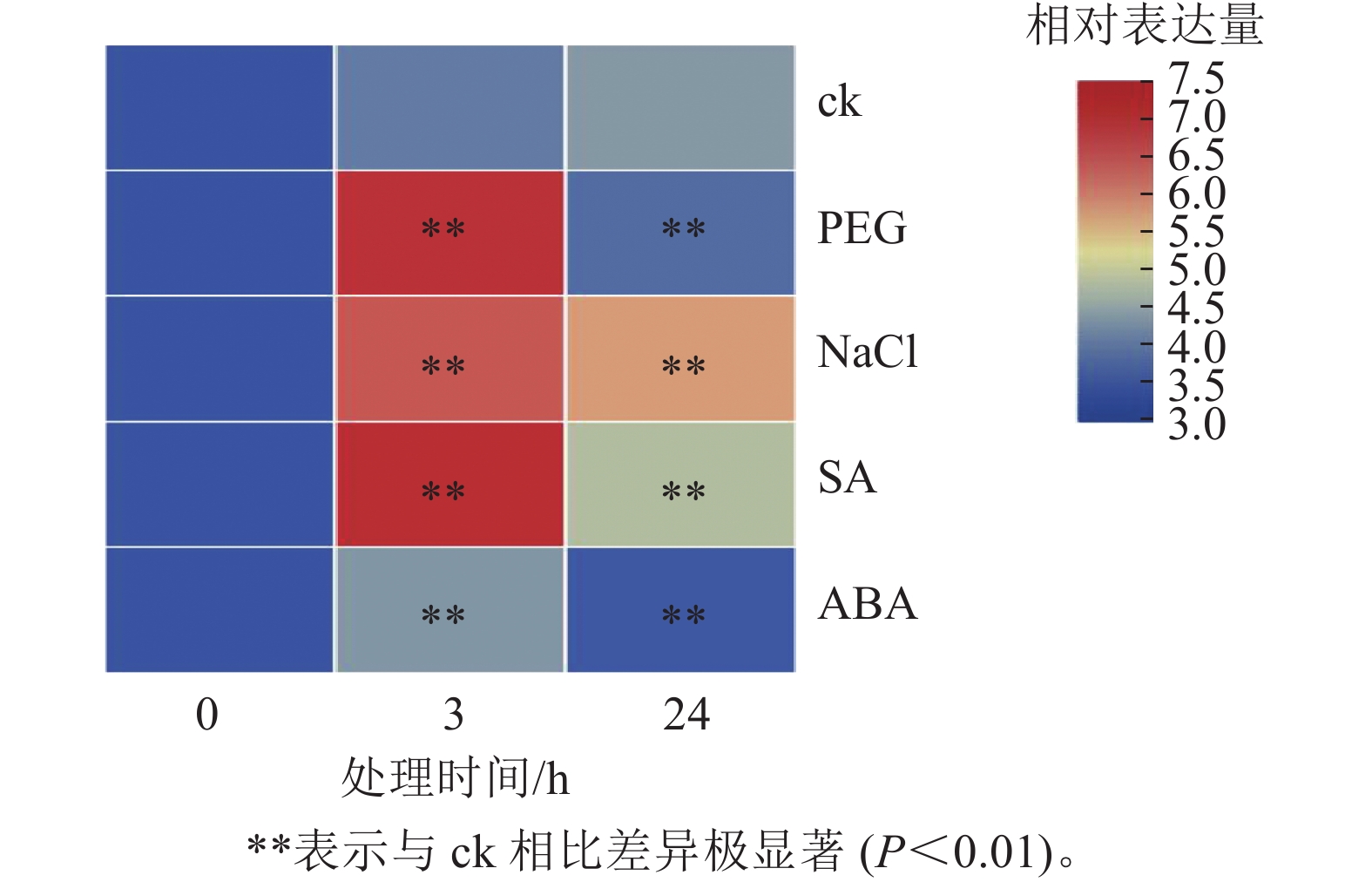
为探究PhebHLH6在非生物胁迫及激素处理下的表达模式,对2月龄毛竹实生苗进行了干旱胁迫、盐胁迫、SA和ABA处理,并通过荧光定量的方式对该基因相对表达量进行了检测(图4)。PhebHLH6基因对干旱和盐胁迫均具有强烈的响应,对SA和ABA也有一定的应答作用。
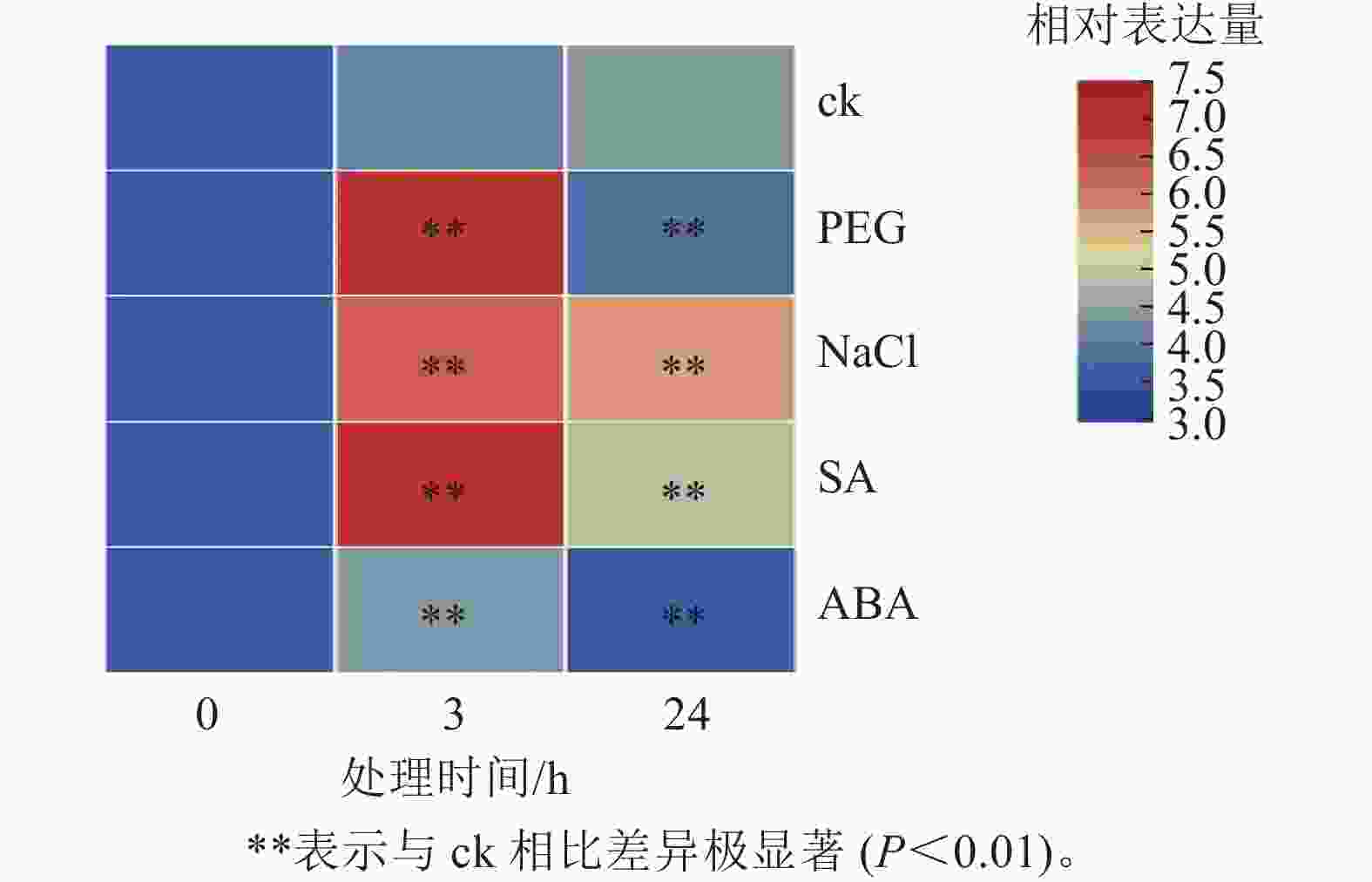
图 4 非生物胁迫以及激素处理下PhebHLH6表达模式
Figure 4. Expression patterns of PhebHLH6 under abiotic stress and hormone treatment
-
如图4所示:干旱胁迫下,与ck相比,PhebHLH6的相对表达量呈先上升后下降趋势,处理3 h后相对表达量为未处理的20倍左右,24 h后相对表达量低于ck,说明PhebHLH6在处理早期受到强烈诱导,处理后期诱导信号减弱。高盐处理下,PhebHLH6在处理3 h后相对表达量明显上升,在处理24 h后相对表达量有一定下降,但与ck相比,相对表达量仍有极显著上调(P<0.01),说明PhebHLH6持续被高盐诱导,但在处理早期诱导程度高于后期。
-
与ck相比,PhebHLH6 在SA处理3 h后相对表达量升高20倍,呈现极显著上调趋势(图4,P<0.01)。在处理24 h后,相对表达量呈下降趋势,说明PhebHLH6在处理后24 h期间持续被SA诱导,但诱导作用主要发生于早期,在处理24 h后诱导作用有所下降。ABA处理下,PhebHLH6相对表达量前期有微弱上调,24 h相对表达量呈下降趋势,说明ABA在处理前期对PhebHLH6诱导作用不强。
-
有研究指出:bHLH家族成员主要作用于植物的信号转导、生长发育及响应生物及非生物胁迫等重要途径[25]。竹子与水稻同为禾本科植物,进化关系较近,毛竹中很多基因功能研究往往参考水稻展开[1]。通过同源序列比对结果可知:PhebHLH6与水稻同源基因蛋白序列相似度为72.35%,序列相似度很高,其功能可能具有相似性。HE等[26]研究发现:水稻OsbHLH6在嫩芽和根中特异性表达,其过表达植株在低磷情况下比野生型拥有更长根系。PhebHLH6在毛竹不同组织中的表达水平分析结果表明:该基因在1.5和3.0 m的笋顶端表达量最高,推测该基因可能与毛竹笋芽顶端生长发育相关。
bHLH家族成员在植物响应逆境胁迫中也发挥重要作用。如张子佳等[27]对水稻bHLH家族响应环境胁迫表达谱分析发现:大部分家族成员响应干旱胁迫和参与ABA调控途径。水稻中OsbHLH148可以通过形成复合体的形式调控水稻抗干旱能力[28]。 OsbHLH6 (RERJ1)通过在细胞核与细胞质间的移动,动态调节水杨酸和茉莉酸(JA)激素信号来控制[29]。OsJAZ9作为bHLH家族的转录因子,通过与OsNINJA和OsbHLH蛋白形成转录调控复合物控制JA合成通路来调控植株的耐盐性[30]。OsbHLH6基因在2004年首次被鉴定为JA响应基因[31],且研究发现该基因受外界伤害和干旱胁迫诱导表达上调[32]。与之相同的是,毛竹PhebHLH6也会受到干旱和盐胁迫的强烈诱导。此外,文献报道OsbHLH6可以通过动态调节SA和JA信号通路来控制抗病能力[28]。本研究表明:毛竹PhebHLH6受SA强烈诱导,在处理后表达量极显著上升,与水稻中同源基因OsbHLH6表达模式一致,推测其可能具有功能相似性,但这些推论还需进一步验证。
-
本研究从毛竹中比对克隆得到1条bHLH同源基因,命名为PhebHLH6。PhebHLH6基因编码区长度为801 bp,编码266个氨基酸,序列包含完整的bHLH结构域,属于bHLH转录因子。组织特异性表达水平表明:该基因主要在毛竹1.5和3.0 m笋顶端表达,推测其可能和毛竹生长发育相关。此外,PhebHLH6对非生物胁迫和激素处理均有响应,其中受干旱、高盐胁迫以及SA激素处理强烈诱导,受ABA轻微诱导,表明PhebHLH6可能在植物逆境响应和激素初期信号转导中发挥着重要的作用。
Cloning and expression analysis of PhebHLH6 gene from Phyllostachys edulis
-
摘要:
目的 研究PhebHLH6转录因子在毛竹Phyllostachys edulis逆境胁迫应答中的作用,为毛竹抗逆分子机制研究奠定一定的基础。 方法 以毛竹实生苗为材料进行非生物胁迫处理[干旱胁迫、盐胁迫、水杨酸(SA)和脱落酸(ABA)处理],利用转录组数据筛选出1条差异表达基因,命名为PhebHLH6,并对其进行了基因克隆及生物信息学分析;采用实时荧光定量PCR方法分析PhebHLH6在干旱、盐胁迫及SA、ABA处理下的表达模式。 结果 PhebHLH6基因编码区长度为801 bp,编码266个氨基酸,包含bHLH结构域,属于典型的bHLH转录因子。组织特异性表达分析表明:PhebHLH6在毛竹各个组织均有表达,其中在1.5和3.0 m的笋顶部表达丰度最高。在干旱和高盐胁迫处理下,PhebHLH6的表达水平在处理3 h时被强烈诱导,但在处理24 h后显著下调。在SA和ABA处理下,PhebHLH6的表达水平被SA和ABA 诱导也呈先上升再下降的趋势,其中受SA强烈诱导,受ABA诱导作用较弱。 结论 PhebHLH6可能参与了毛竹干旱和盐胁迫早期响应途径,并可能在SA和ABA信号通路中起一定的调控作用。图4表2参32 Abstract:Objective The objective is to study the role of PhebHLH6 transcription factor in stress response of Phyllostachys edulis; so as to explore the molecular mechanism of resistance to stress in Ph. edulis. Methods Seedlings of Ph. edulis were treated with abiotic stress, including treatments of drought stress, salt stress, salicylic acid (SA) and abscisic acid (ABA). A differentially expressed gene named PhebHLH6 was screened using transcriptome data, and its gene cloning and bioinformatic analysis were performed. Real-time fluorescent quantitative PCR was used to analyze the expression patterns of PhebHLH6 under drought, salt stress and SA and ABA treatments. Result The coding region of PhebHLH6 gene had a base of 801 bp, encoding 266 amino acids, including bHLH domain, which was a typical bHLH transcription factor. Tissue-specific expression analysis showed that PhebHLH6 was expressed in nearly all the tissues of Ph. edulis, with the highest abundance at the top of 1.5 m and 3.0 m shoots. Under drought and high salt stress, the expression levels of PhebHLH6 were strongly induced after 3 h of treatment but significantly down-regulated after 24 h of treatment. Under SA and ABA hormone treatment, the expression levels of PhebHLH6 increased first and then decreased when induced by SA and ABA, with strong SA induction and weak ABA induction. Conclusion PhebHLH6 may be involved in the early response pathway to drought and salt stress of Ph. edulis and may play a regulatory role in SA and ABA hormone signaling pathways. [Ch, 4 fig. 2 tab. 32 ref.] -
Key words:
- Phyllostachys edulis /
- PhebHLH6 /
- gene clone /
- expression analysis
-
图 1 不同物种bHLH6基因氨基酸序列比对
Figure 1 Alignment of bHLH6 amino acid sequences from different species
图 2 不同物种bHLH6-like氨基酸序列进化树构建
Figure 2 Phylogenetic tree construction of BHLH6-like amino acid sequences
图 3 毛竹不同组织中PhebHLH6基因的表达
Figure 3 Expression level of PhebHLH6 gene in different tissues
图 4 非生物胁迫以及激素处理下PhebHLH6表达模式
Figure 4 Expression patterns of PhebHLH6 under abiotic stress and hormone treatment
表 1 基因克隆及表达所用引物序列
Table 1. Primers used in gene clone and quantitative real-time PCR
用途 引物名称 引物序列(5′→3′) 基因克隆 PhebHLH6-F ATGGACGCGGACATGGGCGACA PhebHLH6-R CTAATAGCTCATCGAGCTCGGG
GGGCTTC实时荧光定量
PCR (RT-qPCR)Q-PhebHLH6-F CGAGAAGCTATACGCGATCC Q-PhebHLH6-F CTGCAGCTGCTGGATGTAAT Q-NTB-F TCTTGTTTGACACCGAAGAGGAG Q-NTB-F AATAGCTGTCCCTGGAGGAGTTT  下载: 导出CSV
下载: 导出CSV
表 2 PhebHLH6基因启动子顺式作用元件分析
Table 2. Cis-element analysis of PhebHLH6 gene promoter
作用元件 序列 数量 功能 作用元件 序列 数量 功能 ABRE CACGTG 9 脱落酸响应元件 Sp1 GGGCGG 1 光响应元件 ARE AAACCA 1 厌氧诱导顺式作用元件 chs-CMA2a TCACTTGA 1 光响应元件 CAAT-box CCAAT 15 启动子和增强子区域调控元件 TGA-element AACGAC 2 生长素响应元件 CGTCA-motif CGTCA 5 茉莉酸甲酯响应元件 O2-site GATGATGTGG 1 玉米醇溶蛋白代谢调节元件 MRE AACCTAA 1 光响应元件 TATA-box TATA 35 核心启动子元件 G-Box TACGTG 3 光响应元件 TGACG-motif TGACG 5 茉莉酸甲酯响应元件
下载: 导出CSV
-
[1] PENG Zhenhua, LU Ying, LI Lubin, et al. The draft genome of the fast-growing non-timber forest species moso bamboo (Phyllostachys heterocycla) [J]. Nature Genetics, 2013, 45(4): 456 − 461. [2] 胡智勇. 毛竹的生物学特性及栽植技术[J]. 安徽农学通报, 2014, 20(12): 117 − 118. HU Zhiyong. Biological characteristics and planting techniques of Phyllostachys edulis [J]. Anhui Agricultural Science Bulletin, 2014, 20(12): 117 − 118. [3] 徐秀荣, 杨克彬, 王思宁, 等. 毛竹bHLH转录因子的鉴定及其在干旱和盐胁迫条件下的表达分析[J]. 植物科学学报, 2019, 37(5): 610 − 620. XU Xiurong, YANG Kebin, WANG Sining, et al. Identification of bHLH transcription factors in moso bamboo (Phyllostachys edulis) and their expression analysis under drought and salt stress [J]. Plant Science Journal, 2019, 37(5): 610 − 620. [4] 吕玉龙. 高温干旱对毛竹林的危害及抗旱经营措施建议[J]. 林业实用技术, 2014(8): 53 − 55. LÜ Yulong. Harm of high temperature and drought on moso bamboo forest and suggestions on drought resistance management [J]. Practical Forestry Technology, 2014(8): 53 − 55. [5] 毛美红, 丁笑章, 傅柳方, 等. 干旱对毛竹林新竹成竹影响的调查分析[J]. 世界竹藤通讯, 2012, 10(1): 12 − 15. MAO Meihong, DING Xiaozhang, FU Liufang, et al. Investigation of the effect of drought on new moso forest cultivation [J]. World Bamboo and Rattan, 2012, 10(1): 12 − 15. [6] CHEN Yiyun, LI Mengyao, WU Xuejun, et al. Genome-wide analysis of basic helix-loop-helix family transcription factors and their role in responses to abiotic stress in carrot [J]. Molecular Breeding, 2015, 35(5): 1 − 12. [7] SONNENFELD M J, DELVECCHIO C, SUN Xuetao. Analysis of the transcriptional activation domain of the Drosophila tango bHLH-PAS transcription factor [J]. Development Genes and Evolution, 2005, 215(5): 221 − 229. [8] BAILEY P C, MARTIN C, TOLEDO-ORTIZ G, et al. Update on the basic helix-loop-helix transcription factor gene family in Arabidopsis thaliana [J]. The Plant Cell, 2003, 15(11): 2497 − 2502. [9] LI Xiaoqing, TANG Yuanping, YUAN Zheng, et al. Genome-wide analysis of basic/helix-loop-helix transcription factor family in rice and Arabidopsis [J]. Plant Physiology, 2006, 141(4): 1167 − 1184. [10] LEDENT V, VERVOORT M. The basic helix-loop-helix protein family: comparative genomics and phylogenetic analysis [J]. Genome Research, 2001, 11(5): 754 − 770. [11] RIECHMANN J L, HEARD J E, MARTIN G, et al. Arabidopsis transcription factors: genome-wide comparative analysis among eukaryotes [J]. Science, 2000, 290(5499): 2105 − 2110. [12] AN Jianping, LI Haohao, SONG Laiqing, et al. The molecular cloning and functional characterization of MdMYC2, a bHLH transcription factor in apple [J]. Plant Physiology and Biochemistry, 2016, 108: 24 − 31. [13] KONDOU Y, NAKAZAWA M, KAWASHIMA M, et al. RETARDED GROWTH OF EMBRYO1, a new basic helix-loop-helix protein, expresses in endosperm to control embryo growth [J]. Plant Physiology, 2008, 147(4): 1924 − 1935. [14] HEISLER M G, ATKINSON A, BYLSTRA Y H, et al. SPATULA, a gene that controls development of carpel margin tissues in Arabidopsis, encodes a bHLH protein [J]. Development, 2001, 128(7): 1089 − 1098. [15] WANG Houping, LI Yang, PAN Jinjing, et al. The bHLH transcription factors MYC2, MYC3, and MYC4 are required for jasmonate-mediated inhibition of flowering in Arabidopsis [J]. Molecular Plant, 2017, 10(11): 1461 − 1464. [16] TOLEDO-ORTIZ G, HUQ E, QUAIL P H. The Arabidopsis basic/helix-loop-helix transcription factor family [J]. Plant Cell, 2003, 15(8): 1749 − 1770. [17] HEIM M A, JAKOBY M, WERBER M, et al. The basic helix-loop-helix transcription factor family in plants: a genome-wide study of protein structure and functional diversity[J]. Molecular Biology and Evolution, 20(5): 735 − 747. [18] FAN Yu, YANG Hao, LAI Dili, et al. Genome-wide identification and expression analysis of the bHLH transcription factor family and its response to abiotic stress in sorghum [Sorghum bicolor (L. ) Moench] [J/OL]. BMC Genomics, 2021, 22: 415[2022-07-30]. doi: 10.1186/s12864-021-07652-9. [19] ZHAO Hansheng, GAO Zhimin, WANG Le, et al. Chromosome-level reference genome and alternative splicing atlas of moso bamboo (Phyllostachys edulis) [J/OL]. GigaScience, 2018, 7(10): giy115[2022-07-30]. doi: 10.1093/gigascience/giy115. [20] RYCHLIK W. OLIGO 7 primer analysis software [J]. PCR Primer Design, 2007, 402: 35 − 59. [21] GUO Zhenhua, MA Pengfei, YANG Guoqian, et al. Genome sequences provide insights into the reticulate origin and unique traits of woody bamboos [J]. Molecular Plant, 2019, 12(10): 1353 − 1365. [22] CHEN Chengjie, CHEN Hao, ZHANG Yi, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data [J]. Molecular Plant, 2020, 13(8): 1194 − 1202. [23] LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the \begin{document}$2^{-\Delta\Delta C{t}} $\end{document} [24] FAN Chunjie, MA Jinmin, GUO Qirong, et al. Selection of reference genes for quantitative real-time PCR in bamboo (Phyllostachys edulis) [J/OL]. PLoS One, 2013, 8(2): e56573[2022-07-31]. doi: 10.1371/journal. pone. 0056573. [25] MAO Ke, DONG Qinglong, LI Chao, et al. Genome wide identification and characterization of apple bHLH transcription factors and expression analysis in response to drought and salt stress[J/OL]. Front in Plant Science, 2017, 8: 480[2022-07-30]. doi: 10.3389/fpls.2017.00480. [26] HE Qiuju, LU Hong, GUO Huaxing, et al. OsbHLH6 interacts with OsSPX4 and regulates the phosphate starvation response in rice [J]. The Plant Journal, 2020, 105(3): 649 − 667. [27] 张子佳, 王迪, 傅彬英. 水稻转录因子bHLH家族基因响应环境胁迫表达谱分析[J]. 分子植物育种, 2008, 6(3): 425 − 431. ZHANG Zijia, WANG Di, FU Binying. Expression patterns of rice bHLH genes responsive to environmental stresses [J]. Molecular Plant Breeding, 2008, 6(3): 425 − 431. [28] 李朝霞, 高强, 刘雅正, 等. 玉米 ZmPTF1 基因克隆和过表达分析[J]. 湖南农业大学学报(自然科学版), 2007, 33(1): 92 − 96. LI Zhaoxia, GAO Qiang, LIU Yazheng, et al. Cloning of ZmPTF1 from Zea mays and its over expression analysis [J]. Journal of Hunan Agricultural University (Natural Sciences), 2007, 33(1): 92 − 96. [29] MENG Fanwei, YANG Chao, CAO Jidong, et al. A bHLH transcription activator regulates defense signaling by nucleo-cytosolic trafficking in rice [J]. Journal of Integrative Plant Biology, 2020, 62(10): 1552 − 1573. [30] WU Hua, YE Haiyan, YAO Ruifeng, et al. OsJAZ9 acts as a transcriptional regulator in jasmonate signaling and modulates salt stress tolerance in rice [J]. Plant Science, 2015, 232: 1 − 12. [31] KIRIBUCHI K, SUGIMORI M, TAKEDA M, et al. RERJ1, a jasmonic acid-responsive gene from rice, encodes a basic helix-loop-helix protein [J]. Biochemical and Biophysical Research Communications, 2004, 325(3): 857 − 863. [32] KIRIBUCHI K, JIKUMARU Y, KAKU H, et al. Involvement of the basic helix-loop-helix transcription factor RERJ1 in wounding and drought stress responses in rice plants [J]. Bioscience,Biotechnology,and Biochemistry, 2005, 69(5): 1042 − 1044. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220553
 点击查看大图
点击查看大图
计量
- 文章访问数: 2235
- HTML全文浏览量: 365
- PDF下载量: 360
- 被引次数: 0
