






-
类胡萝卜素是一种亲脂类异戊二烯,是一种自然界中广泛存在的生物色素,主要反射黄色、橙色和红色的光[1]。在植物器官中,类胡萝卜素在质体中的积累对吸收过量光能、清除活性氧、合成植物激素提供前体物质都起重要的作用[2]。八氢番茄红素合成酶(phytoene synthase, PSY)是类胡萝卜素合成途径中的第1个限速酶,它的作用是将2分子的香叶酰香叶酰二磷酸(geranylgeranyl diphosphate,GGPP)合成无色的八氢番茄红素[3]。随后,八氢番茄红素经过包括八氢番茄红素脱氢酶(phytoene desaturase,PDS)在内的4次去饱和反应和2次异构化反应的多顺式转化,最终生成红色的全反式番茄红素[4]。而β-羟化酶(β-carotene hydroxylase,HYB)既可以单独作用使β-胡萝卜素经羟化作用转化为紫黄质,也可与ε-羟化酶(ε-carotene hydroxylase,HYE)协同作用使α-胡萝卜素经羟化作用形成叶黄素[5]。桂花Osmanthus fragrans是重要的观赏植物,花色和花香是其主要观赏性状。已有研究发现:类胡萝卜素既是桂花花瓣中主要色素成分[6],也是桂花香气物质的前体物质[7]。目前已在多种植物中发现,植物器官中的类胡萝卜素含量及相关基因的表达水平受到多种因素的影响,如温度[8]、光照[9]、干旱胁迫[10]、乙烯[11]等。前期研究[12]发现:在200 mg·L−1脱落酸处理下,桂花花色明显加深,花瓣类胡萝卜素含量上升,类胡萝卜素合成关键基因OfPSY、OfPDS和OfHYB表达水平显著上调。相关基因在前人的研究中均已克隆到[13-15],但对其调控的作用机制仍知之甚少。基因的启动子作为上游调控因子识别并结合的部位,是基因表达调控的重要作用位点。为进一步揭示桂花花色形成及其调控的分子机制,本研究克隆了OfPSY、OfPDS和OfHYB基因的启动子,通过作用元件分析、表达载体构建和瞬时表达分析,初步明确其作用。
-
8~10年生丹桂品种‘堰虹桂’Osmanthus fragrans‘Yanhong Gui’栽植于浙江农林大学桂花资源圃;烟草Nicotiana benthamiana栽培于浙江农林大学园林植物实验室。
-
DNA提取试剂盒、Premix Taq聚合酶、质粒载体PMD18-T、大肠埃希菌Escherichia coli DH5α、切胶回收试剂盒、DNA片段纯化试剂盒、限制性内切酶EcoR Ⅰ、Nco Ⅰ、DNA连接酶等购自Takara公司(大连)。
-
参照DNA提取试剂盒所用方法提取桂花‘堰虹桂’基因组DNA。
-
根据桂花基因组数据[16]中的OfPSY、OfPDS、OfHYB基因启动子序列,用Primer Premier 5.0分别设计上下游引物。引物由有康生物公司(杭州)合成(表1)。
表 1 启动子克隆所用引物
Table 1. Primers used for promoter cloning
引物名称 序列(5′→3′) OfPDSP-F TTAAATGGACGACTCATGTAATA OfPDSP-R CTCAAATTAACAGCAGAAACAT OfPSYP-F AAGCTTCAAAATTGCTGCTCAACTCATAC OfPSYP-R TCTAGAGCTGATACTGAACTATTAACGGTC OfHYBP-F AAGCTTTGGGTCTTACCTAACATCTTGGC OfHYBP-R TCTAGAGGACGGTAGTTTCAAGGGGGTG -
以桂花‘堰虹桂’基因组DNA为模板,分别用引物OfPSYP-F和OfPSYP-R、OfPDSP-F和OfPDSP-R、OfHYBP-F和OfHYBP-R对其启动子进行扩增。将扩增产物连接至质粒载体PMD18-T并转化大肠埃希菌DH5α,随后鉴定阳性克隆并送至有康公司(杭州)测序。启动子作用元件分析通过在线网站Plant CARE (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )进行。 -
设计包含酶切位点的引物(表2),用于构建启动子表达载体。使用限制性内切酶EcoR Ⅰ和Nco Ⅰ分别对PCAMBIA3301-LUC载体和添加了酶切位点的启动子进行双酶切。用T4连接酶连接回收的启动子和载体片段。再经转化大肠埃希菌DH5α鉴定,得到重组的PSYP::LUC、PDSP::LUC、HYBP::LUC载体。再将重组质粒转化农杆菌Agrobacterium tumefaciens GV3101。
表 2 构建PCAMBIA3301-LUC载体所用引物
Table 2. Primers used to construct PCAMBIA3301-LUC vector
引物名称 序列(5′→3′) OfPDSP-F GGTACCTTAAATGGACGACTCATGTAATA OfPDSP-R CCATGGCTCAAATTAACAGCAGAAACAT OfPSYP-F GGTACCAAGCTTCAAAATTGCTGCTCAACTCATAC OfPSYP-R CCATGGTCTAGAGCTGATACTGAACTATTAACGGTC OfHYBP-F GGTACCAAGCTTTGGGTCTTACCTAACATCTTGGC OfHYBP-R CCATGGTCTAGAGGACGGTAGTTTCAAGGGGGTG -
将含有重组质粒的农杆菌菌液在含有利福平和卡那霉素的LB培养基中振荡培养至D(600)达0.8~1.0,在4 ℃下以4 000 r·min−1离心10 min,收集菌体,随后以2 mL含10 mmol·L−1MES、10 mmol·L−1 MgCl2和150 μmol·L−1的乙酰丁香酮的悬浮液重悬菌体2次。选取长势较好的烟草,将悬浮液用1 mL注射器从叶片下表皮注射到烟草叶片中直至整个叶片呈现水渍状。将烟草置于暗处培养1 d后转移至人工气候室继续培养1~2 d,随后将植株分别进行37 ℃处理和200 mg·L−1脱落酸喷施处理,12 h后将注射后的叶片取下,喷洒1 mmol·L−1的荧光素钠盐溶液,暗处放置5 min后在CCD冷冻发光仪下观察LUC荧光信号。参考Luciferase (Promega)荧光素酶报告系统试剂盒对酶活性进行检测,以对照组的比值为单位1,得到不同处理组的相对Luciferase活性,每组实验均包括3次技术重复和生物学重复。利用SPSS 19.0软件进行数据差异分析。
-
以桂花‘堰虹桂’基因组DNA为模板,用引物OfPSYP-F和OfPSYP-R、OfPDSP-F和OfPDSP-R、OfHYBP-F和OfHYBP-R对其启动子进行扩增,分别得到OfPSY启动子长度为1 908 bp (图1),OfPDS启动子长度为1521 bp (图2),OfHYB启动子长度为1 830 bp (图3)的序列。利用Plant CARE在线软件对启动子序列的结合位点进行分析。在OfPSYP中,存在TATA-box、CAAT-box等启动子基本元件,和光响应元件、脱落酸(ABA)响应元件、赤霉素响应元件等响应元件,以及MYB、MYC结合位点(表3);在OfPDSP中,存在TATA-box、CAAT-box等启动子基本元件,和脱落酸响应元件、茉莉酸甲酯响应元件、赤霉素响应元件、光响应元件、厌氧诱导型元件、防御和胁迫响应元件等响应元件,以及MYB、MYC结合位点(表4);在OfHYBP中,存在TATA-box、CAAT-box等启动子基本元件,和脱落酸响应元件、生长素响应元件、低温响应元件、乙烯响应元件、茉莉酸甲酯响应元件、光响应元件、厌氧诱导型元件等响应元件,以及MYB、MYC结合位点(表5)。

图 1 OfPSY基因的启动子序列
Figure 1. Promoter sequence of OfPSY gene

图 2 OfPDS基因的启动子序列
Figure 2. Promoter sequence of OfPDS gene

图 3 OfHYB基因的启动子序列
Figure 3. Promoter sequence of OfHYB gene
表 3 OfPSY启动子中的顺式作用元件
Table 3. Cis-acting elements in OfPSY promoters
元件名称 数量 序列(5′→3′) 功能 AAGAA-motif 1 GTAAAGAAA ABRE 1 ACGTG 脱落酸响应元件 Box 4 1 ATTAAT 参与光响应的部分保守DNA序列 CAAT-box 20 CAAT 一般元件 G-box 1 TACGTG 光响应元件 GARE-motif 1 TCTGTTG 赤霉素响应元件 GT1-motif 1 GGTTAAT 光响应元件 MYB 1 TAACCA MYB 结合位点 MYC 1 CATGTG MYC 结合位点 P-box 1 CCTTTTG 赤霉素响应元件 STRE 1 AGGGG TATA-box 14 TATA 一般元件 TCT-motif 1 TCTTAC 光响应元件的一部分 Unnamed_1 1 CGTGG 表 4 OfPDS启动子中的顺式作用元件
Table 4. Cis-acting elements in OfPDS promoters
元件名称 数量 序列(5′→3′) 功能 ARE 3 AAACCA 厌氧诱导相关的顺式调节元件 AT~TATA-box 1 TATATA 一般元件 Box 4 1 ATTAAT 参与光响应的部分保守
DNA序列CAAT-box 21 CAAT 一般元件 CCGTCC motif 1 CCGTCC CGTCA-motif 1 CGTCA 茉莉酸甲酯响应元件 G-box 1 TACGTG 光响应元件 GA-motif 1 ATAGATAA 光响应元件 MBS 2 CAACTG MYB 结合位点 MYB 1 TAACCA MYB 结合位点 MYC 1 CATGTG MYC 结合位点 P-box 2 CCTTTTG 赤霉素响应元件 TATA-box 9 TATA 一般元件 TC-rich repeats 1 ATTCTCTAAC 参与防御和胁迫的元件 TCT-motif 3 TCTTAC 光响应元件的一部分 Unnamed_1 1 CGTGG Unnamed_4 4 CTCC WRE3 1 CCACCT 表 5 OfHYB启动子中的顺式作用元件
Table 5. Cis-acting elements in OfHYB promoters
元件名称 数量 序列(5′→3′) 功能 元件名称 数量 序列(5′→3′) 功能 ARE 2 AAACCA 厌氧诱导相关的顺式调节元件 GT1-motif 2 GGTTAA 光响应元件 A-box 1 CCGTCC 顺势调控元件 Gap-box 1 CAAATGAA 光响应元件的一部分 ABRE 4 ACGTG 脱落酸响应元件 LTR 1 CCGAAA 低温响应元件 AE-box 1 AGAAACTT 光响应元件的一部分 MYC 1 CATGTG MYC结合位点 AT~TATA-box 1 TATATA 一般元件 Myb 1 TAACTG MYB结合位点 AuxRR-core 1 GGTCCAT 生长素响应元件 TATA 1 TATAAAAT 一般元件 CAAT-box 20 CAAT 一般元件 TATA-box 6 TATA 一般元件 CCGTCC motif 1 CCGTCC TCCC-motif 1 TCTCCCT 光响应元件的一部分 CGTCA-motif 1 CGTCA 茉莉酸甲酯响应元件 TCT-motif 2 TCTTAC 光响应元件的一部分 ERE 1 ATTTTAAA 乙烯响应元件 Unnamed_1 1 GAATTTAATTAA 60K蛋白质结合位点 G-box 2 TACGTG 光响应元件的一部分 Unnamed_4 9 CTCC G-Box 1 CACGTT 光响应元件的一部分 WRE3 1 CCACCT GC-motif 1 CCCCCG 参与缺氧特异性诱导的元件 -
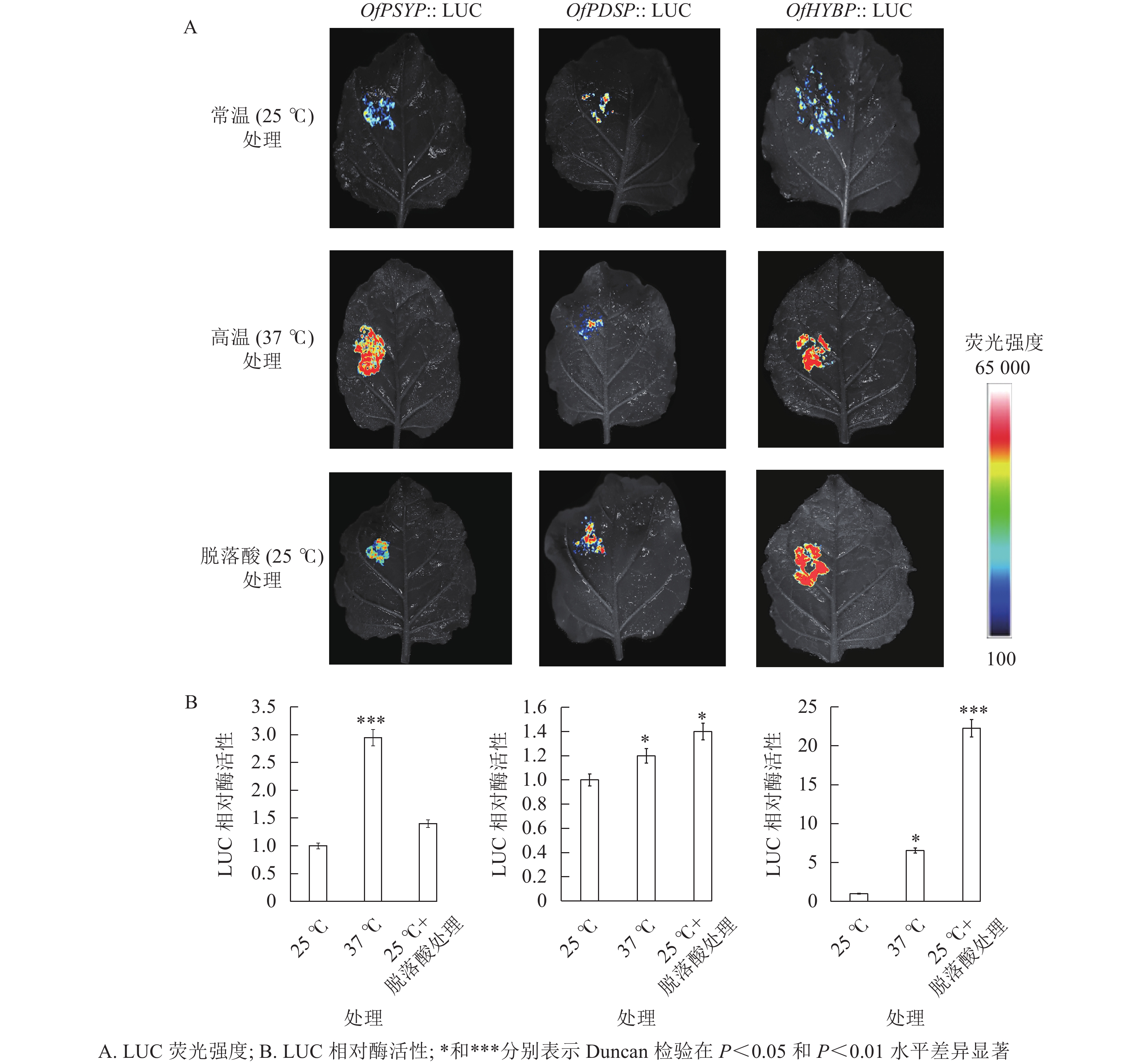
重组载体瞬时表达的荧光成像结果显示:注射了含重组载体农杆菌的烟草叶片均显现出荧光(图4),表明OfPSYP、OfPDSP、OfHYBP均能够驱动LUC报告基因的表达,具有启动子活性。相对于25 ℃处理的烟草叶片,在37 ℃处理下,注射了OfPSYP::LUC、OfPDSP::LUC和OfHYBP::LUC的烟草叶片呈现出更强的荧光信号;在200 mg·L−1脱落酸处理下,注射了OfPDSP::LUC和OfHYBP::LUC的烟草叶片呈现出更强的荧光信号。结果表明:相对高温胁迫上调了OfPSY、OfPDS和OfHYB的启动子活性,脱落酸上调了OfPDS和OfHYB的启动子活性。
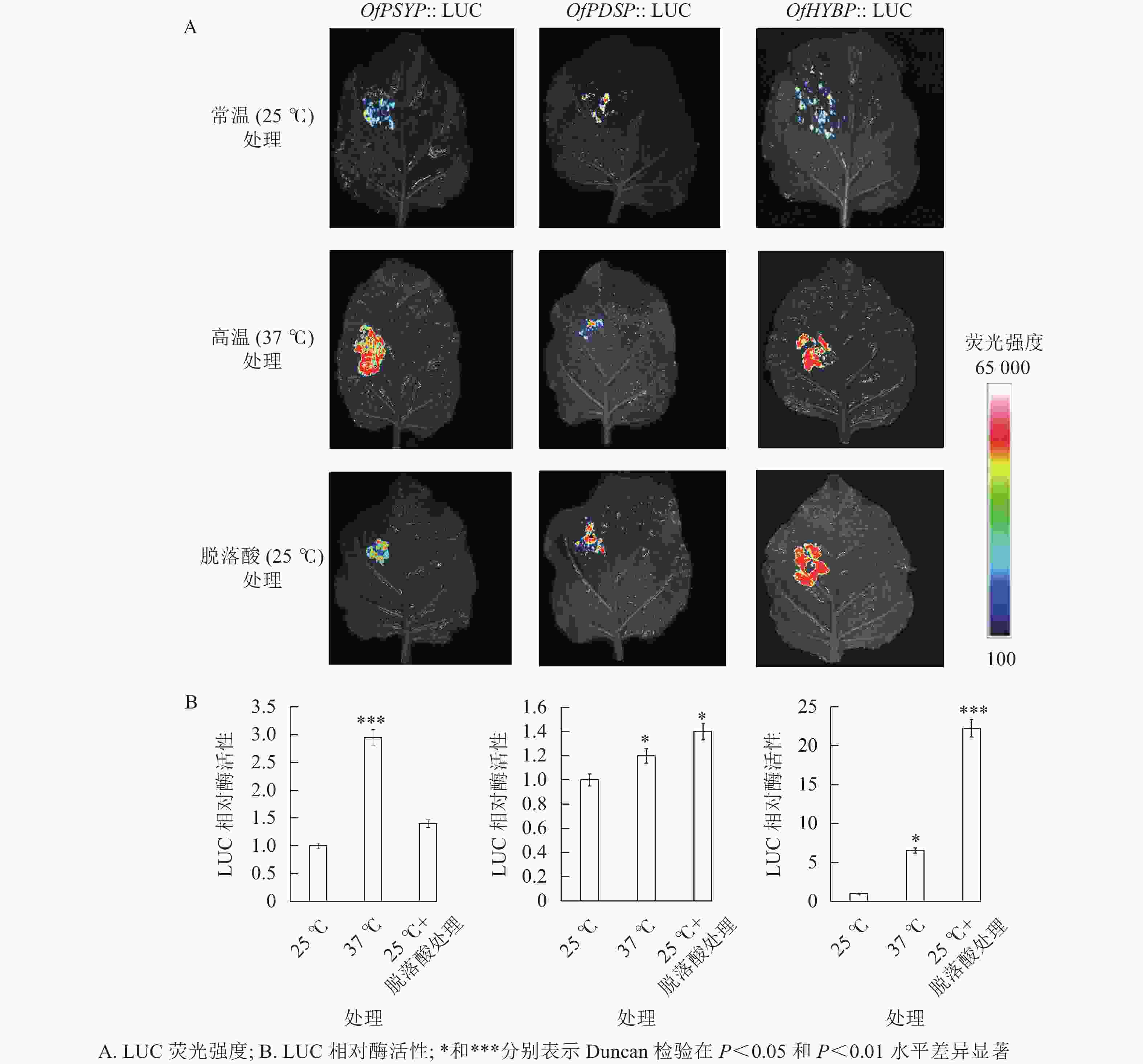
图 4 瞬时表达分析OfPSY、OfPDS和OfHYB启动子表达特性
Figure 4. Transient expression analysis of OfPSY, OfPDS and OfHYB promoter expression characteristics
-
植物中类胡萝卜素的成分和含量是由一系列酶促反应完成的。研究发现类胡萝卜素代谢关键基因的表达受到各种环境因素和激素的调控。在香蕉Musa nana中,高温可以上调α-胡萝卜素和β-胡萝卜素生物合成途径相关基因的转录水平[17];在蓝莓Vaccinium spp.中,红光和远红光对果实中类胡萝卜素合成和降解基因的表达均起到上调作用[18];在大豆Glycine max中,氯化钠(NaCl)、聚乙二醇(PEG)、高温、低温等胁迫和ABA处理对类胡萝卜素降解基因起明显的上调作用[19]。此外,转录因子对类胡萝卜素代谢关键基因的启动子存在直接调控作用。在柑橘Citrus reticulata中,CsMADS6基因可以结合CsPSY和CsPDS基因的启动子[20],从而促进其基因的表达;在番木瓜Carica papaya中,CpEIN3a既可以直接识别并结合CpPDS和CpCHYB基因的启动子,也可与CpNAC1/2基因共同促进CpPDS的表达[21],而CpSBP1则对CpPDS基因存在负调控作用[22];在胡萝卜Daucus carota中,DcAREB3可响应盐胁迫和ABA处理,识别并结合DcPSY2启动子的ABRE作用元件,从而促进其表达[23]。
前期通过对桂花进行ABA处理发现,相对于未处理的桂花,经200 mg·L−1 ABA处理的桂花花瓣中的类胡萝卜素含量上升;对花瓣中类胡萝卜素代谢关键基因实时荧光定量表达显示:经ABA处理后OfPSY、OfPDS、OfHYB等基因的表达量显著上调,推测ABA通过调控这几个基因的表达,从而影响了桂花花色[12]。本研究在OfHYB启动子中发现了4个ABRE作用元件,该作用元件被认为是AREB转录因子的结合位点[24]。研究发现AREB转录因子能够识别并结合2个相距较近的ABRE作用元件[25]。在OfHYB启动子的4个ABRE作用元件中,有3个元件之间相距19和12 bp,表明OfHYB基因极有可能受到ABA调控,与此同时,OfHYB启动子上发现了最多的激素响应元件,除ABA响应元件外,还存在生长素、茉莉酸甲酯和乙烯响应元件,表明OfHYB基因的表达可能受到多种激素的调控。本研究通过高温和外源施加ABA,研究了几个启动子表达特性,进一步验证了相对高温对OfPSY和OfHYB基因启动子以及ABA对OfPDS和OfHYB启动子表达的调控作用。此外,在3个基因启动子中均存在数个光响应元件,在桂花中已经发现OfCCD1的表达可能受光照影响[26−28],说明桂花花色物质的合成与降解均可能与光信号传导有关,但其具体作用机制仍有待进一步研究。本研究克隆得到的启动子经瞬时表达验证均具有启动子活性,下一步可将其构建酵母单杂载体,通过寻找上游的调控因子,明确桂花花瓣类胡萝卜素合成基因转录调控的分子机制。
Cloning and expression characterization of OfPSY, OfPDS and OfHYB gene promoters in Osmanthus fragrans
-
摘要:
目的 探究高温和外源脱落酸对桂花Osmanthus fragrans类胡萝卜素生物合成的3个相关基因,包括八氢番茄红素合成酶基因(PSY)、八氢番茄红素脱氢酶基因(PDS)、β-胡萝卜素羟化酶基因(HYB)的调控作用,为阐释桂花类胡萝卜素代谢调控的机制提供研究基础。 方法 根据桂花基因组数据库的序列,从桂花品种‘堰虹桂’‘Yanhong Gui’中克隆OfPSY、OfPDS、OfHYB基因的启动子序列,并进行生物信息学分析,再构建PCAMBIA3301-LUC载体在烟草Nicotiana benthamiana中瞬时表达,结合高温(37 ℃)和200 mg·L−1脱落酸处理,分析启动子活性。 结果 获得OfPSY、OfPDS、OfHYB基因的部分启动子,其长度分别为1 908、1 521及1 830 bp。作用元件分析表明:3个启动子中均存在TATA-box和CAAT-box等启动子基本元件、光响应元件、脱落酸响应元件以及MYB和MYC结合位点。此外,在OfPSY启动子中,存在赤霉素响应元件;在OfPDS启动子中,存在茉莉酸甲酯响应元件、赤霉素响应元件、厌氧诱导型元件参与防御和胁迫的元件;在OfHYB启动子中,存在生长素、乙烯、茉莉酸甲酯等激素响应元件、低温响应元件和厌氧诱导型元件。烟草瞬时转化试验表明:相对高温能激活OfPSY、OfPDS和OfHYB的启动子活性,脱落酸能激活OfPDS和OfHYB的启动子活性。 结论 高温和脱落酸可能通过调控桂花类胡萝卜素合成基因启动子活性,影响桂花类胡萝卜素的积累。图4表5参28 Abstract:Objective This paper aims to investigate the regulation of high temperature and abscisic acid (ABA) on promoters of three carotenoid biosynthesis genes in Osmanthus fragrans including PSY, PDS, HYB, providing the research foundation for regulation of carotenoid biosynthesis in O. fragrans. Method According to the sequences in genome database of O. fragrans, promoters of OfPSY, OfPDS, OfHYB were cloned from an O. fragrans cultivar ‘Yanhong Gui’, which were used for bioinformatics analysis. Then, these promoters were inserted into PCAMBIA3301-LUC vectors and used for transient expression in tobacco(Nicotiana benthamiana) leaves under treatments of high temperature (37 ℃) and 200 mg·L−1ABA. Result Our research cloned the promoter of OfPSY, OfPDS and OfHYB from genomic database of O. fragrans in length as 1908 bp, 1521 bp and 1830 bp respectively. All three promoters contain promoter basic elements TATA-box and CAAT-box, light response element, ABA response element as well as the binding site of MYB and MYC. Moreover, there was gibberellins (GA) response element in OfPSYP; OfPDSP contains methyl jasmonate (MeJA) and GA response element, pathogen induction element as well as element involved in defense and stress responsiveness; OfHYBP contains hormone response elements as auxin, ethylene and MeJA, as well as low temperature response element and pathogen induction element. The assays of in transient expression in tobacco leaves showed that relative high temperature can activate the promoter activities of OfPSY, OfPDS and OfHYB, and ABA can activate the promoter activities of OfPDS and OfHYB. Conclusion High temperature and ABA probably affect the carotenoid accumulation in O. fragrans by regulating the promoter activities of carotenoid biosynthesis genes. [Ch, 4 fig. 5 tab. 28 ref.] -
Key words:
- Osmanthus fragrans /
- carotenoids /
- promoters /
- temperature /
- abscisic acid
-
图 4 瞬时表达分析OfPSY、OfPDS和OfHYB启动子表达特性
Figure 4 Transient expression analysis of OfPSY, OfPDS and OfHYB promoter expression characteristics
表 1 启动子克隆所用引物
Table 1. Primers used for promoter cloning
引物名称 序列(5′→3′) OfPDSP-F TTAAATGGACGACTCATGTAATA OfPDSP-R CTCAAATTAACAGCAGAAACAT OfPSYP-F AAGCTTCAAAATTGCTGCTCAACTCATAC OfPSYP-R TCTAGAGCTGATACTGAACTATTAACGGTC OfHYBP-F AAGCTTTGGGTCTTACCTAACATCTTGGC OfHYBP-R TCTAGAGGACGGTAGTTTCAAGGGGGTG  下载: 导出CSV
下载: 导出CSV
表 2 构建PCAMBIA3301-LUC载体所用引物
Table 2. Primers used to construct PCAMBIA3301-LUC vector
引物名称 序列(5′→3′) OfPDSP-F GGTACCTTAAATGGACGACTCATGTAATA OfPDSP-R CCATGGCTCAAATTAACAGCAGAAACAT OfPSYP-F GGTACCAAGCTTCAAAATTGCTGCTCAACTCATAC OfPSYP-R CCATGGTCTAGAGCTGATACTGAACTATTAACGGTC OfHYBP-F GGTACCAAGCTTTGGGTCTTACCTAACATCTTGGC OfHYBP-R CCATGGTCTAGAGGACGGTAGTTTCAAGGGGGTG
下载: 导出CSV
表 3 OfPSY启动子中的顺式作用元件
Table 3. Cis-acting elements in OfPSY promoters
元件名称 数量 序列(5′→3′) 功能 AAGAA-motif 1 GTAAAGAAA ABRE 1 ACGTG 脱落酸响应元件 Box 4 1 ATTAAT 参与光响应的部分保守DNA序列 CAAT-box 20 CAAT 一般元件 G-box 1 TACGTG 光响应元件 GARE-motif 1 TCTGTTG 赤霉素响应元件 GT1-motif 1 GGTTAAT 光响应元件 MYB 1 TAACCA MYB 结合位点 MYC 1 CATGTG MYC 结合位点 P-box 1 CCTTTTG 赤霉素响应元件 STRE 1 AGGGG TATA-box 14 TATA 一般元件 TCT-motif 1 TCTTAC 光响应元件的一部分 Unnamed_1 1 CGTGG
下载: 导出CSV
表 4 OfPDS启动子中的顺式作用元件
Table 4. Cis-acting elements in OfPDS promoters
元件名称 数量 序列(5′→3′) 功能 ARE 3 AAACCA 厌氧诱导相关的顺式调节元件 AT~TATA-box 1 TATATA 一般元件 Box 4 1 ATTAAT 参与光响应的部分保守
DNA序列CAAT-box 21 CAAT 一般元件 CCGTCC motif 1 CCGTCC CGTCA-motif 1 CGTCA 茉莉酸甲酯响应元件 G-box 1 TACGTG 光响应元件 GA-motif 1 ATAGATAA 光响应元件 MBS 2 CAACTG MYB 结合位点 MYB 1 TAACCA MYB 结合位点 MYC 1 CATGTG MYC 结合位点 P-box 2 CCTTTTG 赤霉素响应元件 TATA-box 9 TATA 一般元件 TC-rich repeats 1 ATTCTCTAAC 参与防御和胁迫的元件 TCT-motif 3 TCTTAC 光响应元件的一部分 Unnamed_1 1 CGTGG Unnamed_4 4 CTCC WRE3 1 CCACCT
下载: 导出CSV
表 5 OfHYB启动子中的顺式作用元件
Table 5. Cis-acting elements in OfHYB promoters
元件名称 数量 序列(5′→3′) 功能 元件名称 数量 序列(5′→3′) 功能 ARE 2 AAACCA 厌氧诱导相关的顺式调节元件 GT1-motif 2 GGTTAA 光响应元件 A-box 1 CCGTCC 顺势调控元件 Gap-box 1 CAAATGAA 光响应元件的一部分 ABRE 4 ACGTG 脱落酸响应元件 LTR 1 CCGAAA 低温响应元件 AE-box 1 AGAAACTT 光响应元件的一部分 MYC 1 CATGTG MYC结合位点 AT~TATA-box 1 TATATA 一般元件 Myb 1 TAACTG MYB结合位点 AuxRR-core 1 GGTCCAT 生长素响应元件 TATA 1 TATAAAAT 一般元件 CAAT-box 20 CAAT 一般元件 TATA-box 6 TATA 一般元件 CCGTCC motif 1 CCGTCC TCCC-motif 1 TCTCCCT 光响应元件的一部分 CGTCA-motif 1 CGTCA 茉莉酸甲酯响应元件 TCT-motif 2 TCTTAC 光响应元件的一部分 ERE 1 ATTTTAAA 乙烯响应元件 Unnamed_1 1 GAATTTAATTAA 60K蛋白质结合位点 G-box 2 TACGTG 光响应元件的一部分 Unnamed_4 9 CTCC G-Box 1 CACGTT 光响应元件的一部分 WRE3 1 CCACCT GC-motif 1 CCCCCG 参与缺氧特异性诱导的元件
下载: 导出CSV
-
[1] MCQUINN R P, GIOVANNONI J J, POGSON B J. More than meets the eye: from carotenoid biosynthesis, to new insights into apocarotenoid signaling [J]. Current Opinion in Plant Biology, 2015, 27: 172 − 179. [2] NISAR N, LI Li, LU Shan, et al. Carotenoid metabolism in plants [J]. Molecular Plant, 2015, 8(1): 68 − 82. [3] HAN Y, LI L, DONG M, et al. cDNA cloning of the phytoene synthase (PSY) and expression analysis of PSY and carotenoid cleavage dioxygenase genes in Osmanthus fragrans [J]. Biologia, 2013, 68(2): 258 − 263. [4] MCQUINN R P, WONG B, GIOVANNONI J J. AtPDS overexpression in tomato: exposing unique patterns of carotenoid self-regulation and an alternative strategy for the enhancement of fruit carotenoid content [J]. Plant Biotechnology Journal, 2017, 16(2): 482 − 494. [5] DU Hao, WANG Nili, CUI Fei, et al. Characterization of a β-carotene hydroxylase gene DSM2 conferring drought and oxidative stress resistance by increasing xanthophylls and ABA synthesis in rice [J]. Plant Physiology, 2010, 154(3): 1304 − 1318. [6] WANG Yiguang, ZHANG Chao, DONG Bin, et al. Carotenoid accumulation and its contribution to flower coloration of Osmanthus fragrans[J/OL]. Frontiers in Plant Science, 2018: 1499[2022-01-04]. doi: 10.3389/fpls.2018.01499. [7] HAN Yuanji, WANG Hongyun, WANG Xiaodan, et al. Mechanism of floral scent production in Osmanthus fragrans and the production and regulation of its key floral constituents, β-ionone and linalool [J]. Horticulture Research, 2019, 6(1): 432 − 443. [8] 侯丹, 付建新, 张超, 等. 桂花品种‘堰虹桂’‘玉玲珑’和‘杭州黄’的香气成分及释放节律[J]. 浙江农林大学学报, 2015, 32(2): 208 − 220. HOU Dan, FU Jianxin, ZHANG Chao, et al. Flower scent composition of Osmanthus fragrans ‘Yanhong Gui’‘Yu Linglong’ and ‘Hangzhou Huang’, and their emission patterns [J]. Journal of Zhejiang A&F University, 2015, 32(2): 208 − 220. [9] HE Yuan, MA Yafeng, DU Yu, et al. Differential gene expression for carotenoid biosynthesis in a green alga Ulva prolifera based on transcriptome analysis [J]. BMC Genomics, 2018, 19(1): 916 − 930. [10] ZHANG Lancui, MA Gang, KATO M, et al. Regulation of carotenoid accumulation and the expression of carotenoid metabolic genes in citrus juice sacs in vitro [J]. Journal of Experimental Botany, 2012(2): 871 − 886. [11] 范敏, 金黎平, 黄三文, 等. 干旱胁迫对马铃薯类黄酮和类胡萝卜素合成关键酶基因表达的影响[J]. 园艺学报, 2008, 35(4): 535 − 542. FAN Min, JIN Liping, HUANG Sanwen, et al. Effects of drought stress on gene expression of key enzymes for flavonoid and carotenoid synthesis in potato [J]. Acta Horticulturae Sinica, 2008, 35(4): 535 − 542. [12] LIU Yudong, SHI Yuan, SU Deding, et al. SlGRAS4 accelerates fruit ripening by regulating ethylene biosynthesis genes and SlMADS1 in tomato[J/OL]. Horticulture Research, 2021, 8(3)[ 2022-01-02]. doi: 10.1038/S41438-020-00431-9. [13] LIU Yudong, DONG Bin, ZHANG Chao, et al. Effects of exogenous abscisic acid (ABA) on carotenoids and petal color in Osmanthus fragrans ‘Yanhonggui’ [J]. Plants, 2020, 9(4): 454 − 466. [14] HAN Y, WANG X, CHEN W, et al. Differential expression of carotenoid-related genes determines diversified carotenoid coloration in flower petal of Osmanthus fragrans [J]. Trees Genetics &Genomes, 2014, 10(2): 329 − 338. [15] BALDERMANN S, KATO M, FLEISCHMANN P, WATANABE N. Biosynthesis of α-and β-ionone, prominent scent compounds, in flowers of Osmanthus fragrans [J]. Acta Biochimica Polonica, 2012, 59(1): 79 − 81. [16] YANG Xiulian, YUE Yuanzheng, LI Haiyan, et al. The chromosome-level quality genome provides insights into the evolution of the biosynthesis genes for aroma compounds of Osmanthus fragrans [J/OL]. Horticulture Research, 2018, 5: 72[2021-12-23]. doi: 10.1038/s41438-018-0108-0. [17] FU Xiumin, CHENG Sihua, FENG Chao, et al. Lycopene cyclases determine high α-/β-carotene ratio and increased carotenoids in bananas ripening at high temperatures [J]. Food Chemistry, 2019, 283: 131 − 140. [18] KARPPINEN K, ZORATTI L, SARALA M, et al. Carotenoid metabolism during bilberry (Vaccinium myrtillus L. ) fruit development under different light conditions is regulated by biosynthesis and degradation [J]. BMC Plant Biology, 2016, 16(1): 95 − 111. [19] WANG Ruikai, WANG Chun’e. Genome-wide identification and transcription analysis of soybean carotenoid oxygenase genes during abiotic stress treatments [J]. Molecular Biology Reports, 2013, 40(8): 4737 − 4745. [20] LU Suwen, ZHANG Yin, ZHU Kaijie, et al. The citrus transcription factor CsMADS6 modulates carotenoid metabolism by directly regulating carotenogenic genes [J]. Plant Physiology, 2018, 176(4): 2657 − 2676. [21] FU Changchun, HAN Yanchao, KUANG Jianfei, et al. Papaya CpEIN3a and CpNAC2 co-operatively regulate carotenoid biosynthesis-related genes CpPDS2/4, CpLCY-e and CpCHY-b during fruit ripening [J]. Plant and Cell Physiology, 2017, 58(12): 2155 − 2165. [22] HAN Yanchao, GAO Haiyan, CHEN Hangjun, et al. The involvement of papaya CpSBP1 in modulating fruit softening and carotenoid accumulation by repressing CpPME1/2 and CpPDS4[J/OL]. Scientia Horticulturae, 2019, 256: 108582[2022-01-02]. doi:10.1016/j.scienta.2019.108582. [23] KEVIN S, PAULINA F, FELIPE Q I L, et al. Unraveling the induction of phytoene synthase 2 expression by salt stress and abscisic acid in Daucus carota [J]. Journal of Experimental Botany, 2018, 69(16): 4113 − 4126. [24] YOSHIDA T, FUJITA Y, SAYAMA H, et al. AREB1, AREB2, and ABF3 are master transcription factors that cooperatively regulate ABRE-dependent ABA signaling involved in drought stress tolerance and require ABA for full activation [J]. The Plant Journal, 2010, 61(4): 672 − 685. [25] CHOI H I, HONG J H, HA J O, et al. ABFs, a family of ABA-responsive element binding factors [J]. Journal of Biological Chemistry, 2000, 275(3): 1723 − 1730. [26] BALDERMANN S, KATO M, KUROSAWA M, et al. Functional characterization of a carotenoid cleavage dioxygenase 1 and its relation to the carotenoid accumulation and volatile emission during the floral development of Osmanthus fragrans Lour. [J]. Journal of Experimental Botany, 2010, 61(11): 2967 − 2977. [27] ZHANG Chao, WANG Yiguang, FU Jianxin, et al. Transcriptomic analysis and carotenogenic gene expression related to petal coloration in Osmanthus fragrans ‘Yanhong Gui’ [J]. Trees, 2016, 30(4): 1207 − 1223. [28] 刘玉成, 王艺光, 张超, 等. 桂花OfCCD1基因启动子克隆与表达特性[J]. 浙江农林大学学报, 2018, 35(4): 596 − 603. LIU Yucheng, WANG Yiguang, ZHANG Chao, et al. Cloning and transient expression assay of OfCCD1 gene promoters from Osmanthus fragrans [J]. Journal of Zhejiang A&F University, 2018, 35(4): 596 − 603. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220110
 点击查看大图
点击查看大图

计量
- 文章访问数: 2353
- HTML全文浏览量: 546
- PDF下载量: 203
- 被引次数: 0
