












-
bZIP转录因子家族是植物中最大、最保守且功能最多样的基因家族之一,广泛参与调节多种生物学过程[1-2]。ABF (ABRE binding factor)转录因子属于bZIP基因家族的A亚族[3],特异存在于植物中,ABFs是ABA信号转导途径的关键基因,能够特异识别ABA响应元件(ABA-responsive element,AREB),并通过与AREB结合参与下游响应ABA信号靶基因的调控[4]。月季Rosa chinensis中RhABF2基因能够与含有ABRE元件的RhFer1启动子结合,共同调节铁平衡参与调控花瓣衰老过程[5];睡莲Nymphaea colorata bZIP家族A亚组的11个成员的启动子区域有大量ABA响应元件,在ABA信号转导中具有重要作用[6];菊花Chrysanthemum×morifolium中锌指蛋白BBX19和ABF3互作,通过诱导ABA响应基因表达来调控菊花的耐旱性[7]。桂花Osmanthus fragrans属于木犀科Oleaceae木犀属Osmanthus常绿乔木,是中国传统十大名花之一。温度、湿度是影响桂花花开放的重要因素。为了解ABFs转录因子在桂花中的作用,本研究以秋桂品种‘堰虹桂’O. fragrans‘Yanhonggui’为材料,在桂花基因组中筛选出相关OfABFs基因序列,通过实时荧光定量PCR(RT-qPCR)分析OfABFs基因在‘堰虹桂’不同组织和花开放进程中的时空表达模式,为后续阐明ABFs基因家族各成员的生物学功能提供理论支持。
-
以浙江农林大学桂花资源圃的桂花品种‘堰虹桂’盆栽植物为材料,选取株龄相同、长势一致的健康植株,分别于起始1期(圆珠期前3 d,B1),起始2期(圆珠期前2 d,B2),起始3期(圆珠期前1 d,B3),圆珠期(S1),顶壳期(S2),铃梗期(S3),初开期(S4),盛开期(S5),盛开末期(S6),衰老期(S7)共10个时期进行取样(图1)[8]。同样,取桂花不同组织的样品,包括根、新叶、新枝、老叶、老枝、盛开期的花朵,每个时期样品采集3份,液氮速冻后储藏于−80 ℃冰箱。

图 1 桂花花开放各时期表型图
Figure 1. Phenotypic of O. fragrans in different periods
-
桂花核苷酸序列来源于桂花基因组数据库[9],从拟南芥Arabidopsis thaliana信息资源(TAIR)数据库和PlantTFDB 数据库中获得拟南芥的ABFs氨基酸序列,以此作为查询对象,使用NCBI-Blast (
https://blast.ncbi.nlm.nih.gov/Blast.cgi )在桂花基因组蛋白数据库中进行同源比对,确定桂花中的ABFs基因序列。利用NCBI-CDD (https://www.ncbi. nlm.nih.gov/cdd/ )对OfABFs进行保守功能域预测。使用SMART (http://smart.embl-heidelberg.de/ )分析OfABFs蛋白结构域。RNA的提取使用RNA prep Pure Plant Plus Kit提取试剂盒[天根生化科技(北京)有限公司],使用核酸分析仪(赛默飞世尔科技公司)对RNA的纯度与浓度进行检测,并用质量分数为1%的琼脂糖凝胶电泳检测RNA的完整性。使用HiScriptⅡ Q Select RT SuperMix for qPCR(+gDNA wiper)试剂盒(南京诺维赞生物科技股份公司),对花开放不同时期的cDNA进行合成。
运用Primer 5.0进行简单克隆引物设计(表1),以花开放时期的花芽cDNA为模板进行PCR扩增。PCR反应体系(20 μL):上下游引物(10 μmol·L−1)各1 μL、cDNA模板1 μL、GreenTaqMix10 μL和ddH2O 7 μL。PCR扩增程序:预变性95 ℃,3 min;变性95 ℃,15 s,退火温度以各自引物最佳温度为准,15 s,延伸72 ℃,3 min,35个循环;72 ℃延伸5 min。将PCR反应产物进行1%琼脂糖凝胶电泳检测,目的片段回收纯化后与pMD-18T载体连接,连接产物转化大肠埃希菌Escherichia coli DH5α感受态细胞,PCR初步鉴定阳性克隆后送杭州有康生物科技有限公司测序,经比对后确定序列。
表 1 桂花OfABFs简单克隆引物
Table 1. Primers for Cloning of OfABFs in O. fragrans
基因名称 引物序列(5′→3′) 产物长度/bp 引物温度/℃ OfABF1 F: ATGGGGAGTAATTTGAACTTCAAGAATCCG
R: TTACCATGGACCTGTCTGAGTTCGTCTCA1 557 54 OfABF2 F: ATGGGGTCATACATGAACTTCAAG
R: CTACCAAGGTCCCGTCAGCA1 353 58 OfABF3 F: ATGGGGAGTAATTTTAACTTCAAGAATTTTGG
R: TTACCATGGCCCTGTCTGTGTACGTC1 455 56 OfABF4 F: ATGTGGAGTAATTTTAACTTCAAGAATGT
R: TCACCATGGCCCCGTTTG1 572 56 OfABF6 F: ATGAACTTTGGGCCGGACACATCA
R: CTACCAAGGTCCTGTCAGCGTCCTTCT1 329 45 -
利用TBtools[10]从桂花基因组Gff文件中提取桂花OfABFs基因的染色体位置信息,通过在线分析工具TBtools进行绘图可视化。
-
用DNAMAN对已筛选的桂花OfABFs转录因子进行氨基酸序列比对,分析其保守结构域和氨基酸序列同源性;通过在线分析软件MEME 5.1.0分析ABFs的保守基序(
http://meme-suite.org/tools/meme ),并用TBtools对其进行绘图和可视化。对桂花OfABFs二级结构特征采用在线软件SOPMA 进行预测分析;使用在线网站Swiss-Model (https://swissmodel.expasy.org/ )构建OfABFs蛋白的三级结构模型。 -
运用ExPASy在线软件(
http://web.expasy.org/protparam/ )对OfABFs进行蛋白质、分子量、理论电点、酸碱性等预测分析和编码;利用SignalP 4.1 Server分析桂花OfABFs蛋白有无信号肽。亚细胞定位预测通过在线工具wolfpsort (https:// wolfpsort.hgc.jp/ )完成。 -
从美国国家生物信息中心(NCBI) (www.ncbi.nlm.nih.gov)中获取拟南芥、烟草Nicotiana tabacum、水稻Oryza sativa、大麦Hordeum vulgare、小麦Triticum aestivum、番茄Solanum lycopersicum、油橄榄Olea europaea的ABFs氨基酸序列,使用MEGA 11.0软件进行同源聚类,建立系统发育树并选择Bootstrap (1 000次)评估检测系统进化树。
-
利用Primer Premier 5.0对5个OfABFs基因进行荧光定量特异性引物设计(表2),桂花OfACT作为内参基因[11],以‘堰虹桂’不同组织及不同花开放时期的花芽cDNA为模板进行RT-qPCR扩增。反应体系为20 μL:cDNA模板2 μL,2×Taq Pro Universal SYBR qPCR Master Mix 10 μL,上下游引物(10 μmol·L−1)各0.4 μL,ddH2O 7.2 μL。RT-qPCR程序:95 ℃ 30 s;95 ℃ 5 s,60 ℃ 30 s,共40个循环;然后以95 ℃持续15 s,60 ℃持续1 min,95 ℃持续15 s作为熔解曲线分析程序。每个样品设置3次重复。
表 2 桂花OfABFs转录因子RT-qPCR特异性引物
Table 2. Specific primers for RT-qPCR of OfABFs in O. fragrans
基因名称 用于荧光定量的引物序列(5′→3′) OfABF1 F: GGAAGGATGGGAGTGGTAGTG R: TTAGGCTTTCCAGCCAACTGA OfABF2 F: TGGTGCAAAAGATGGAAGTGC R: ATTTGCTGTCCAAGAGGGCTG OfABF3 F: GAGTATGGTGGCGGGAAGAAT R: GGTAGGCTTTCCAGCAAACTG OfABF4 F: TCTGGAGAGAGAGGTGGAATGA R: CAAACTGGGCATCTTCCCTTAC OfABF6 F: GAAGGGGTAGTGGAGGATCAA R: ACCACCATAAATTCCACCAGC -
通过对桂花基因组数据分析,得到5条OfABFs基因序列,依次命名为OfABF1、OfABF2、OfABF3、OfABF4、OfABF6。用‘堰虹桂’花开放时期的花芽(混样) cDNA为模板进行PCR扩增(图2)。结果显示:OfABFs的开放阅读框(ORF)长度为1329~1572 bp,编码442~523个氨基酸;其中基因OfABF4氨基酸序列最长,编码523个氨基酸;基因OfABF6的氨基酸序列最短,编码442个氨基酸。

图 2 桂花OfABFs基因片段PCR扩增
Figure 2. PCR amplification of OfABFs of O. fragrans
-
根据‘堰虹桂’基因组Gff文件定位5个OfABFs基因。结果显示:5个桂花OfABFs基因分布在5条染色体上,且各个基因在染色体上分布的数量相等,1号、6号、11号、12号和22号染色体上各分布1个OfABF基因(图3)。

图 3 桂花OfABFs基因家族染色体定位分析
Figure 3. Chromosome location analysis of OfABFs genes in O. fragrans
-
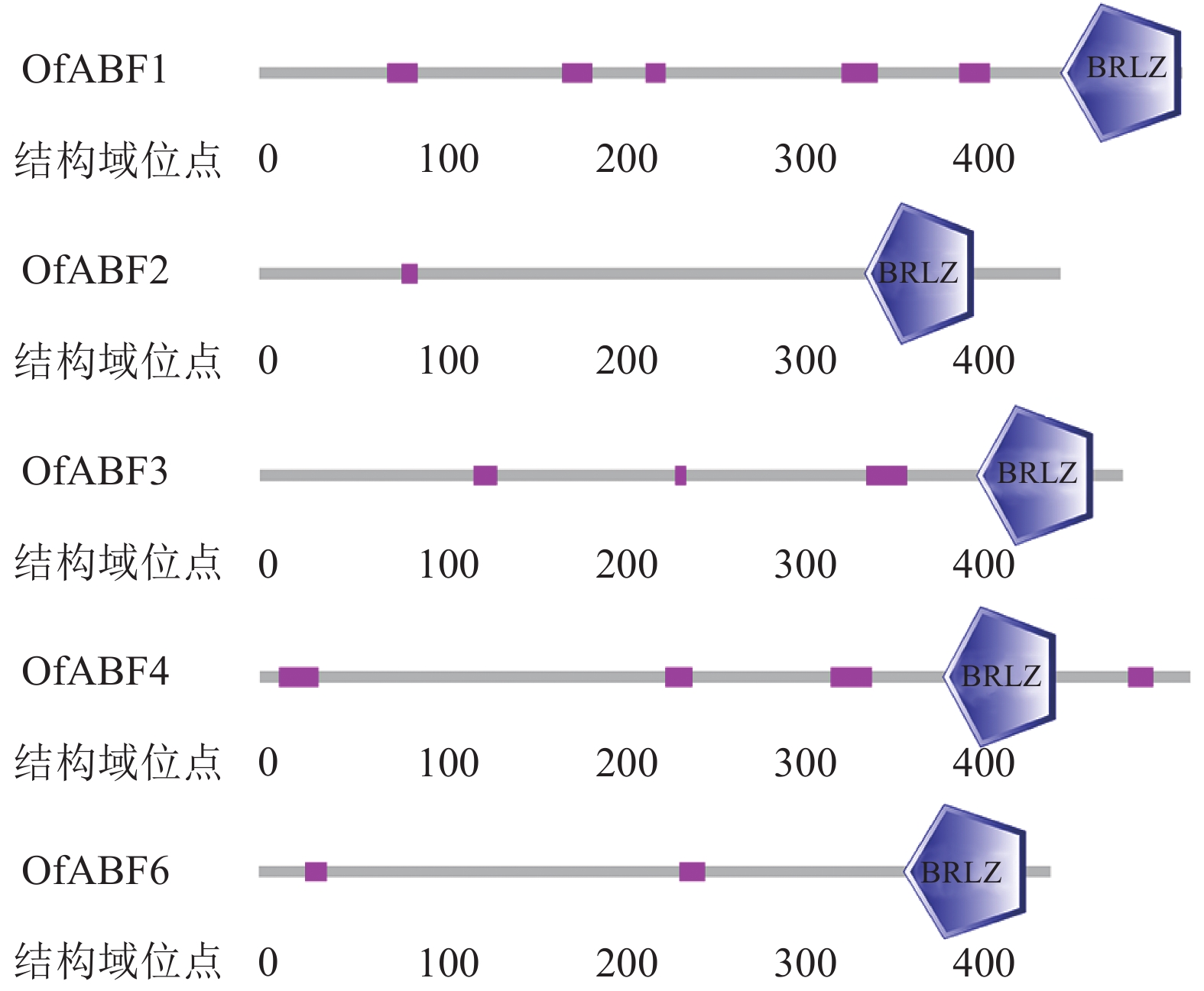
利用SMART在线分析软件对桂花OfABFs进行功能结构域预测。结果显示:5个桂花ABFs均含有BRLZ结构域(图4)。BRLZ结构域是bZIP家族特有的保守域,由碱性区和亮氨酸拉链区组成,能够识别特定的DNA序列,以二聚体形式发挥功能[12]。通过DNAMAN软件,将OfABFs氨基酸序列进行比对(图5)。结果显示:OfABF3和OfABF4同源性最高,OfABF1和OfABF6同源性最低;5个C端均含有能被激酶识别的保守序列RXXS/T。
图 4 桂花OfABFs功能结构域分析
Figure 4. Functional domains of OfABFs in O. fragrans

图 5 桂花OfABFs转录因子氨基酸序列比对
Figure 5. Sequence alignment of OfABFs transcription factor in O. fragrans
对桂花ABFs氨基酸序列进行保守基序分析发现(图6):OfABFs蛋白结构域中含有10个蛋白基序(Motif 1~Motif 10),其中Motif 7在除OfABF4外的4个蛋白中均被鉴定到,Motif 8在除OfABF6外的4个蛋白中均被鉴定到,其余8个Motif高度保守,是OfABFs核心结构域的组成部分。

图 6 桂花OfABFs蛋白质保守结构域
Figure 6. Conserved motifs OfABFs proteins in O. fragrans
桂花OfABFs蛋白的二级结构预测表明:桂花OfABFs蛋白的二级结构均含有α-螺旋、β-折叠、延伸链、无规则卷曲,不同结构占比从大到小依次为无规则卷曲、α-螺旋、延伸链、β-折叠。其中,β-折叠占比最小,均在5%以下;延伸链占比为10%~13%;α-螺旋和无规则卷曲占二级结构总量的83%,为OfABFs蛋白二级结构的主要构成元件(表3)。
表 3 桂花OfABFs蛋白二级结构分析
Table 3. Secondary structure analysis of protein OfABFs in O. fragrans
基因名称 α-螺旋/% β-折叠/% 延伸/% 无规则卷曲/% 基因名称 α-螺旋/% β-折叠/% 延伸/% 无规则卷曲/% OfABF1 27.80 4.05 13.32 54.83 OfABF4 28.49 4.78 11.85 54.88 OfABF2 28.67 1.78 12.22 57.33 OfABF6 31.67 3.17 10.86 54.30 OfABF3 30.99 3.10 11.16 54.75 桂花OfABFs蛋白三级结构分析显示(图7):OfABFs蛋白三级结构均含丰富的α-螺旋和无规则卷曲,这与其二级结构预测结果一致。含无规则卷曲较多的蛋白稳定性要低于含α-螺旋、β-折叠多的蛋白,在OfABFs蛋白中无规则卷曲比例均高于50%,为不稳定蛋白,这与其理化性质分析预测结果一致(表3)。
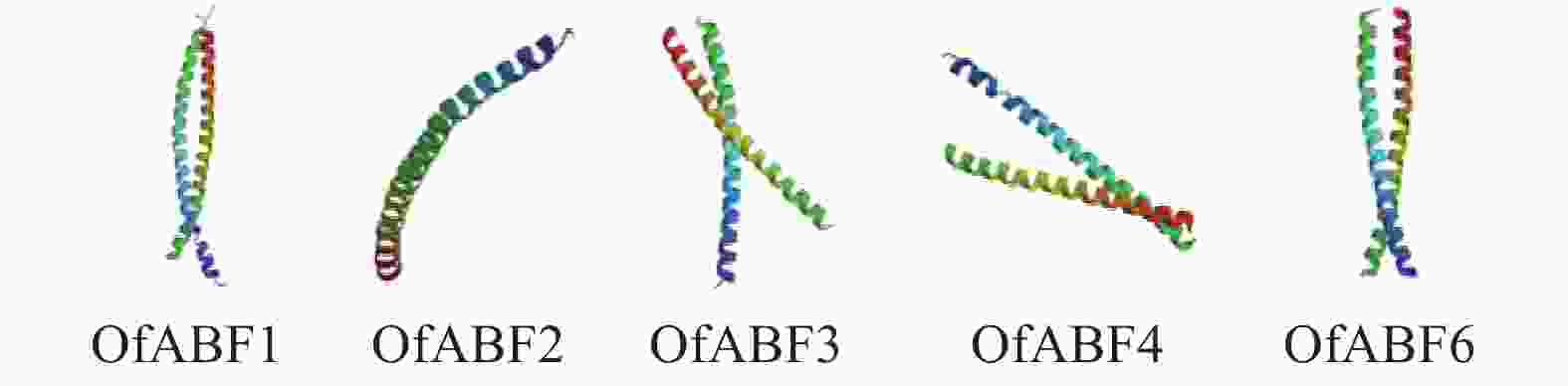
图 7 桂花OfABFs蛋白三级结构预测
Figure 7. Prediction of tertiary structure of the OfABFs protein in O. fragrans
-
对桂花ABFs家族蛋白理化性质的分析结果表明:桂花氨基酸长度为450~523 个,相对分子质量为25.7~56.2 kDa,理论等电点为4.77~10.03,编码氨基酸序列最长的的是OfABF4,其相对分子质量最大,为56.2 kDa;编码氨基酸序列最短的是OfABF2,其相对分子质量也最小,为25.6 kDa。5个OfABFs基因成员的不稳定系数为46.76~54.43。如果将不稳定系数>40的蛋白判断为不稳定蛋白,那么5个基因的蛋白不稳定系数均高于40,均为不稳定蛋白。桂花ABFs蛋白脂溶性系数为65.29~78.07,亲水性总平均系数为−0.659~−0.482,均小于0,为亲水性蛋白。信号肽分析结果显示:桂花ABFs编码蛋白均不含有信号肽,表明桂花ABFs家族蛋白均非分泌蛋白。亚细胞定位预测发现(表4):5个编码蛋白均定位在细胞核,这与其作为转录因子的生物学功能相吻合。
表 4 桂花OfABFs蛋白理化性质分析
Table 4. Physicochemical properties of OfABFs proteins of O. fragrans
基因名称 序列名称 氨基酸
长度/个相对分
子量/kDa等电点 酸性氨
基酸/个碱性氨
基酸/个不稳定
系数脂溶性
系数亲水性平
均系数亚细胞定
位预测信号肽 OfABF1 gui0017720.1 518 55.6 9.24 49 56 46.76 78.07 −0.482 细胞核 无 OfABF2 gui0167370.1 450 25.7 4.77 25 16 46.95 71.04 −0.536 细胞核 无 OfABF3 gui0180650.1 484 51.9 9.35 43 50 52.80 67.77 −0.615 细胞核 无 OfABF4 gui0305040.1 523 56.2 5.38 60 52 50.61 68.66 −0.659 细胞核 无 OfABF6 gui0100630.1 442 46.8 10.03 35 50 54.43 65.29 −0.593 细胞核 无 -
构建系统进化树进一步研究OfABFs的进化关系,结果表明(图8):8个物种的25条氨基酸序列可分为5个分支(分支Ⅰ~Ⅴ),其中OfABFs分布在2个分支中。OfABF1、OfABF3和OfABF4均分布在分支Ⅰ,该分支包括油橄榄OeTRAB1、烟草NtABF2、马铃薯Solanum tuberosum StABF、马铃薯StABF1、番茄SlAREB1和拟南芥AtABF2,说明OfABF1、OfABF3和OfABF4在进化过程中与这些物种亲缘关系较近;OfABF2和OfABF6与番茄SlABF4聚为分支Ⅱ。
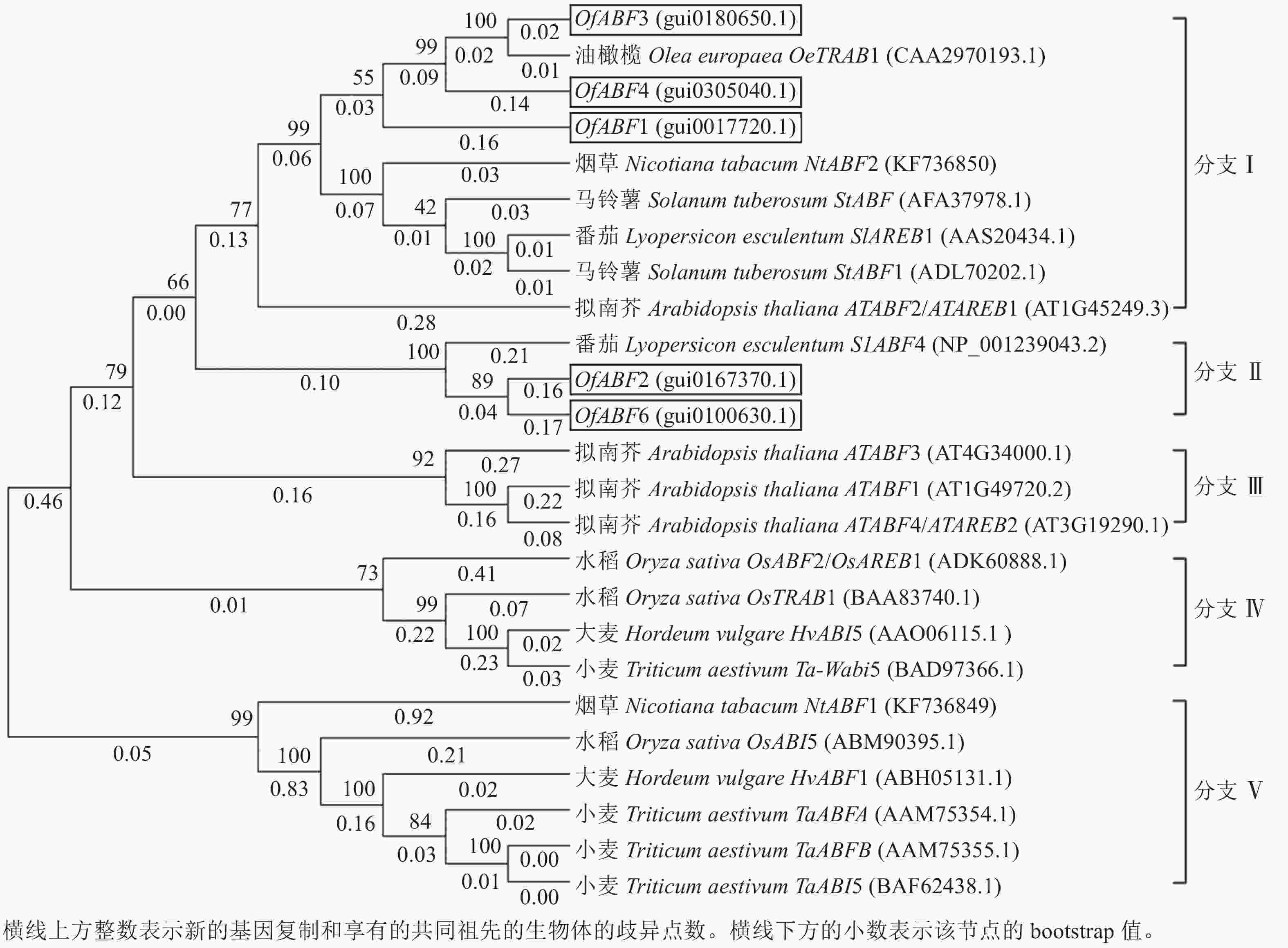
图 8 桂花OfABFs与各物种ABFs转录因子进化分析
Figure 8. Studies on the relationship between O. fragrans OfABFs and the evolution of ABFs transcription factors in different species
-
利用荧光定量分析5个基因在根、嫩枝、老枝、新叶、老叶等不同组织中的表达情况。定量结果(图9)显示:OfABF1在花中表达量最高,在老叶和新叶中表达没有显著差异,在新枝和老枝中相对表达量最低;OfABF2在老叶中表达最高,其次为新叶,在花中表达略高于枝;OfABF3在叶中表达水平相对其他组织高,在花和枝中表达较低。OfABF4在花中相对表达量最高,其次是新叶和老叶,老枝中表达量最低,OfABF6的相对表达量在花中最高,在其他组织中的表达量均较低。OfABF1、OfABF4、OfABF6在所有组织中均有表达,但在花中的相对表达水平最高,尤其是OfABF6。
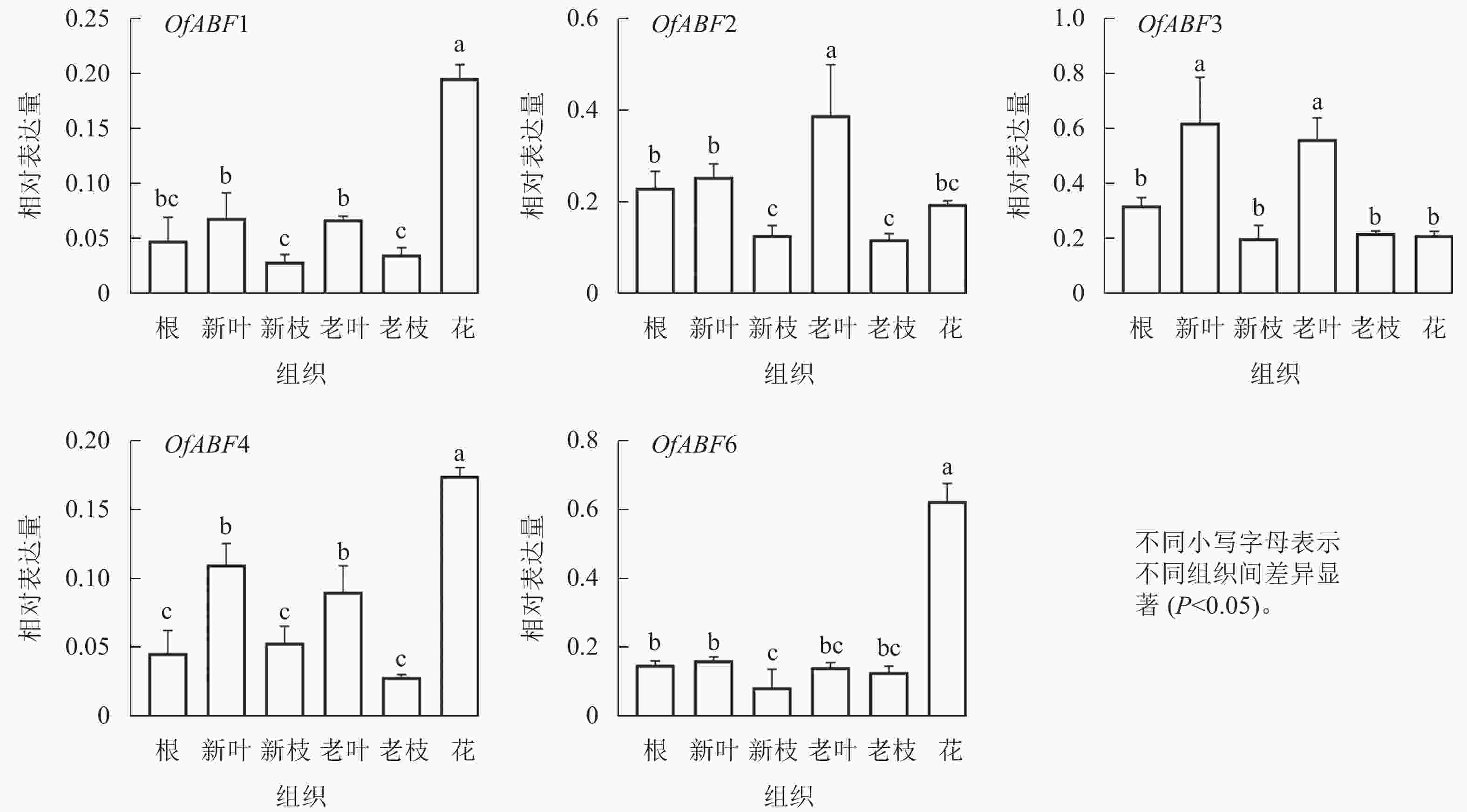
图 9 5个OfABFs基因在不同组织的表达结果
Figure 9. Expression results of 5 OfABFs genes in different tissues
由图10可见:起始1期(B1)到起始3期(B3)花芽芽体肥大并延伸。圆珠期(S1)到盛开末期(S6)为花开放时期,OfABF1在顶壳期(S2)被转录激活,表达显著升高,铃梗期(S3)后表达水平虽呈下降趋势,但仍保持较高水平;OfABF2在起始1期(B1)到圆珠期(S1)时期几乎不表达,在顶壳期(S2)的表达量到达峰值,且相对表达水平显著高于其他时期;OfABF3和OfABF4在花朵衰老期(S7)的相对表达量最高,在盛开末期(S6)的表达量其次,圆珠期(S1)至盛开期(S5)表达趋势变化不显著;OfABF6的相对表达量在花朵衰老期(S7)最高。由此说明:OfABF1和OfABF2可能与桂花花开放进程有关,OfABF3、OfABF4和OfABF6可能参与桂花花衰老的调控。
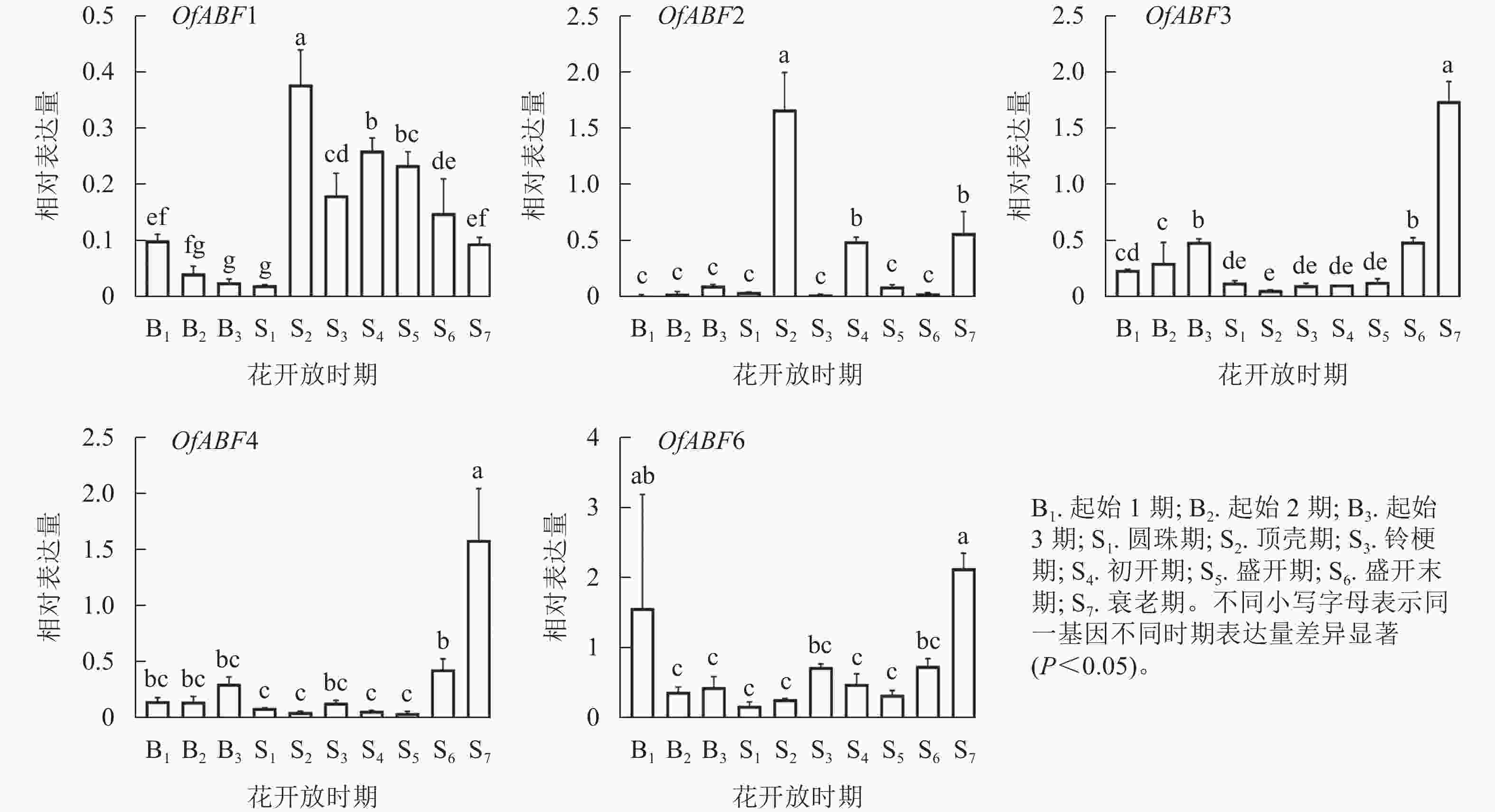
图 10 5个OfABFs基因在花开放时期的表达结果
Figure 10. Express an results of 5 OfABFs genes in different flower opening periods
-
ABF转录因子在模式植物拟南芥、烟草以及园艺作物中的研究较多。ABFs已在拟南芥、烟草、杨树Populus中被鉴定[12-15]。LI等[16]对29种陆地植物中的95种ABFs蛋白长度、分子量进行分析,发现ABF的氨基酸长度为254~485 个;分子量为27.81~52.95 kDa。目前,关于桂花OfABFs的研究较少,通过鉴定与分析桂花OfABF转录因子,可进一步了解桂花OfABFs的生物学功能及其表达分析。
OfABFs蛋白的二级结构组成与烟草、睡莲、野菊Chrysanthemum indicum、蓝莓Vaccinium corymbosum、大豆Glycine max[6, 17-20]等其他物种中的结构相类似,均以α-螺旋和无规则卷曲为主要构成元件,少量β-折叠和延伸链散布于整个多肽链中。说明在基因变异和进化时,虽然基因外显子的数量上发生变化,但仍具有其保守性,这与杨颖等[2]的研究结果基本一致。基于蛋白行使特定功能依赖其三维结构,本研究使用蛋白三级结构预测软件SWISS-MODEL同源模拟桂花OfABFs蛋白三级结构,结果显示:OfABF2与烟草[17]中的同源蛋白质NtABF1、NtABF2三级结构类似,除OfABF2蛋白外,其余成员蛋白三级结构在空间结构上相似度较高,进一步说明ABF蛋白在进化上较保守,具有对植物生长发育的重要功能。序列比对结果显示:5个OfABFs的C端均具有bZIP转录因子特有的BRLZ结构域,且含有保守序列RXXS/T,推测OfABFs转录因子可通过被磷酸化激活,从而结合下游基因启动子区域的ABRE元件,调控相关下游胁迫响应基因[20-21]。
系统发育结果显示:桂花ABFs基因家族成员全部分布在以双子叶植物油橄榄、番茄、马铃薯、烟草为主的分支Ⅰ和分支Ⅱ中,无家族成员分布在以单子叶植物小麦、大麦、水稻等为主的分支Ⅲ、分支Ⅳ和分支Ⅴ中。由此推测,桂花ABFs与双子叶植物亲缘关系较近,与单子叶植物的亲缘关系较远。表明在进化过程中,ABF在种属间是高度保守的。
根据亲缘关系推测桂花OfABFs基因功能,如在分支Ⅰ中,OfABF1、OfABF3、OfABF4与马铃薯StABF、番茄SlAREB1、烟草NtABF2、拟南芥AtAREB1亲缘性较高。在对环境的胁迫应答过程中,番茄SlAREB1的过表达可增加物种干旱和盐胁迫耐受性[22],诱导有机酸积累的变化并参与其合成的基因编码酶的表达[23];烟草NtABF2与拟南芥AtAREB1响应非生物胁迫,前者通过与胁迫响应基因启动子区的ABRE元件结合相关基因的转录[16, 23-24],后者受ABA含量等渗透胁迫诱导,作为关键基因参与ABA信号调控通路以增强物种抗旱性[25],显著提高ABA依赖性逆境响应基因的表达水平[25-28];在分支Ⅱ中,OfABF2、OfABF6与番茄SlABF4同源性较高,且SlABF4作为ABA响应因子,ABA通过其对SICOL4产生负调控作用,使其与下游乙烯通路相关的基因互作[29],调控乙烯的合成和信号转导进而调控番茄果实的成熟进程[29-30]。由此推测:桂花OfABFs可以作为ABA响应因子,参与ABA信号转导,影响植物整个生长发育过程[31]。牡丹Paeonia suffruticosa切花保鲜的研究发现:ABA以诱导乙烯大量合成的方式间接加速牡丹切花的衰老[32]。还有研究发现:在月季花瓣衰老过程中内源ABA含量迅速上升,促进RhABF2的表达量上调,从而推测RhABF2在月季花瓣衰老过程中发挥调节作用[7]。在桂花中OfABF3、OfABF4和OfABF5的相对表达量在衰老期急剧上升,推测在桂花花瓣衰老过程中内源ABA含量上升,OfABF3、OfABF4和OfABF5可能参与调节桂花花瓣的衰老。推测OfABFs基因响应内源ABA,参与桂花衰老的可能性较大。5个OfABFs基因在桂花中的功能值得进一步探讨。
-
以桂花品种‘堰虹桂’为材料,在基因组数据库中筛选出相关5条OfABFs基因序列。生物信息学分析得出:桂花ABFs蛋白具有亲水性,较不稳定,无信号肽段。其蛋白二、三级结构预测具有相似特性,无规则卷曲和α-螺旋为其结构组成的主要元件;5条OfABFs基因序列均含有bZIP转录因子家族保守结构域;亚细胞定位预测表明:5条OfABFs编码蛋白均定位在细胞核。荧光定量PCR结果表明:OfABF1、OfABF2、OfABF3、OfABF4和OfABF6在桂花中的表达具有组织特异性,其中OfABF1、OfABF4和OfABF6在花中表达较高。OfABF1在顶壳期(S2)被转录激活,表达显著升高,铃梗期(S3)后表达水平虽呈下降趋势,但仍保持较高水平;OfABF2在顶壳期(S2)表达量达到峰值,且相对表达水平著显高于其他时期;OfABF3、OfABF4和OfABF6在花朵衰老期(S7)的相对表达量最高。推测OfABF1、OfABF2可能与桂花花开放进程有关,而OfABF3、OfABF4和OfABF6响应ABA参与调控花朵衰老的可能性较大。
Cloning and expression analysis of OfABFs gene in Osmanthus fragrans
-
摘要:
目的 研究OfABFs基因家族成员在桂花‘堰虹桂’Osmanthus fragrans ‘Yanhonggui’不同组织和花开放进程中的表达模式,筛选参与调控花开放的关键成员。 方法 在桂花基因组数据库中筛选出相关OfABFs基因序列,克隆序列全长并对其进行生物信息学分析,通过荧光定量PCR分析OfABFs基因在‘堰虹桂’不同组织及花开放进程中的时空表达模式。 结果 筛选得到5条OfABFs基因序列。生物信息学分析得出:桂花OfABFs蛋白具有亲水性,较不稳定,无信号肽段;预测其蛋白二、三级结构具有相似的特性,无规则卷曲和α-螺旋为其结构组成的主要元件;5条OfABFs转录因子均含有bZIP转录因子家族保守结构域。亚细胞定位预测表明:5条OfABFs编码蛋白均定位在细胞核。荧光定量PCR结果表明:OfABF1、OfABF2、OfABF3、OfABF4和OfABF6在桂花中的表达具有组织特异性,其中OfABF1、OfABF4和OfABF6在花中有较高表达。OfABF1在顶壳期被转录激活,表达显著升高;铃梗期后表达水平虽呈下降趋势,但仍保持较高水平。OfABF2的表达在顶壳期到达峰值,且相对表达水平明显高于其他时期。OfABF3、OfABF4和OfABF6在花朵衰老期的相对表达量最高。 结论 OfABF1、OfABF2可能与桂花花开放进程有关,而OfABF3、OfABF4和OfABF6参与调控桂花花朵衰老的可能性较大。图10表4参32 Abstract:Objective The aim of this study is to explore the expression patterns of OfABF gene family members in different tissues and flower opening process of Osmanthus fragrans ‘Yanhonggui’, and screen the key members involved in regulating flower opening. Method The related OfABFs gene sequence was screened from the O. fragrans genome database for the full-length sequence and bioinformatics analysis. The spatio-temporal expression mode of OfABFs gene in different tissues and flower opening process of O. fragrans ‘Yanhonggui’ were analyzed by fluorescence quantitative PCR. Result 5 OfABFs sequences were screened. Bioinformatics analysis showed that O. fragrans OfABFs gene family protein was hydrophilic and unstable, and had no signal peptide. It was predicted that the protein secondary and tertiary structure had similar characteristics and irregular curl, and α-spiral was the main component of its structure. The 5 OfABFs transcription factors all contained the conserved domain of bZIP transcription factor family. Subcellular localization prediction showed that 5 OfABFs coding proteins were located in the nucleus. The results of fluorescence quantitative PCR showed that the expressions of OfABF1, OfABF2, OfABF3, OfABF4 and OfABF6 were tissue-specific, among which OfABF1, OfABF4 and OfABF6 were highly expressed in flowers. OfABF1 was transcriptionally activated in the apical shell stage, and its expression increased significantly. Although the expression level decreased after boll stem stage, it still maintained a high level. The expression of OfABF2 peaked in the apical shell stage, and the relative expression level was significantly higher than that in other stages. The relative expression levels of OfABF3, OfABF4 and OfABF6 were the highest in flower senescence stage. Conclusion OfABF1 and OfABF2 may be related to the blooming process of O. fragrans, while OfABF3, OfABF4 and OfABF6 are more likely to participate in the regulation of O. fragrans flower senescence. [Ch, 10 fig. 4 tab. 32 ref.] -
Key words:
- Osmanthus fragrans /
- ABF transcription factor /
- expression analysis /
- flower opening
-
图 3 桂花OfABFs基因家族染色体定位分析
Figure 3 Chromosome location analysis of OfABFs genes in O. fragrans
图 5 桂花OfABFs转录因子氨基酸序列比对
Figure 5 Sequence alignment of OfABFs transcription factor in O. fragrans
图 7 桂花OfABFs蛋白三级结构预测
Figure 7 Prediction of tertiary structure of the OfABFs protein in O. fragrans
图 8 桂花OfABFs与各物种ABFs转录因子进化分析
Figure 8 Studies on the relationship between O. fragrans OfABFs and the evolution of ABFs transcription factors in different species
图 9 5个OfABFs基因在不同组织的表达结果
Figure 9 Expression results of 5 OfABFs genes in different tissues
图 10 5个OfABFs基因在花开放时期的表达结果
Figure 10 Express an results of 5 OfABFs genes in different flower opening periods
表 1 桂花OfABFs简单克隆引物
Table 1. Primers for Cloning of OfABFs in O. fragrans
基因名称 引物序列(5′→3′) 产物长度/bp 引物温度/℃ OfABF1 F: ATGGGGAGTAATTTGAACTTCAAGAATCCG
R: TTACCATGGACCTGTCTGAGTTCGTCTCA1 557 54 OfABF2 F: ATGGGGTCATACATGAACTTCAAG
R: CTACCAAGGTCCCGTCAGCA1 353 58 OfABF3 F: ATGGGGAGTAATTTTAACTTCAAGAATTTTGG
R: TTACCATGGCCCTGTCTGTGTACGTC1 455 56 OfABF4 F: ATGTGGAGTAATTTTAACTTCAAGAATGT
R: TCACCATGGCCCCGTTTG1 572 56 OfABF6 F: ATGAACTTTGGGCCGGACACATCA
R: CTACCAAGGTCCTGTCAGCGTCCTTCT1 329 45  下载: 导出CSV
下载: 导出CSV
表 2 桂花OfABFs转录因子RT-qPCR特异性引物
Table 2. Specific primers for RT-qPCR of OfABFs in O. fragrans
基因名称 用于荧光定量的引物序列(5′→3′) OfABF1 F: GGAAGGATGGGAGTGGTAGTG R: TTAGGCTTTCCAGCCAACTGA OfABF2 F: TGGTGCAAAAGATGGAAGTGC R: ATTTGCTGTCCAAGAGGGCTG OfABF3 F: GAGTATGGTGGCGGGAAGAAT R: GGTAGGCTTTCCAGCAAACTG OfABF4 F: TCTGGAGAGAGAGGTGGAATGA R: CAAACTGGGCATCTTCCCTTAC OfABF6 F: GAAGGGGTAGTGGAGGATCAA R: ACCACCATAAATTCCACCAGC
下载: 导出CSV
表 3 桂花OfABFs蛋白二级结构分析
Table 3. Secondary structure analysis of protein OfABFs in O. fragrans
基因名称 α-螺旋/% β-折叠/% 延伸/% 无规则卷曲/% 基因名称 α-螺旋/% β-折叠/% 延伸/% 无规则卷曲/% OfABF1 27.80 4.05 13.32 54.83 OfABF4 28.49 4.78 11.85 54.88 OfABF2 28.67 1.78 12.22 57.33 OfABF6 31.67 3.17 10.86 54.30 OfABF3 30.99 3.10 11.16 54.75
下载: 导出CSV
表 4 桂花OfABFs蛋白理化性质分析
Table 4. Physicochemical properties of OfABFs proteins of O. fragrans
基因名称 序列名称 氨基酸
长度/个相对分
子量/kDa等电点 酸性氨
基酸/个碱性氨
基酸/个不稳定
系数脂溶性
系数亲水性平
均系数亚细胞定
位预测信号肽 OfABF1 gui0017720.1 518 55.6 9.24 49 56 46.76 78.07 −0.482 细胞核 无 OfABF2 gui0167370.1 450 25.7 4.77 25 16 46.95 71.04 −0.536 细胞核 无 OfABF3 gui0180650.1 484 51.9 9.35 43 50 52.80 67.77 −0.615 细胞核 无 OfABF4 gui0305040.1 523 56.2 5.38 60 52 50.61 68.66 −0.659 细胞核 无 OfABF6 gui0100630.1 442 46.8 10.03 35 50 54.43 65.29 −0.593 细胞核 无
下载: 导出CSV
-
[1] 刘金义. 葡萄bZIP转录因子家族的全基因组鉴定、表达分析及VvbZIP45/VvbZIP08(AREB/ABF类)基因的功能研究[D]. 南京: 南京农业大学, 2014. LIU Jinyi. Genome-wide Identification, Expression Analysis of bZIP Transcription Factor Family and Functional Characterization of VvbZIP45/VvbZIP08(AREB/ABF) Gene in Grapevine(Vitis vinifera spp.) [D]. Nanjing: Nanjing Agricultural University, 2014. [2] 杨颖, 高世庆, 唐益苗, 等. 植物bZIP转录因子的研究进展[J]. 麦类作物学报, 2009, 29(4): 730 − 737. YANG Yin, GAO Shiqing, TANG Yimiao, et al. Advance of bZIP transcription factors in plants [J]. Journal of Triticeae Crops, 2009, 29(4): 730 − 737. [3] 洪岚, 刘旭, 李玲. 植物AREB/ABF转录因子及其参与的ABA信号转导[J]. 植物生理学报, 2011, 47(3): 211 − 217. HONG Lan, LIU Xu, LI Ling. AREB/ABF transcription factors and their involvement in ABA signal transduction [J]. Plant Physiology Journal, 2011, 47(3): 211 − 217. [4] CHOI H I, HONG J H, HA J O, et al. ABFs, a family of ABA-responsive element binding factors [J]. Journal of Biological Chemistry, 2000, 275(3): 1723 − 1730. [5] 刘计涛. 月季铁蛋白基因RhFer1参与花瓣失水胁迫耐性和衰老进程的功能分析[D]. 北京: 中国农业大学, 2018. LIU Jitao. Functional Analysis of Rh Ferritinl Involved in Dehydration Tolerance and Senescence of Cut Rose Flowers[D]. Beijing: China Agricultural University, 2018. [6] 叶方婷, 潘鑫峰, 毛志君, 等. 睡莲转录因子bZIP家族的分子进化以及功能分析[J]. 中国农业科学, 2021, 54(21): 4694 − 4708. YE Fangting, PAN Xinfeng, MAO Zhijun, et al. Molecular evolution and function analysis of bZIP family in Nymphaea colorata [J]. Scientia Agricultura Sinica, 2021, 54(21): 4694 − 4708. [7] XU Yanjie, ZHAO Xin, PALINUER A, et al. A zinc finger protein BBX19 interacts with ABF3 to negatively affect drought tolerance in chrysanthemum [J]. Plant Journal, 2020, 103(5): 1783 − 1795. [8] 王英, 张超, 付建新, 等. 桂花花芽分化和花开放研究进展[J]. 浙江农林大学学报, 2016, 33(2): 340 − 347. WANG Ying, ZHANG Chao, FU Jianxin, et al. Progresses on flower bud differentiation and flower opening in Osmanthus fragrans [J]. Journal of Zhejiang A&F Univiversity, 2016, 33(2): 340 − 347. [9] MOU Wangshu, LI Dongdong, LUO Zisheng, et al. SlAREB1 transcriptional activation of NOR is involved in abscisic acid-modulated ethylene biosynthesis during tomato fruit ripening [J]. Plant Science, 2018, 276: 239 − 249. [10] CHEN Chengjie, CHEN Hao, ZHANG Yi, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data [J]. Molecular Plant, 2020, 13(8): 1194 − 1202. [11] 付建新, 张超, 王艺光, 等. 桂花组织基因表达中荧光定量PCR内参基因的筛选[J]. 浙江农林大学学报, 2016, 33(5): 727 − 733. FU Jianxin, ZHANG Chao, WANG Yiguang, et al. Reference gene selection for quantitative real-time polymerase chain reaction (qRT-PCR) normalization in the gene expression of sweet osmanthus tissues [J]. Journal of Zhejiang A&F Univiversity, 2016, 33(5): 727 − 733. [12] JAKOBY M, WEISSHAAR B, DRÖGE-LASER W, et al. bZIP transcription factors in Arabidopsis [J]. Trends in Plant Science, 2002, 7(3): 106 − 111. [13] JI Lexiang, WANG Jia, YE Meixia, et al. Identification and characterization of the populus AREB/ABF subfamily [J]. Journal of Integrative Plant Biology, 2013, 55(2): 177 − 186. [14] FUJITA Y, YOSHIDA T, YAMAGUCHI-SHINOZAKI K. Pivotal role of the AREB/ABF-SnRK2 pathway in ABRE-mediated transcription in response to osmotic stress in plants [J]. Physiologia Plantarum, 2013, 147(1): 15 − 27. [15] 杨玲, 吴玉乾, 谢晓东, 等. 烟草ABF转录因子基因的克隆与生物信息学分析[J]. 烟草科技, 2014(6): 73 − 81, 92. YANG Ling, WU Yuqian, XIE Xiaodong, et al. Clone and bioinformatics analysis of ABF transcription factor gene from Nicotiana tabacum [J]. Tobacco Science &Technology, 2014(6): 73 − 81, 92. [16] LI Fangfang, MEI Fangming, ZHANG Yifang, et al. Genome-wide analysis of the AREB/ABF gene lineage in land plants and functional analysis of TaABF3 in Arabidopsis [J/OL]. BMC Plant Biology, 2020, 20[2022-02-25]. doi: 10.1186/s12870-020-02783-9. [17] 杨玲. 烟草ABF转录因子基因的克隆及功能分析[D]. 重庆: 重庆大学, 2014. YANG Ling. Cloning and Functional Analysis of ABF Transcription Factor Gene in Nicotiana tabacum[D]. Chongqing: Chongqing University, 2014. [18] 刘贺, 黄沁梅, 刘颖婕, 等. 野菊bZIP转录因子全基因组鉴定及生物信息学分析[J]. 分子植物育种, 2022, 20(14): 4586 − 4600. LIU He, HUANG Qinmei, LIU Yingjie, et al. Genome-wide identification and bioinformatics analysis of Chrysanthemum indicum bZIP transcription factor family [J]. Molecular Plant Breeding, 2022, 20(14): 4586 − 4600. [19] 李天杰, 吴颖, 高龙飞, 等. 蓝莓ABF转录因子VcABF2基因的克隆与表达分析[J/OL]. 分子植物育种, 2021-11-20[2022-02-25]. http://kns.cnki.net/kcms/detail/46.1068.s.20211117.1020.004.html. LI Tianjie, WU Ying, GAO Longfei, et al. Cloning and expression analysis of ABF transcription factor gene VcABF2 in blueberry[J/OL]. Mol Plant Breed, 2021-11-20[2022-02-25]. http://kns.cnki.net/kcms/detail/46.1068.s.20211117.1020.004.html. [20] 林延慧, 唐力琼, 徐靖, 等. 大豆响应涝害bZIP基因Glyma04g04170的生物信息学分析及互作蛋白预测[J]. 大豆科学, 2020, 39(5): 728 − 731. LIN Yanhui, TANG Liqiong, XU Jing, et al. Bioinformatics analysis and interacting protein prediction of soybean bZIP gene glyma04g04170 in response to submergence stress [J]. Soybean Science, 2020, 39(5): 728 − 731. [21] 熊孟连, 戴星, 简燕, 等. 脱落酸依赖的与非依赖的信号途径的研究进展[J]. 基因组学与应用生物学, 2020, 39(12): 5796 − 5802. XIONG Menglian, DAI Xing, JIAN Yan, et al. Advances in the study of abscisic acid-dependent and non-dependent signaling pathways [J]. Genomics and Applied Biology, 2020, 39(12): 5796 − 5802. [22] ORELLANA S, YAÑEZ M, ESPINOZA A, et al. The transcription factor SlAREB1 confers drought, salt stress tolerance and regulates biotic and abiotic stress-related genes in tomato [J]. Plant,Cell &Environment, 2010, 33(12): 2191 − 2208. [23] BASTÍAS A, LÓPEZ-CLIMENT M, VALCÁRCEL M, et al. Modulation of organic acids and sugar content in tomato fruits by an abscisic acid-regulated transcription factor [J]. Physiologia Plantarum, 2011, 141(3): 215 − 226. [24] 胡鹏伟, 何朝勇, 洪岚, 等. AREB/ABF转录因子响应胁迫信号的网络调控[J]. 植物生理学报, 2013, 49(6): 540 − 544. HU Pengwei, HE Chaoyong, HONG Lan, et al. Internet regulation of AREB transcription factors responsed stress signal [J]. Plant Physiology Journal, 2013, 49(6): 540 − 544. [25] 涂明星. 葡萄转录因子VlbZIP30抗旱功能及其调控机理研究[D]. 杨凌: 西北农林科技大学, 2021. TU Mingxing. Drought Resistance Function and Regulation Mechanism Analysis of Grapevine Transcription Factor VlbZIP30 Gene[D]. Yangling: Northwest A&F University, 2021. [26] YANG Xiulian, YUE Yuanzheng, LI Haiyan, et al. The chromosome-level quality genome provides insights into the evolution of the biosynthesis genes for aroma compounds of Osmanthus fragrans[J/OL]. Horticulture Research, 2018, 5: 72[2022-02-25]. doi: 10.1038/s41438-018-0108-0. [27] LI Qi, ZHAO Haixia, WANG Xiaoli, et al. Tartary buckwheat transcription factor FtbZIP5, regulated by FtSnRK2.6, can improve salt/drought resistance in transgenic Arabidopsis[J/OL]. International Journal of Molecular Sciences, 2020, 21(3): 1123[2022-02-25]. doi: 10.3390/ijms21031123. [28] ZHAO Biyan, HU Yufeng, LI Juanjuan, et al. BnaABF2, a bZIP transcription factor from rapeseed (Brassica napus L.), enhances drought and salt tolerance in transgenic Arabidopsis[J/OL]. Botanical Studies, 2016, 57: 12[2022-02-25]. doi: 10.1186/s40529-016-0127-9. [29] MOU Wangshu, LI Dongdong, BU Jianwen, et al. Comprehensive analysis of ABA effects on ethylene biosynthesis and signaling during tomato fruit ripening[J]. PLoS One, 2016, 11(4): e0154072[2022-02-25]. doi: 10.1371/journal. pone.0154072. [30] 牟望舒. 脱落酸及脱落酸-乙烯互作调控番茄果实成熟的效应与机理[D]. 杭州: 浙江大学, 2019. MOU Wangshu. The Roles and Mechanism of Abscisic Acid and Abscisic Acid-ethylene Crosstalk in the Regulation of Tomato Fruit Ripening[D]. Hangzhou: Zhejiang University, 2019. [31] 魏明, 王含, 李成浩. 毛果杨PtAREB9基因启动子的克隆与功能初步分析[J]. 植物生理学报, 2015, 51(11): 1927 − 1932. WEI Ming, WANG Han, LI Chenghao, et al. Cloning and functional identification of promoter region of PtAREB9 from Populus trichocarpa [J]. Plant Physiology Journal, 2015, 51(11): 1927 − 1932. [32] 崔院院, 郭先锋, 邢树堂, 等. 脱落酸在牡丹切花衰老中的作用[J]. 中国农学通报, 2015, 31(22): 136 − 141. CUI Yuanyuan, GUO Xianfeng, XING Shutang, et al. Role of abscisic acid in senescence of tree peony cut rlower [J]. Chinese Agricultural Science Bulletin, 2015, 31(22): 136 − 141. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220264
 点击查看大图
点击查看大图

计量
- 文章访问数: 2144
- HTML全文浏览量: 410
- PDF下载量: 88
- 被引次数: 0
