






-
开花是被子植物生活史上的一个重要阶段,是大多数有花植物性成熟的标志。植物的花开放物候体现了花开放的季节性和周期性规律,是植物应对环境变化的策略,也是开花生殖重要的组成部分,可以为进一步研究植物的形态结构、生理生态特性以及进行杂交育种提供依据[1]。然而植物花开放受多种因素影响,形成了一个复杂的调控网络,调控机制尚未清楚。目前,花开放由内部和外部因素共同影响。外部因素包括光、温度、相对湿度、气压以及生物因素等[2],内部因素主要是激素变化、细胞壁扩张、生物钟节律、水通道蛋白合成和碳水化合物代谢等途径[3−5]。许多植物激素与植物的成花诱导和开花结实关系紧密,如柑橘Citrus reticulata花瓣中脱落酸(ABA)含量随开花时间的延长而增加,高含量赤霉素(GA3)延迟番茄Solanum lycopersicum[6]开花,较低含量使番茄开花和花序发育较早。花瓣上下表皮细胞的生长差异、膨胀与收缩,影响着花朵开闭[7]。此外,花朵开放需要消耗大量的能量,碳水化合物作为主要的能源物质,其含量对花开放进程有很大的影响。
紫薇Lagerstroemia indica是桃金娘目Myratles千屈菜科Lythraceae紫薇属Lagerstroemia的落叶灌木或小乔木[8]。原产中国长江流域及华南各省,现各地广为栽培。紫薇花色鲜艳美丽,有紫色、红色、粉色、白色等多种花色,具有很高的观赏价值,其花期在夏季,花期长、枝干光滑、抗污染,是园林绿化的好材料。目前,关于紫薇花开放的研究多在开花生物学、形态学、繁育特性等方面[9−13],紫薇花开放生理层面的研究尚未见详细报道。本研究对紫薇开花期物候进行了观测,研究了紫薇花开放过程中细胞形态、细胞相对含水量、碳水化合物质量分数和内源激素等的变化,以揭示紫薇花开放的生理机制,有利于优化紫薇栽培措施,提高其观赏价值和经济效益,对紫薇花期调控以及花开放相关分子机制研究有重要意义。
-
选取栽植在浙江农林大学校园(119°72′69″N,30°25′65″E)的10株冠幅、株高、树龄较为一致的紫薇,植株生长状况良好且无病虫害。在2022年7月上旬时进行观察并采样,根据花开放物候观测,采集花开放各个时期的花以及花部结构。将样品于液氮中速冻,保存在−80 ℃超低温冰箱中备用。设置3次生物学重复。
-
于2022年7月上旬上午8:00开始观测紫薇植株,依据紫薇单花开放时期,分阶段观察并记录花朵开放程度,进行拍照记录并描述开放进程的形态特点,记录各时期持续时间。
-
根据紫薇花开放的时期,选取每个时期的花瓣进行扫描电镜观察,观察上下表皮细胞的形态[14]。于晴天上午8:00剪取花枝插入水中,及时带回实验室进行解剖,剥除萼片、雌蕊、雄蕊等花器官,留存花瓣,洗净擦干后,剪成0.5 cm × 0.5 cm大小,进行固定及脱水。将经过梯度脱水处理的样品放置于Hitachi HCP-2型临界点干燥仪中进行干燥处理,并采用离子溅射镀膜法进行镀膜。最后将镀过膜、喷过金的样品放置在Hitachi SU-
8010 型扫描电镜中进行观察。 -
利用烘干称量法[15]测定。分别称取0.1 g紫薇不同花开放时期的花瓣、萼片及花苞/花的鲜质量,洗净后放入烧杯中,蒸馏水浸泡8 h,擦干后称取水饱和质量,放入烘箱80 ℃烘干至恒量,得到干质量,求相对含水量。以上样品均进行3次生物学重复。
-
称取0.1 g花瓣样品,按照试剂盒说明书操作,分别在520、480、620、620 nm 波长下测定葡萄糖、果糖、蔗糖和淀粉的吸光度。用不同浓度的葡萄糖标准溶液测吸光度,绘制标准曲线,求出标准直线方程。以上样品均进行 3 次生物学重复。
蒽酮、葡萄糖标准品均购于浙江卡尔生物技术有限公司,试剂盒均购于苏州格锐思生物科技有限公司。淀粉采用了蒽酮比色法进行测定,葡萄糖则基于GOPOD 氧化酶法测定,果糖运用间苯二酚比色法测定,而蔗糖则利用蒽酮比色法进行检测。
-
称取1.0 g样品,用ACQUITY UPLC H-Class系统(沃特世)测定紫薇花开放各时期ABA、生长素(IAA)、GA3和玉米素核苷(ZR)的质量分数[15−17]。各样品重复测定3次。
-
使用Excel 2016进行数据统计,SPSS 25进行数据分析,采用皮尔逊(Pearson)检测法分析各指标之间的相关性,用Origin 9.1作图。
-
紫薇单花花开放可分为紧蕾期(S1)、花蕾膨大期(S2)、初开期(S3)和盛开期(S4)等4个时期(图1)。其中,S1时期花蕾紧闭,花蕾通体呈绿色,花瓣尚未显色,为白色偏淡绿色;S2时期花瓣开始显色,呈淡紫色,花蕾上部从绿色变为红色;S3时期花瓣逐渐裂开,花瓣及萼片逐渐伸展;S4时期时柱头伸长,花瓣及萼片完全伸展,花药开裂,柱头呈绿色并具黏液。
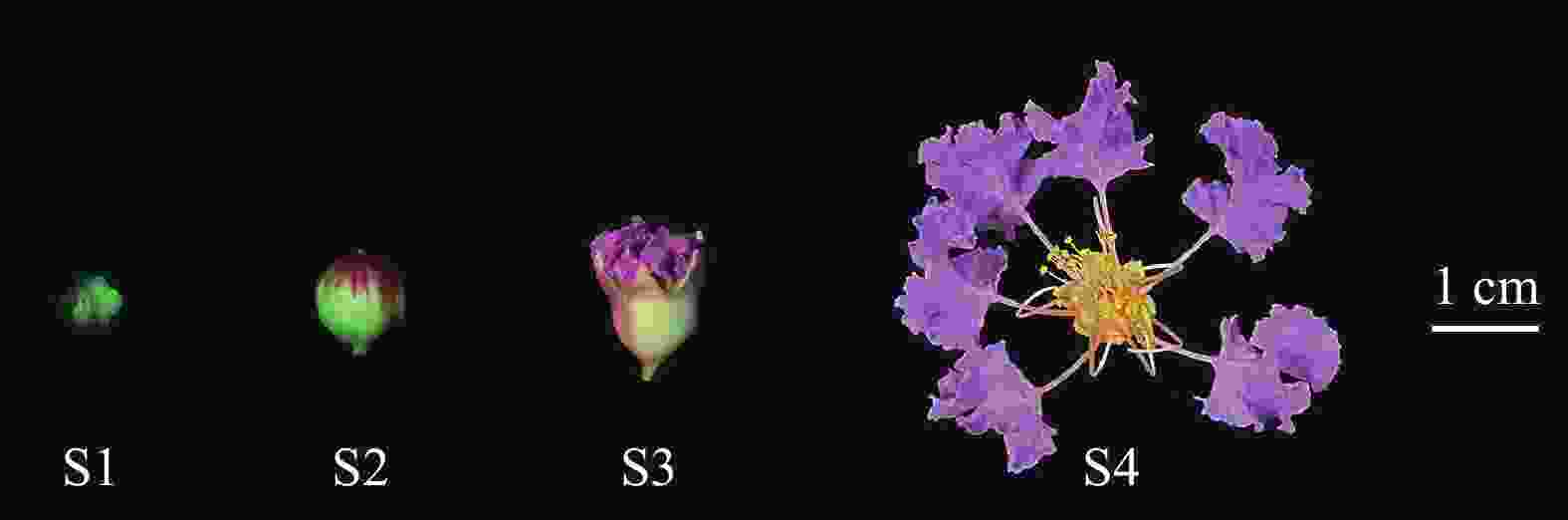
图 1 紫薇的单花开放进程
Figure 1. Flower opening process of L. indica single flower
紫薇一般在凌晨开放,气温越高,开裂时间提前;但遇阴雨天气时,开裂时间延后,甚至不开裂,花瓣停止生长,导致开花数量减少。紫薇在23:00时,花蕾膨大至最大,花瓣开始蓬松变大;到23:15时,花蕾顶部裂开一条缝,此时花瓣迅速膨大变松直至花蕾完全裂开;翌日0:30时,被花萼紧包的花瓣逐渐向外膨胀,爪基部外露;直至6:00时,花萼完全舒展,花瓣相互分离最终完全平展,花丝向四周伸展,雄蕊群接触面积增大,花药开裂,开始散粉,柱头透明绿色,开始分泌黏液。开花第3天,花丝干枯呈褐色,花瓣边缘开始萎蔫,颜色变淡,柱头黄绿色,干燥萎缩,无黏液。开花4~6 d,花瓣凋谢。
-
如图2所示:在S1时期,紫薇花瓣上下表皮细胞轮廓明显且顺滑,呈不规则多边形,排列较紧密,细胞表面光滑,下表皮细胞较上表皮细胞稍大。在S2时期,上下表皮细胞逐渐变大、密集饱满,形状为不规则长方形,细胞间排列紧密,细胞表面不光滑布满条纹状纹饰,下表皮细胞边缘呈锯齿状,较上表皮细胞稍大,且布满褶皱。在S3时期,花瓣上下表皮细胞间隙较上一时期大,细胞分布呈波状、条纹状、辐射状等纹饰,细胞不规则,形状各异。在S4时期,花瓣上下表皮细胞明显增大,且出现皱缩,细胞表面变得不平整;下表皮细胞也出现皱缩现象,但不如上表皮明显。
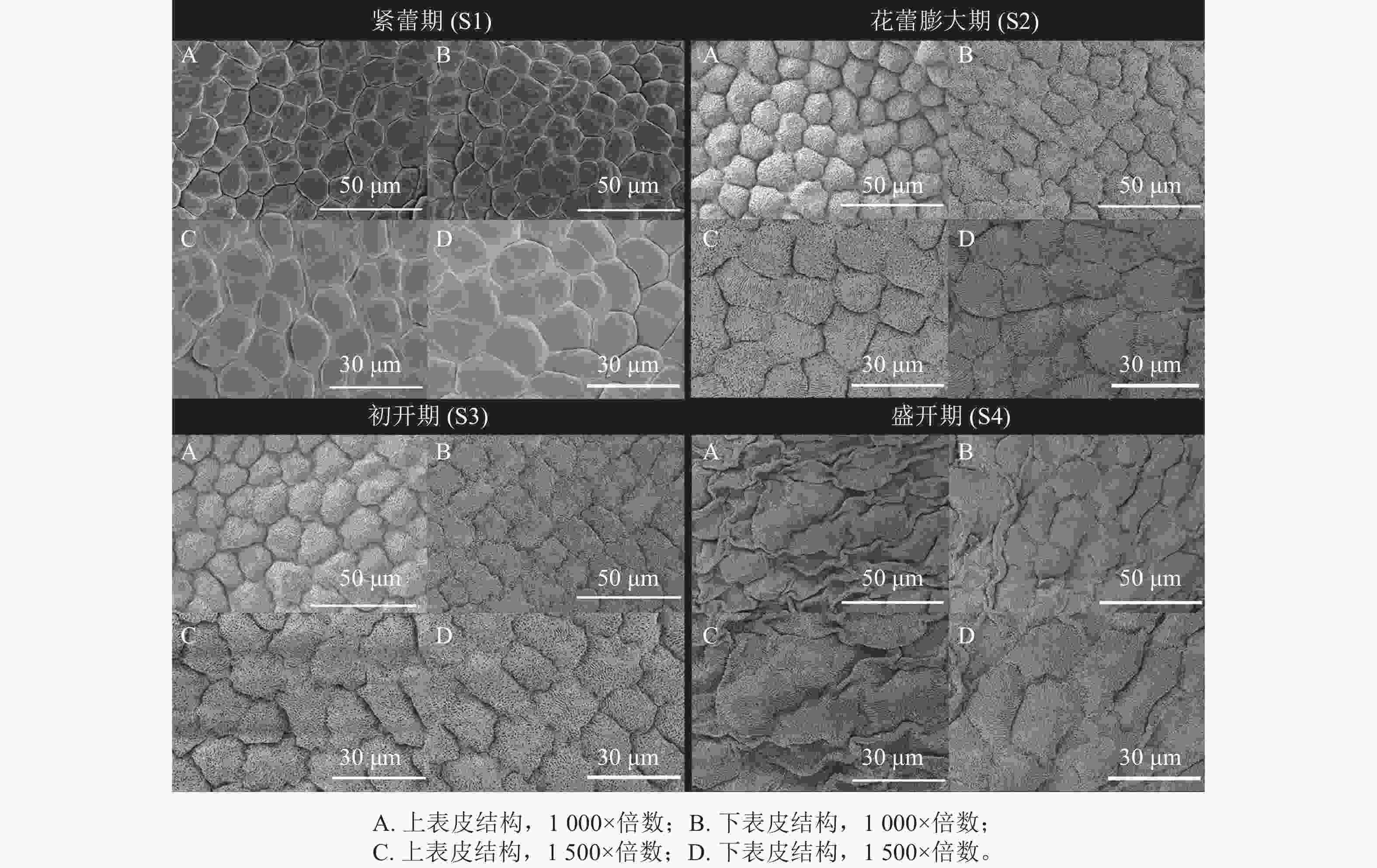
图 2 紫薇开花过程中的花瓣表皮结构变化
Figure 2. Changes in epidermal structure of the petals during L. indica flowering petals at different developmental stages
-
在紫薇花开放过程中,发现花瓣、萼片以及整个花苞/整朵花在紫薇花开放期间的相对含水量整体都呈上升趋势(图3),且都在S4时期达到最高,分别为72.97%、88.97%和88.92%。这表明在开花进程中,花部细胞需要具有较高的含水量以保持其紧张度维持正常开放。4个时期的萼片和整个花苞/整朵花相对含水量表现出显著差异(P<0.05),S1与S2时期花瓣无显著差异。
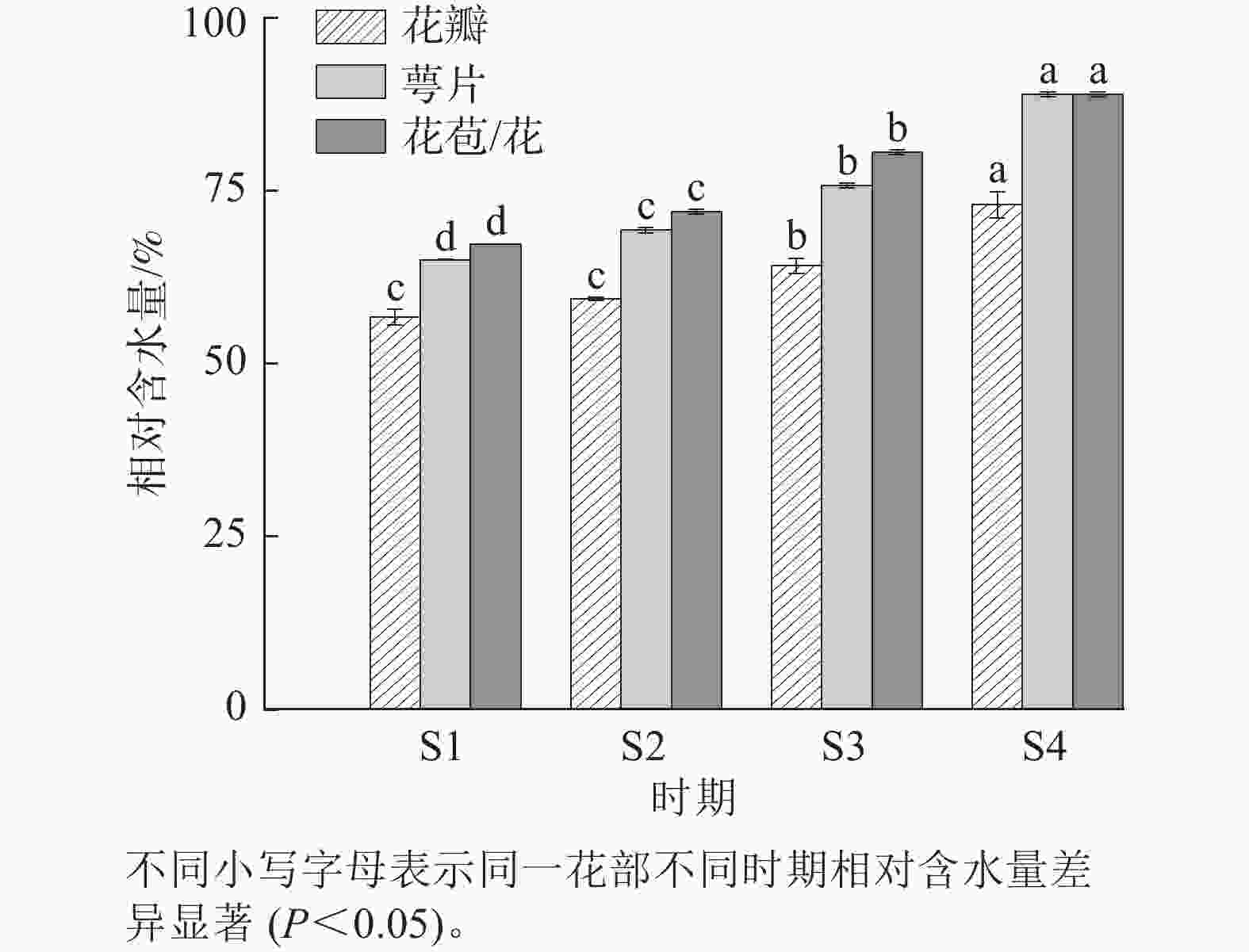
图 3 紫薇开花过程中花部相对含水量的变化
Figure 3. Changes in the relative water content of flowers during L. indica flowering
-
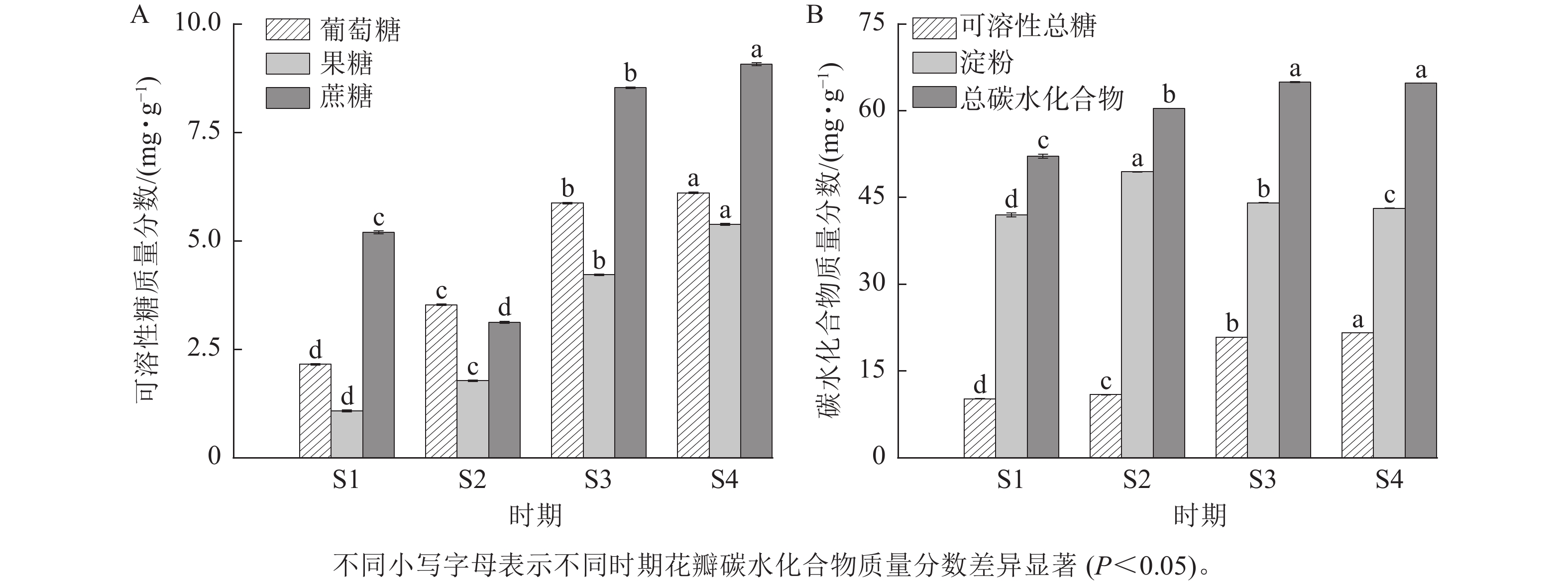
由图4可知:紫薇花开放过程中,葡萄糖、果糖和可溶性总糖质量分数具有类似的变化趋势,从S1到S4时期逐渐上升,其中果糖和可溶性总糖质量分数在S3时期上升幅度较大。紫薇花瓣中蔗糖质量分数呈先下降后上升的趋势,从S3时期开始增加,在S4时期达最大值,为9.07 mg·g−1。淀粉质量分数则呈先上升后下降的趋势,在S1时期质量分数最低,为41.97 mg·g−1。总碳水化合物质量分数在S3与S4时期差异不显著,葡萄糖、果糖、蔗糖、可溶性总糖和淀粉质量分数在4个时期的紫薇花瓣中表现出显著差异(P<0.05)。
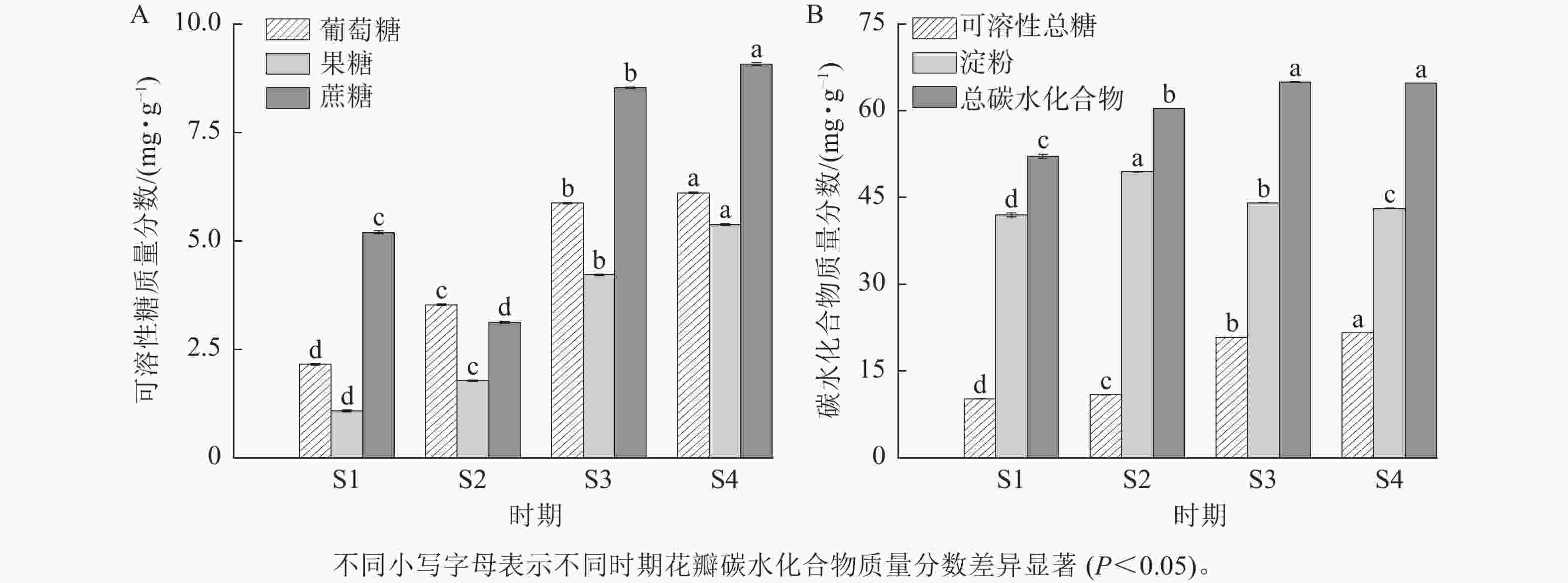
图 4 紫薇开花过程中花瓣碳水化合物质量分数的变化
Figure 4. Changes in carbohydrate content of L. indica petals during flowering
-
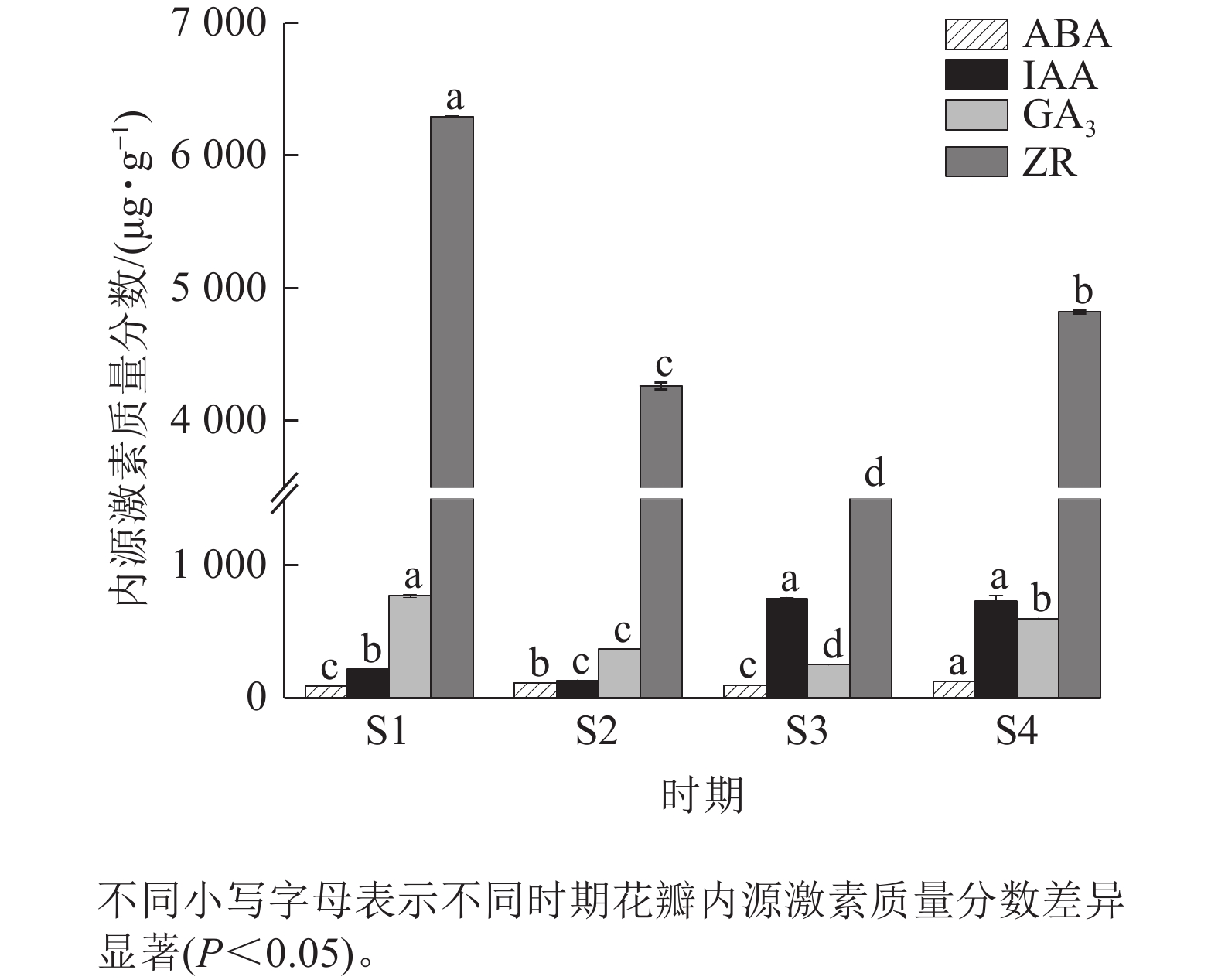
由图5可知:4个时期紫薇花瓣的ZR质量分数最高,ABA质量分数最低。ABA质量分数整体变化不明显,在紫薇开花期间呈“上升—下降—上升”趋势,在S4时期达到最高值,为123.45 μg·g−1。IAA质量分数呈“下降—上升—下降”趋势,在S3时期上升幅度较大,但S3时期与S4时期差异变化不显著。GA3与ZR质量分数变化趋势相似,呈先下降后上升趋势,均在S1时期最高,分别为770.65、6 294.81 μg·g−1。
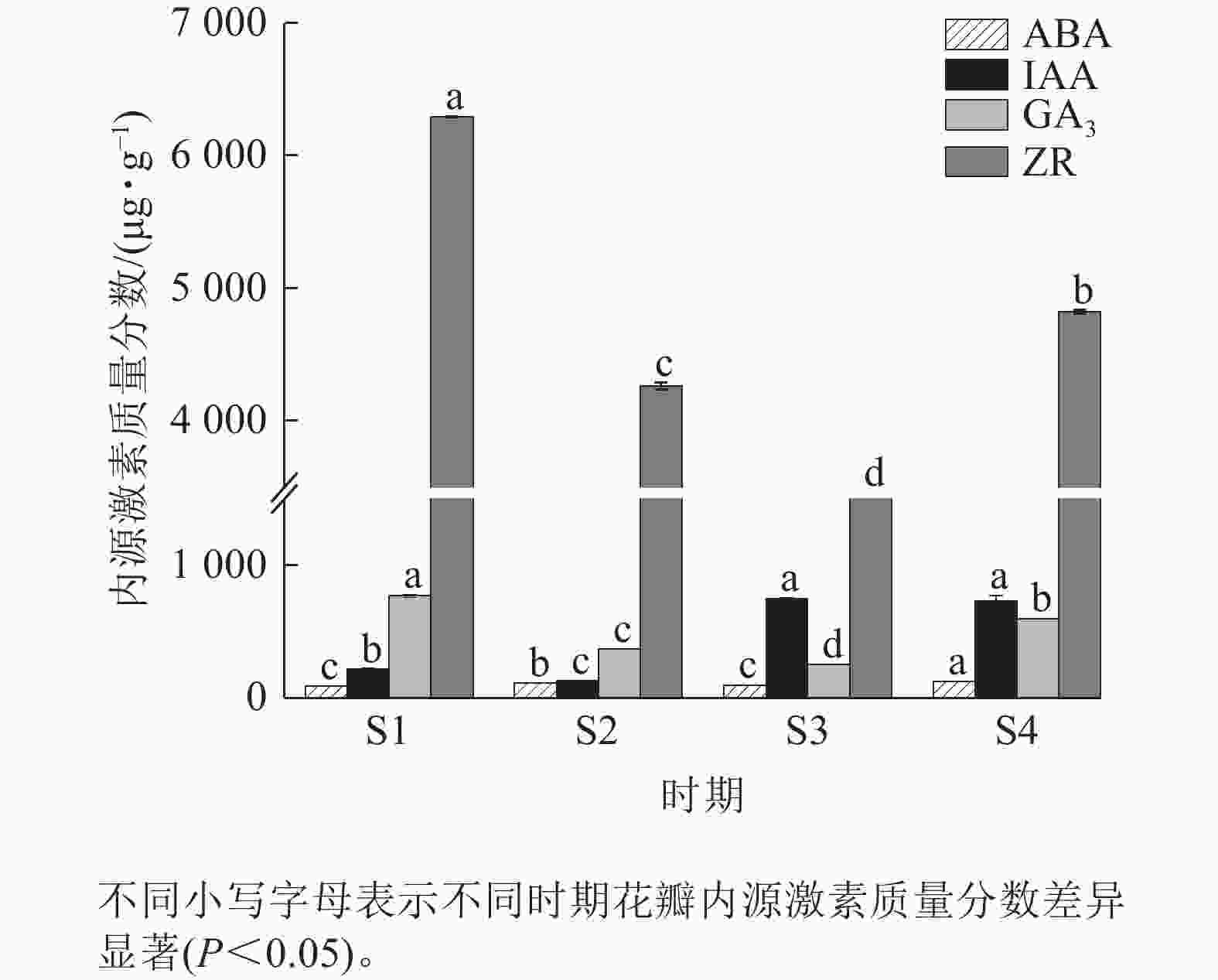
图 5 紫薇开花过程中花瓣的内源激素质量分数变化
Figure 5. Changes in endogenous hormone content in L. indica petals during flowering
如表1所示:从S1到S2时期,IAA/ABA逐渐降低,在S2时期降至最低,在S3时期迅速回升,在S4时期又回落。IAA/GA3在S1~S3时期逐渐升高,在S3时期最高,然后到S4时期有所下降。在紫薇花开放过程中,ZR/ABA先降低且降幅较大,在S3到S4时期略回升。ZR/GA3呈先升高后降低的趋势,在S3时期达到峰值,到S4时期又下降。ABA/GA3与ZR/GA3变化类似,均在S1~S3时期有所上升,然后在S4时期回落。
表 1 紫薇花开放过程中内源激素比值变化
Table 1. Changes of endogenous hormone ratio during L. indica flower opening
时期 IAA/ABA IAA/GA3 ZR/ABA ZR/GA3 ABA/GA3 S1 2.38 0.29 68.07 8.17 0.12 S2 1.17 0.36 37.90 11.61 0.31 S3 7.83 3.00 32.69 12.52 0.38 S4 5.92 1.22 39.05 8.06 0.31 -
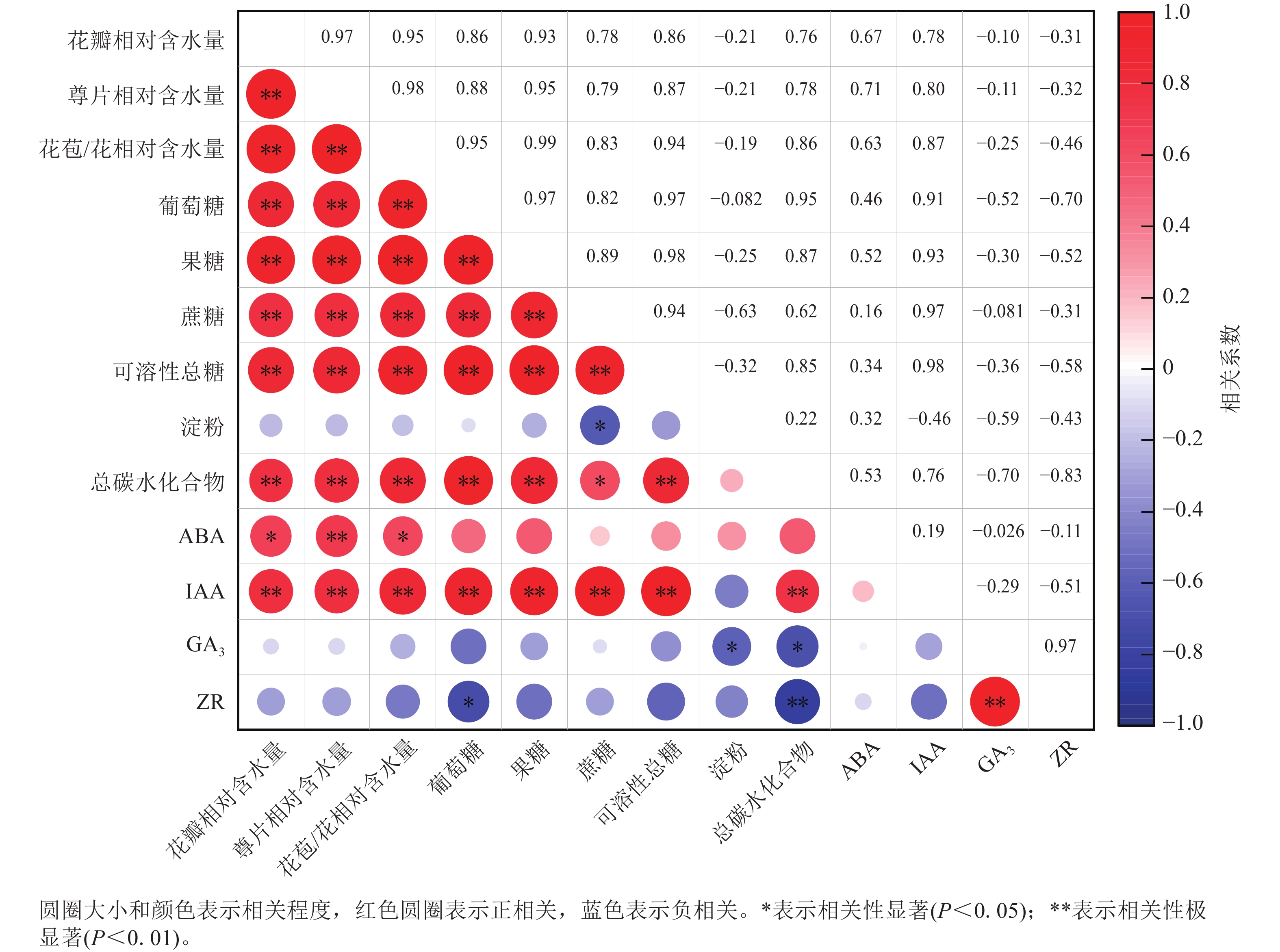
相关性分析(图6)显示:ZR与GA3质量分数呈极显著正相关(P<0.01),相关系数为0.97,与总碳水化合物质量分数呈极显著负相关(P<0.01),相关系数为−0.83。花瓣相对含水量与萼片相对含水量、花苞/花相对含水量、葡萄糖、果糖、蔗糖、可溶性总糖、总碳水化合物以及IAA质量分数呈极显著正相关(P<0.01),相关性系数分别为0.97、0.95、0.86、0.93、0.78、0.86、0.76和0.78,与ABA质量分数呈显著正相关(P<0.05),相关性系数为0.67。淀粉与蔗糖质量分数呈显著负相关(P<0.05),相关系数为−0.46。
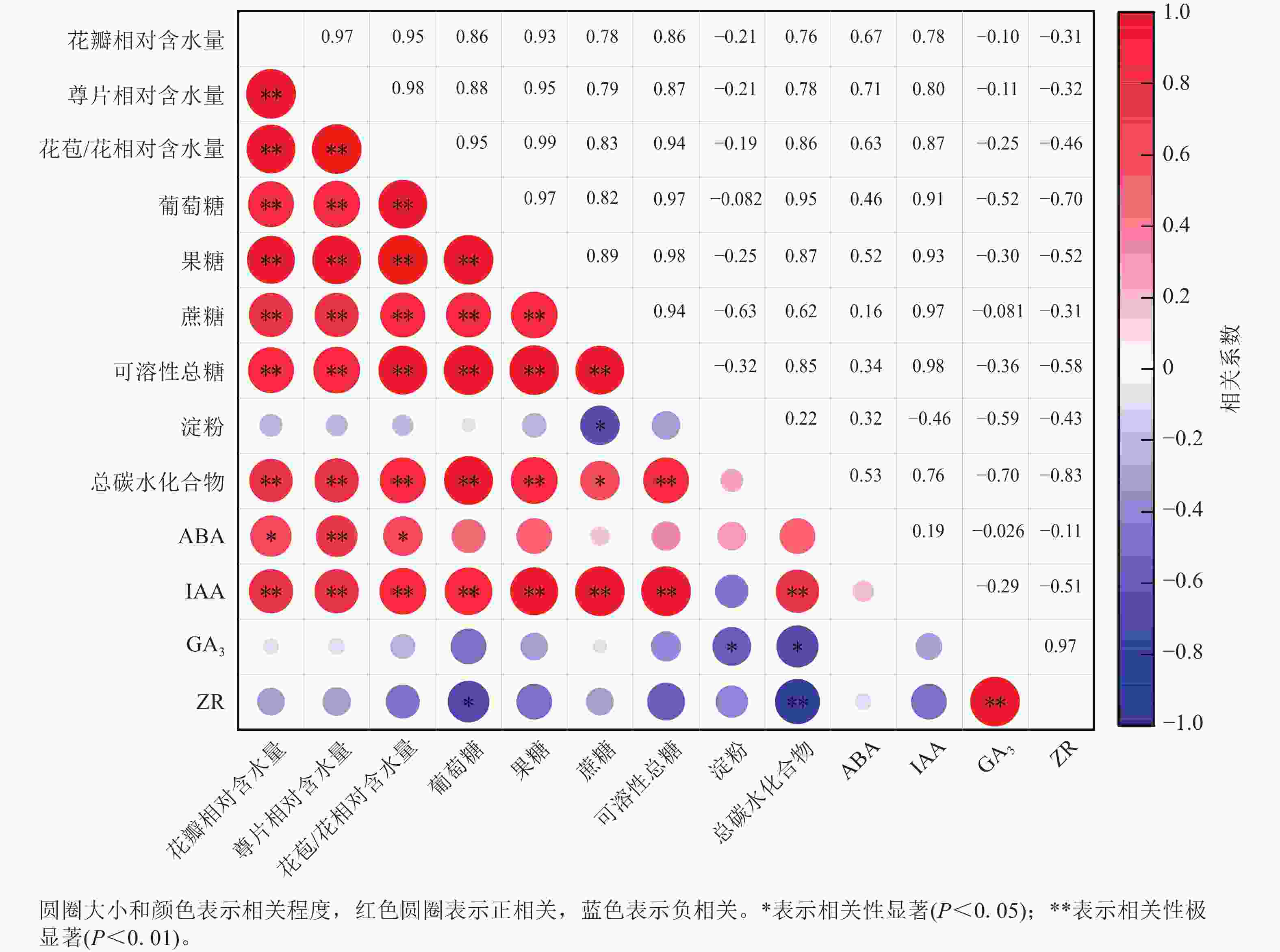
图 6 紫薇开花过程中花部细胞相对含水量、花瓣碳水化合物与内源激素的相关性
Figure 6. Correlation among cell relative water content, carbohydrates and endogenous hormones in L. indica petals during flowering
-
本研究将紫薇花开放过程分为紧蕾期(S1)、花蕾膨大期(S2)、初开期(S3)和盛开期(S4)等4个时期。利用扫描电镜对紫薇花开放4个时期花瓣上下表皮细胞进行观测,发现细胞形态在花开放过程中有显著变化。在花开放的各个时期,上表皮细胞数量略多于下表皮细胞,这与洋桔梗Eustoma grandiflorum[18]的研究结果一致。上表皮细胞的显著扩张和挤压现象可能增强了花瓣的颜色和质地,提高了对授粉者的吸引力,而细胞表面的不平整可能增加了花瓣的结构稳定性,有助于维持开放状态,便于授粉昆虫的访问[19]。
细胞水分在营养物质的运输以及渗透压的调节中起着关键作用。在开花进程中,含水量高有助于维持花瓣和萼片的紧张度和鲜艳度。相对含水量能够反映植物自身水分状况[20]。随着花开放的进行,紫薇各花部器官细胞的相对含水量均呈逐渐升高的趋势,梅花Armeniaca mume、蜡梅Chimonanthus praecox、江南槐Robinia pseudoacacia f. decaisneana和珍珠梅Sorbaria sorbifolia中也出现类似情况[21−23]。紫薇各花部器官细胞相对含水量增加有助于细胞保持一定的张力,能够促进紫薇花朵的开放。
碳水化合物是植物生长发育过程的重要结构物质和能量供给,其积累和消耗在很大程度上影响着花芽分化的基本进程[24]。碳水化合物主要指糖类物质,其中可溶性糖和可溶性淀粉是植物体内各项生命活动中起主要作用的糖类物质[25]。本研究中,紫薇开花过程中总碳水化合物呈逐渐上升趋势,推测总碳水化合物的积累有利于花芽发育。可溶性总糖作为植物能量的载体,是植物体内碳水化合物运输的主要形式,其质量分数变化能够反映植物体内的碳素营养状况[26]。淀粉是一种碳水化合物,已被证实是呼吸和代谢的中间产物,对植物开花有很大的影响[27−28]。许多物种开花初期花瓣细胞中含有大量淀粉,如百合Lilium spp.[29]和突厥蔷薇Rosa damascena[30]等,它们通过将淀粉转化为葡萄糖和果糖来促进花朵开放。在本研究中,淀粉呈“上升—下降”趋势,由此推测,在紫薇开花过程中淀粉可能转化为可溶性糖,以满足开花过程中代谢能量的需要。
植物激素在调控植物营养生长和生殖发育方面发挥着重要作用,与花芽发育和开花时间密切相关。本研究紫薇开花过程中,ABA总质量分数较少,推测较低水平的ABA有促进开花的作用。IAA质量分数与开花直接相关,在本研究中,IAA质量分数在紫薇开花前期先下降,在S2时期后急剧上升,到S3时期最高,这与日本杏Prunus armeniacal[31]的研究结果相似,推测高水平IAA可能有利于紫薇开花。在四季花金花茶Camellia perpetua[32]中,高水平的GA3和ZR有利于花芽分化,而低水平的GA3和ZR促使花开放。本研究中,GA3和ZR在紫薇开花过程中质量分数相对较高,S3时期最低,推测低水平的GA3和ZR促进紫薇开花。“激素平衡假说”认为:植物开花并非由单一激素决定,而是多种激素相互作用的结果[1]。本研究中随着花开放进行,花瓣中内源激素的比值也在变化。在紫薇花开放前期IAA/GA3高,盛开期内降低,ZR/GA3和ABA/GA3也呈现同样的趋势,所以这暗示了IAA、ZR、ABA与GA3的比值升高可能促进紫薇开花。ZR/ABA相对较高,在紫薇花开放前期比值一直呈下降趋势,推测ZR/ABA较低可以促进花朵绽放,到S3时期又上升,说明ZR/ABA在开花后上升延缓花瓣衰老,这与金花茶C. petelotii[33]的研究结果一致。此外,紫薇花瓣中IAA/ABA在S2到S3时期呈上升趋势,说明紫薇花瓣中IAA/ABA的升高可促进花朵绽放。
紫薇花开放过程中,花瓣相对含水量、萼片相对含水量、花苞/花相对含水量、葡萄糖、果糖、蔗糖、可溶性总糖、总碳水化合物以及IAA质量分数等9个指标呈极显著正相关,淀粉、GA3和ZR与大部分指标间的相关性不显著。
-
本研究基于物候观测,将紫薇单花开放过程分为4个时期,发现在紫薇花开放过程中花瓣上下表皮细胞逐渐变大,花部细胞相对含水量逐渐升高、碳水化合物质量分数和内源激素质量分数发生动态变化。未来可进一步探究内源激素的水平和比例、非生物胁迫等条件对紫薇花开放的影响和各器官内各物质的分配变化规律,以优化紫薇花开放进程,便于花期调控。
Physiological characteristics of Lagerstroemia indica flowers in opening process
-
摘要:
目的 研究紫薇Lagerstroemia indica花开放过程中的生理特征,为花期调控提供参考。 方法 以紫薇单花开放各时期的花瓣作为材料,测定细胞形态变化、细胞相对含水量、碳水化合物质量分数和内源激素质量分数,用皮尔逊(Pearson)检测法分析各指标之间的相关性。 结果 根据物候观测,紫薇单花开放可分为紧蕾期(S1)、花蕾膨大期(S2)、初开期(S3)和盛开期(S4)等4个时期。扫描电镜观察表明:随着花开放进程,花瓣上下表皮细胞逐渐扩张,且其细胞表面存在波状、条纹状、辐射状等纹饰,各时期下表皮细胞面积均大于上表皮面积。在紫薇花开放期间,花瓣、萼片以及整个花苞/整朵花的相对含水量整体呈上升趋势,而且可溶性总糖质量分数整体也呈上升趋势,淀粉质量分数呈先上升后下降的趋势。脱落酸(ABA)质量分数相对较低且变化较平稳,生长素(IAA)、赤霉素(GA3)质量分数较低且总体表达趋势相异,玉米素核苷(ZR)质量分数较高且变化幅度较大,ZR/ABA较大且变化幅度大。相关性分析显示:9个指标之间极显著正相关(P<0.01),淀粉与蔗糖质量分数显著负相关(P<0.05),与其他指标的相关性不显著。 结论 在紫薇花开放过程中,伴随着花瓣上下表皮细胞形态的显著变化、花部细胞相对含水量的增加、碳水化合物质量分数的上升以及内源激素质量分数的动态调整,这些变化共同促进了紫薇花的正常开放。图6表1参33 Abstract:Objective The study of physiological characteristics of Lagerstroemia indica flowers during the opening process provides reference for further floral regulation. Method This study took the petals of L. indica in different periods of flower opening as experimental materials, and investigated the physiological and biochemical changes, including cell morphology change, cell relative water content, carbohydrate content and endogenous hormone content. The correlations between the indicators were analyzed by Pearson detection method. Result According to the phenological observation, the single flower opening process can be divided into 4 periods: tight bud stage (S1), flower bud expansion stage (S2), early blooming stage (S3) and full blooming stage (S4). Scanning electron microscope observation showed that the upper and lower epidermal cells of the petals gradually expanded during flower opening, and there were undulating, striped and radial patterns on the cell surface with the progress of flower opening, and the area of the lower epidermal cells was larger than the area of the upper epidermis in each stage. The relative water content of petals, sepals and the whole bract/whole flower showed an overall increasing trend, the total soluble sugar content also showed an overall increasing trend, and the starch content showed an increasing and then decreasing trend. Abscisic acid (ABA) content was relatively low and changed steadily, the content of auxin (IAA) and gibberellin (GA3) was low and the overall expression trend was different, the content of zeatin riboside (ZR) was higher and the change was larger, and the ratio of ZR/ABA was larger and the change was large. Correlation analysis showed highly significant positive correlation (P<0.01) between the 9 indicators, significant negative correlation (P<0.05) between starch content and sucrose content, and non-significant correlation between starch content and other indicators. Conclusion During the opening process of L. indica, accompanied by significant changes in the morphology of the upper and lower epidermal cells of the petals, increase in relative water content of floral cells, rise in carbohydrate content and dynamic adjustment of endogenous hormone content, these changes jointly promoted the normal opening of L. indica flowers. [Ch, 6 fig. 1 tab. 33 ref.] -
Key words:
- Lagerstroemia indica /
- flower opening /
- physiological mechanisms /
- endogenous hormones
-
图 2 紫薇开花过程中的花瓣表皮结构变化
Figure 2 Changes in epidermal structure of the petals during L. indica flowering petals at different developmental stages
图 3 紫薇开花过程中花部相对含水量的变化
Figure 3 Changes in the relative water content of flowers during L. indica flowering
图 4 紫薇开花过程中花瓣碳水化合物质量分数的变化
Figure 4 Changes in carbohydrate content of L. indica petals during flowering
图 5 紫薇开花过程中花瓣的内源激素质量分数变化
Figure 5 Changes in endogenous hormone content in L. indica petals during flowering
图 6 紫薇开花过程中花部细胞相对含水量、花瓣碳水化合物与内源激素的相关性
Figure 6 Correlation among cell relative water content, carbohydrates and endogenous hormones in L. indica petals during flowering
表 1 紫薇花开放过程中内源激素比值变化
Table 1. Changes of endogenous hormone ratio during L. indica flower opening
时期 IAA/ABA IAA/GA3 ZR/ABA ZR/GA3 ABA/GA3 S1 2.38 0.29 68.07 8.17 0.12 S2 1.17 0.36 37.90 11.61 0.31 S3 7.83 3.00 32.69 12.52 0.38 S4 5.92 1.22 39.05 8.06 0.31  下载: 导出CSV
下载: 导出CSV
-
[1] 张姣姣, 行凯鑫, 赵顺宇, 等. 植物花朵开闭的调控机制[J]. 植物生理学报, 2023, 59(2): 292−302. ZHANG Jiaojiao, XING Kaixin, ZHAO Shunyu, et al. Mechanism of plant flower opening and closing [J]. Plant Physiology Journal, 2023, 59(2): 292−302. [2] HAMID M, HASSAN G I, WANI A W, et al. Physiology of flowering in apple and almond: a review [J]. International Journal of Current Microbiology and Applied Sciences, 2020, 9(9): 1912−1929. [3] 张吉顺, 张孝廉, 王仁刚, 等. 环境胁迫影响植物开花的分子机制[J]. 浙江大学学报(农业与生命科学版), 2016, 42(3): 289−305. ZHANG Jishun, ZHANG Xiaolian, WANG Rengang, et al. Molecular mechanism of the impact of environmental stress on plant flowering [J]. Journal of Zhejiang University (Agriculture & Life Sciences), 2016, 42(3): 289−305. [4] 林兴军, 马福生, 陈鹏, 等. 咖啡开花过程中叶片碳水化合物含量的变化动态[J]. 热带农业科学, 2019, 39(1): 5−9. LIN Xingjun, MA Fusheng, CHEN Peng, et al. Dynamic changes of carbohydrate content in coffee during flower bud differentiation [J]. Chinese Journal of Tropical Agriculture, 2019, 39(1): 5−9. [5] 韦虹日, 阳乾程, 覃永华, 等. 香木莲开花过程中叶片碳水化合物及营养元素的变化[J]. 中国农学通报, 2024, 40(1): 52−59. WEI Hongri, YANG Qiancheng, QIN Yonghua, et al. Research on changes of leaf carbohydrates and nutrients during the flowering process of Manglietia aromatica [J]. Chinese Agricultural Science Bulletin, 2024, 40(1): 52−59. [6] SILVA G F F, SILVA E M, CORREA J P O, et al. Tomato floral induction and flower development are orchestrated by the interplay between gibberellin and two unrelated microRNA-controlled modules [J]. New Phytologist, 2019, 221(3): 1328−1344. [7] van DOORN W G, KAMDEE C. Flower opening and closure: an update [J]. Journal of Experimental Botany, 2014, 65(20): 5749−5757. [8] 方文培. 中国植物志[M]. 北京: 科学出版社, 2016. FANG Wenpei. Flora of China [M]. Beijing: Science Press, 2016. [9] 王瑞文. 紫薇开花生物学特性及杂交育种的初步研究[D]. 武汉: 华中农业大学, 2010. WANG Ruiwen. Preliminary Study on Flowering Biological Characteristics and Cross Breeding of Lagerstroemia indica[D]. Wuhan: Huazhong Agricultural University, 2010. [10] 黄小珍. 紫薇花器官发育相关基因的克隆及表达特性分析[D]. 长沙: 中南林业科技大学, 2021. HUANG Xiaozhen. Cloning and Expression Analysis of Genes related to Organ Development of Osmunda japonica Thunb[D]. Changsha: Central South University of Forestry & Technology, 2021. [11] 罗雪梦, 王晓明, 曾慧杰, 等. 紫叶紫薇花芽分化及雌雄配子体发育的形态学研究[J]. 中南林业科技大学学报, 2022, 42(2): 129−138. LUO Xuemeng, WANG Xiaoming, ZENG Huijie, et al. Morphological study on floral bud differentiation and the development of female and male gametophytes in purple-leaf Lagerstroemia indica [J]. Journal of Central South University of Forestry & Technology, 2022, 42(2): 129−138. [12] 黄兰清, 吴丽君, 王晓明, 等. 配方施肥对‘紫精灵’紫薇容器苗生长、开花及生理的影响[J]. 植物生理学报, 2022, 58(9): 1735−1746. HUANG Lanqing, WU Lijun, WANG Xiaoming, et al. Effects of formula fertilization on growth, flowering and physiology of container seedlings of Lagerstroemia indica ‘Zi Jing Ling’ [J]. Plant Physiology Journal, 2022, 58(9): 1735−1746. [13] 孙阳, 袁娟, 郭萍, 等. 大花紫薇花部综合特征和繁育特性[J]. 东北林业大学学报, 2023, 51(3): 60−66. SUN Yang, YUAN Juan, GUO Ping, et al. Floral syndrome and breeding system of Lagerstroemia speciosa [J]. Journal of Northeast Forestry University, 2023, 51(3): 60−66. [14] 张健. 丁香属(Syringa)一些植物形态解剖与园林应用探讨[D]. 福州: 福建农林大学, 2009. ZHANG Jian. Study on Anatomical Characters and Landscape Application of Some Species in Syringa [D]. Fuzhou: Fujian Agriculturae and Forestry University, 2009. [15] 王学奎, 黄见良. 植物生理生化实验原理和技术[M]. 3版. 北京: 高等教育出版社, 2015. WANG Xuekui, HUANG Jianliang. Principles and Techniques of Plant Physiological Biochemical Experiment [M]. 3rd ed. Beijing: Higher Education Press, 2015. [16] 饶丹丹. 紫薇新品种‘紫玉’组织培养及内源激素含量变化研究[D]. 长沙: 中南林业科技大学, 2020. RAO Dandan. Study on Tissue Culture and Endogenous Hormone Content Changes of a New Lagerstroemia indica Variety ‘Ziyu’[D]. Changsha: Central South University of Forestry & Technology, 2020. [17] 李红, 李波, 杨曌. 外源ABA对苏打盐碱胁迫的紫花苜蓿内源激素含量的影响[J]. 黑龙江畜牧兽医, 2020(6): 103−106, 111. LI Hong, LI Bo, YANG Zhao. Effect of exogenous ABA on endogenous hormone content of alfalfa under salt and alkali stress [J]. Heilongjiang Animal Science and Veterinary Medicine, 2020(6): 103−106, 111. [18] NORIKOSHI R, SHIBATA T, ICHIMURA K. Cell division and expansion in petals during flower development and opening in Eustoma grandiflorum [J]. The Horticulture Journal, 2016, 85(2): 154−160. [19] 王卅, 张旸, 李玉花. 花瓣锥形表皮细胞形成及对授粉昆虫吸引作用机制研究进展[J]. 园艺学报, 2012, 39(9): 1781−1792. WANG Sa, ZHANG Yang, LI Yuhua. Research progress on the mechanism of petal conical epidermal cells formation and attraction to pollinating insects [J]. Acta Horticulturae Sinica, 2012, 39(9): 1781−1792. [20] SCHARWIES J D, DINNENY J R. Water transport, perception, and response in plants [J]. Journal of Plant Research, 2019, 132(3): 311−324. [21] 陈婧婧. 梅花开花进程及采后生理特性研究[D]. 杭州: 浙江农林大学, 2012. CHEN Jingjing. Study on Flowering Process and Postharvest Physiological Characteristics of Plum Blossom[D]. Hangzhou: Zhejiang A&F University, 2012. [22] 苏和. 2种观赏树木花开放与衰老过程中的生理变化研究[J]. 南方农业, 2021, 15(24): 19−20, 25. SU He. Study on physiological changes of two ornamental trees during flower opening and senescence [J]. South China Agriculture, 2021, 15(24): 19−20, 25. [23] 王支槐. 腊梅开花过程中的生理变化[J]. 西南师范大学学报(自然科学版), 1994, 19(6): 646−650. WANG Zhihuai. Physiological changes of Chimonanthus praecox during flowering [J]. Journal of Southwest China Normal University (Natural Science Edition), 1994, 19(6): 646−650. [24] LASTDRAGER J, HANSON J, SMEEKENS S. Sugar signals and the control of plant growth and development [J]. Journal of Experimental Botany, 2014, 65(3): 799−807. [25] MORITA S, SUGIYAMA S, TATEISHI A, et al. Identification and characterization of plasma membrane intrinsic protein (PIP) aquaporin genes in petals of opening carnation flowers [J]. The Horticulture Journal, 2017, 86(1): 78−86. [26] CHO L H, PASRIGA R, YOON J, et al. Roles of sugars in controlling flowering time [J]. Journal of Plant Biology, 2018, 61(3): 121−130. [27] 高天瑜, 郑斌, 许文天, 等. 杧果成花过程碳水化合物和可溶性蛋白含量的变化[J]. 中国南方果树, 2022, 51(3): 63−69. GAO Tianyu, ZHENG Bin, XU Wentian, et al. Changes on contents of carbohydrate and soluble protein during the flower formation process of mango [J]. South China Fruits, 2022, 51(3): 63−69. [28] 罗惠格, 朱维, 黄泳碧, 等. 阳光玫瑰葡萄生长期花芽分化形态进程及相关生理分子水平变化研究[J]. 果树学报, 2023, 40(1): 74−87. LUO Huige, ZHU Wei, HUANG Yongbi, et al. A study on the morphological process and physiological and molecular changes of flower bud differentiation in Shine Muscat grape during fruit growing season [J]. Journal of Fruit Science, 2023, 40(1): 74−87. [29] GU Jiahui, ZENG Zhen, WANG Yiru, et al. Transcriptome analysis of carbohydrate metabolism genes and molecular regulation of sucrose transport gene LoSUT on the flowering process of developing oriental hybrid lily ‘Sorbonne’ bulb[J/OL]. International Journal of Molecular Sciences, 2020, 21 (9): 3092 [2024-04-20]. DOI: 10.3390/ijms21093092. [30] SERCAN Ö, TONGUÇ M, DAMLA Ö, et al. Flower color and carbohydrate metabolism changes during the floral development of Rosa damascena [J]. South African Journal of Botany, 2023, 156: 234−243. [31] KITAMURA Y, YAMANE H, GAO-TAKAI M, et al. Changes in plant hormone contents in Japanese apricot flower buds during prolonged chilling exposure [J]. Acta Horticulturae, 2018(1208): 251−256. [32] 江海都, 孙菲菲, 秦惠珍, 等. 四季花金花茶花芽分化进程及叶片内源激素变化[J]. 广西植物, 2024, 44(1): 56−67. JIANG Haidu, SUN Feifei, QIN Huizhen, et al. Flower bud differentiation and leaf endogenous hormone changes of Camellia perpetua [J]. Guihaia, 2024, 44(1): 56−67. [33] 杨雪. 金花茶内源激素变化规律及顶生金花茶遗传多样性的SSR分析[D]. 南宁: 广西大学, 2016. YANG Xue. Changes of Endogenous Hormones in Camellia chrysantha and SSR Analysis of Genetic Diversity of Terminal Camellia chrysantha[D]. Nanning: Guangxi University, 2016. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240347
 点击查看大图
点击查看大图

计量
- 文章访问数: 2380
- HTML全文浏览量: 782
- PDF下载量: 40
- 被引次数: 0
