




 下载:
下载:

-
花芽分化是有花植物开花过程中最为重要的阶段,直接影响植物开花的质量和数量[1]。花芽分化是一个高度复杂的生理生化和形态发生过程,受自身调控和外界环境的共同影响[2-3]。目前,在许多物种中已经证实开花促进因子AP1 (APETALA1)、成花素FT (FLOWERING LOCUS T)以及LFY (LEAFY)等[4-5]参与调控花芽分化。SEP (SEPALLATA)是MADS-box转录因子家族中的一员,在花芽萌发时发挥着重要作用,能够调控B和C类功能基因的表达[6]。SEP蛋白可参与花器官的形成与发育,控制萼片、花瓣以及雄蕊的形成[7-8]。AP1可直接或通过LFY间接促进SEP的表达来控制早期的花发育过程[9]。另外,拟南芥Arabidopsis thaliana中的SEP3直接或间接地作用于SVP (SHORT VEGETATIVE PHASE)、SOC1(SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1)和FT等开花因子的下游基因[10],SEP3过表达能够控制其提前开花[11],且LiMADS3 (百合Lilium brownii var. viridulum的SEP3同源基因)转基因拟南芥植株也表现出早期开花表型[12]。PpCMB1隶属SEP类基因的SEP1亚组,将其转入拟南芥与正常生长的相比,PpCMB1过表达具有明显促进抽薹早花的表型[13]。但目前关于SEP类基因CMB1调控花分生组织、花器官和雄蕊等形成的研究较少。
紫薇Lagerstroemia indica是少数夏季开花的木本植物,花型奇特,花期可达100 d,极具观赏价值[14]。紫薇是顶生圆锥花序,其生长锥发育的过程中,首先由花序顶部的花原基先发育,继而两侧不断形成新的小花原基,最终形成完整的圆锥花序。关于紫薇花芽分化进程的研究较少,因此本研究拟通过对紫薇LiCMB1基因克隆,分析LiCMB1在花芽分化过程中不同时期以及不同组织中的表达模式,初步探究LiCMB1对紫薇花芽分化的调控作用,为紫薇的花期调控以及新品种选育等提供理论基础。
-
供试材料采自浙江农林大学紫薇种质资源苗圃(30°15′02″N, 119°43′37″E),选择12株生长条件一致,生长健壮,无病虫害的多年生紫薇植株为材料。观察自2021年3月9日开始,5月7日2次抽梢的新芽开始萌发后每5 d采1次,5月25日花芽开始分化后3 d取样1次,直至分化结束。
采样时在当年生枝条上不同方向二次抽梢后的嫩枝中,随机选取靠近顶芽相同位置的花芽20枚,各采集3份,分别用于石蜡切片和RNA提取。此外,在紫薇营养生长阶段(5月12日),取紫薇的根、嫩茎和嫩叶,在紫薇开花后(7月2日),取萼片、花瓣、雌蕊、长雄蕊和短雄蕊用于组织和器官特异性分析。
-
将鲜样置于体视显微镜下观察并用游标卡尺记录外部的形态大小变化,石蜡切片的制作方法参考SUN等[15]方法并改进。改进方法为:将FAA中固定24 h后的花芽,采用体积分数为50%和70%的无水乙醇分别脱水30 min,转入体积分数为70%的无水乙醇过夜;次日按照从低至高的无水乙醇,二甲苯浓度梯度进行脱水透明处理;经过浸蜡,包埋,切片(LEICA RM2235),厚度为9 μm,番红-固绿双重染色等步骤后,中性树胶封片后,置于显微镜(Axio Imager 2,日本)下观察并拍照。
-
分别提取不同分化时期的花芽和不同组织的样品各0.5 g,按照Plant Total Isolation Kit试剂盒说明书(Vazyme,中国南京)提取 RNA。cDNA反转录参照HiScriptⅢReverse Transcriptase(Vazyme)说明书进行,反转录后的cDNA保存于−20 ℃冰箱备用。
-
从转录组数据中筛选到LiCMB1基因蛋白质编码区(CDS)片段,运用 Primer Premier 5.0软件设计特异性引物(F:ATGGGGAGAGGGAAGGTCGA;R:TTAAAGCGTCCAGTCTTGAAGG),由浙江有康生物科技有限公司合成,以反转录的 cDNA 为模板进行 PCR扩增。PCR 反应体系如下:95 ℃预变性5 min;95 ℃变性30 s,55 ℃退火 30 s,72 ℃延伸 1 min,35个循环;72 ℃延伸10 min,10 ℃延伸5 min。PCR扩增产物置于质量浓度为1%的琼脂糖凝胶电泳分离,切胶回收,然后连接至pMDTM18-T载体后转化大肠埃希菌Escherichia coli DH5α,涂板后进行蓝白斑筛选,挑菌经PCR鉴定获得阳性克隆,将菌液送至浙江有康生物科技有限公司进行测序。
-
采用ORF Finder (
https://www.ncbi.nlm.nih.gov/orffinder/ )将核苷酸序列翻译为氨基酸序列,采用美国国家生物技术信息中心(NCBI)中CDD (Conserved Domain Datebase)在线软件分析保守区域。用Jalview软件(http://www.jalview.org/ )对LiCMB1基因的氨基酸序列进行多重序列比对,使用MEGA 6.0软件邻接法(NJ)构建系统发育树。采用ExPasy提供的Prot-Param (https://web.expasy.org/protparam/ )预测编码蛋白的基本理化性质,采用NetPhos 对磷酸化位点进行分析。分别采用SOPMA (https://npsa-prabi.ibcp.fr/ )和Cell-PLoc 2.0 (http://www.csbio.sjtu. edu.cn/bioinf/Cell-PLoc-2/ )进行蛋白质二级结构和亚细胞定位预测情况。 -
设计实时荧光定量PCR (RT-qPCR)上下游引物(F-q:TCGCCGCCATCATCTTCTC;R-q:CTCTAGCTCCTTTATGCTCATCTCC),并以LiEF-1α作为内参基因[16]。实时荧光定量反应体系如下:SYBR PremixEx Taq 5 μL,cDNA 2 μL,上下游引物各0.4 μL,ddH2O 2.2 μL。RT-qPCR反应程序为:95 ℃预变性30 s,95 ℃变性5 s,60 ℃复性30 s,40个循环,每个样品3次生物学重复。根据
$2^{-\Delta \Delta C_t} $ 法[17]分析LiCMB1的相对表达量,采用Excel 2010和SPSS 22进行数据分析,采用SigmaPlot 12.5进行绘图。 -
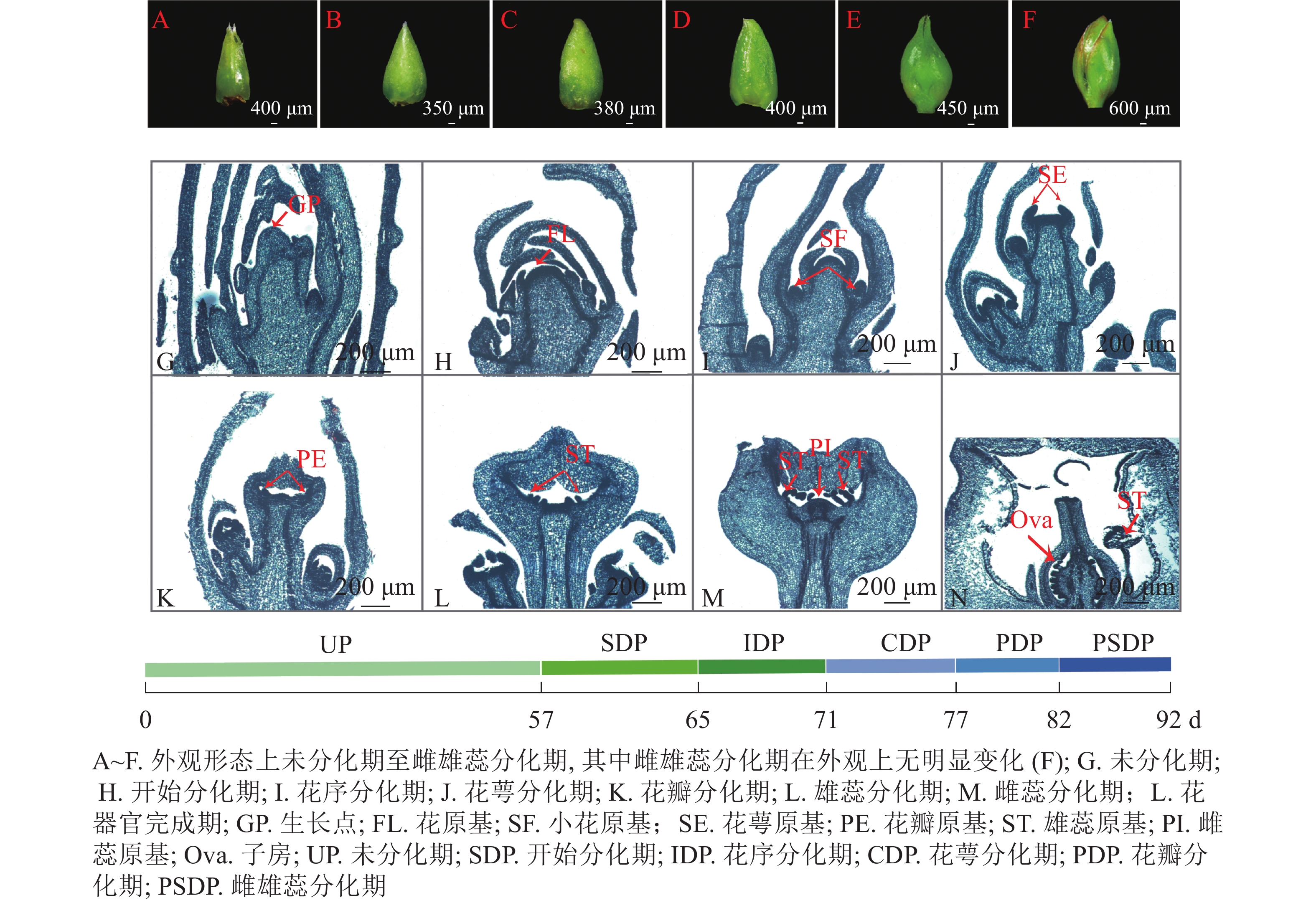
根据改进后的石蜡切片结果以及体视显微镜的观察记录,紫薇在新生营养芽展叶第57 天 (2021年5月25日)后开始进入花芽分化,第82 天(2021年6月18日)后雌、雄蕊出现,第92天(2021年6月28日)时花芽分化结束,形成完整的花器官。整个花芽分化过程可以划分为未分化期、开始分化期、花序分化期、花萼分化期、花瓣分化期、雄蕊分化期以及雌蕊分化期等7个时期(图1)。花芽分化时间集中在5月底至6月中下旬。

图 1 紫薇花芽分化的进程图
Figure 1. Process of flower bud differentiation of L. indica
-
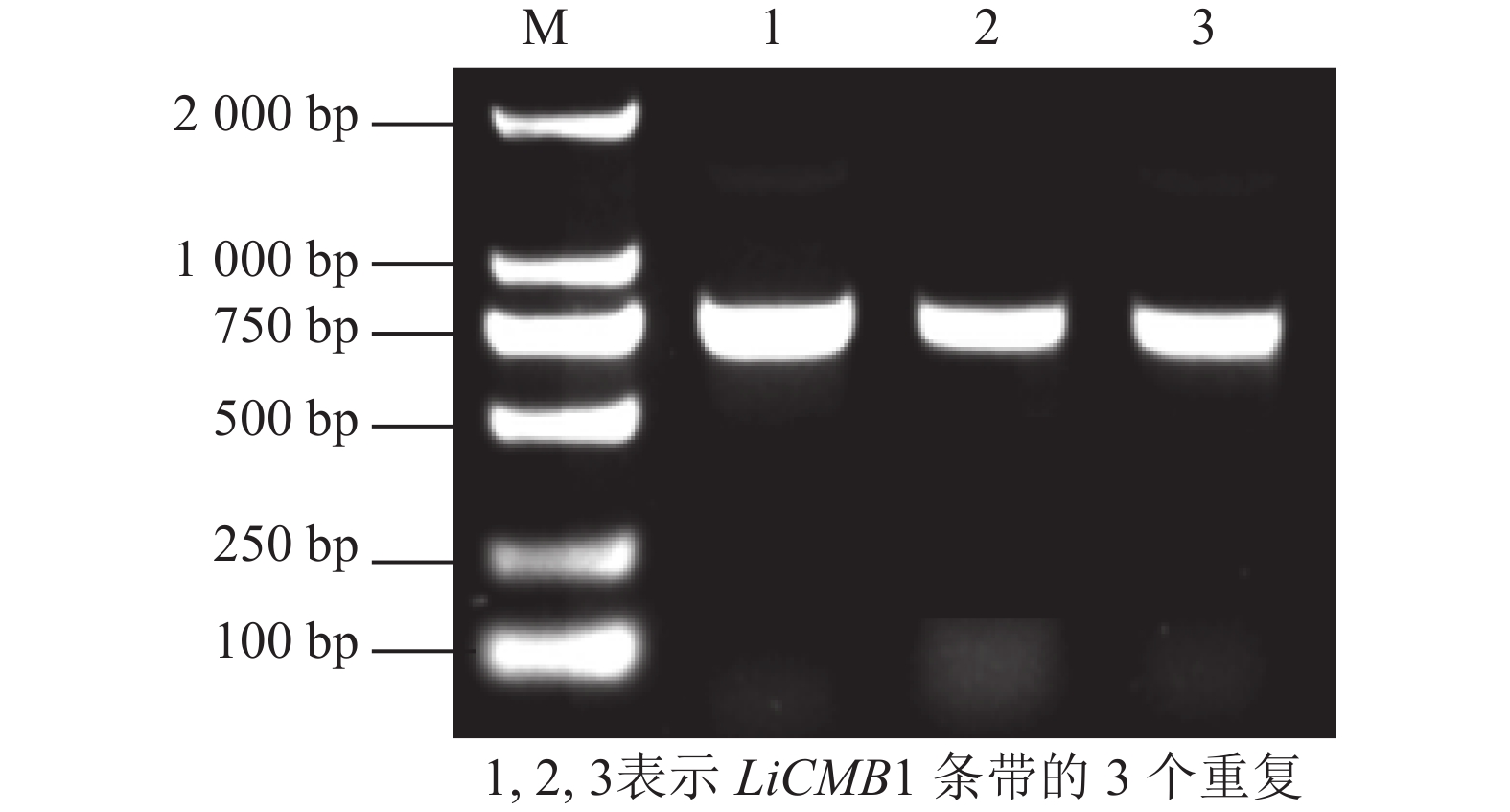
以紫薇的花芽cDNA为模板, PCR扩增获得完整的ORF序列,长度为756 bp(图2),共编码251个氨基酸(图3)。通过NCBI在线BLASTX (
https://blast.ncbi.nlm.nih.gov/Blast.cgi )分析,发现该序列与石榴Punica granatum (XP_031400101.1)、可可树Theobroma cacao (XP_017970029.1)的CMB1基因同源性最高,将其命名为LiCMB1。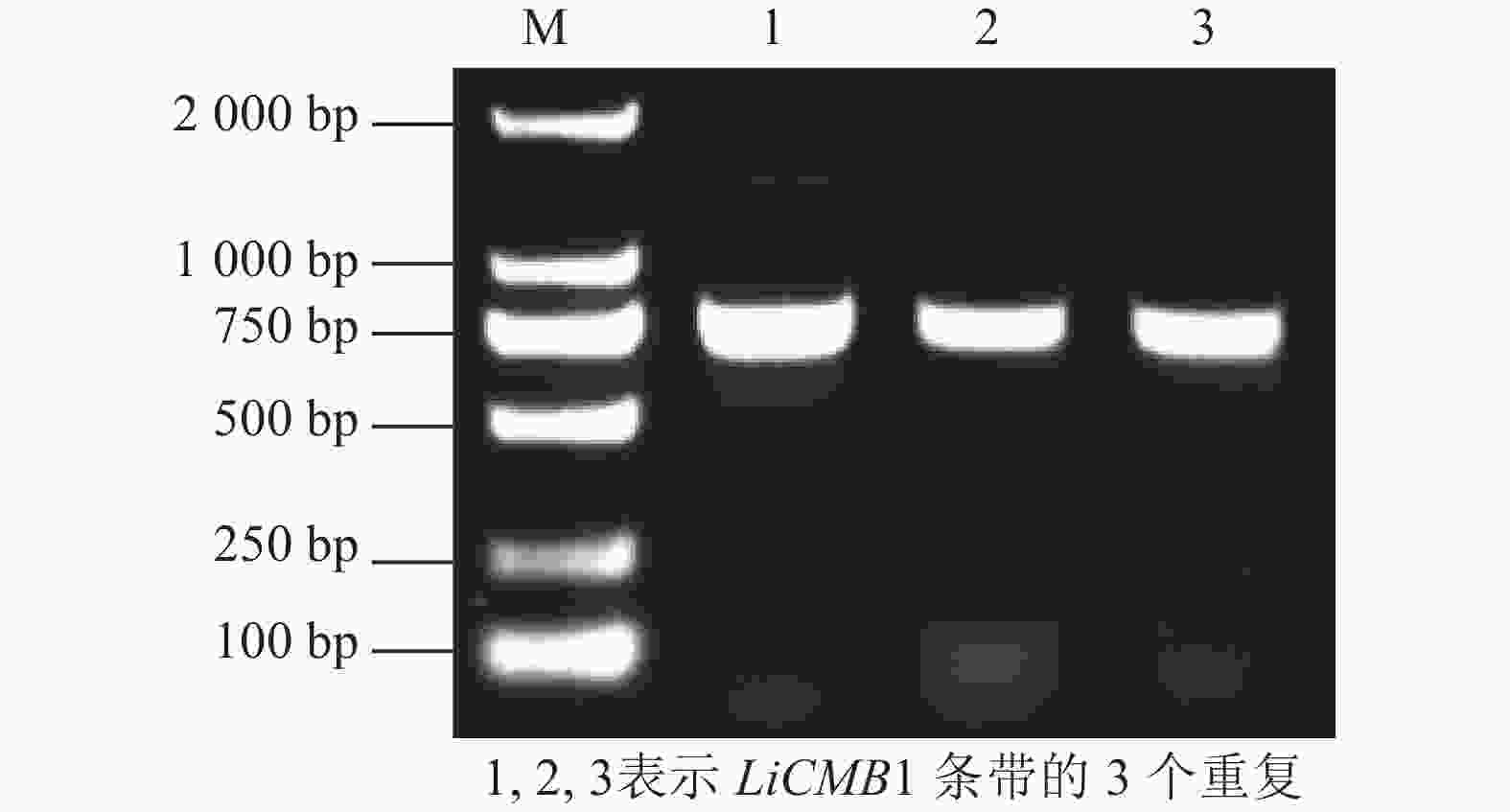
图 2 紫薇LiCMB1的电泳图
Figure 2. Cloning of LiCMB1

图 3 紫薇LiCMB1的氨基酸推导
Figure 3. Amino acid sequence derivation of LiCMB1
-
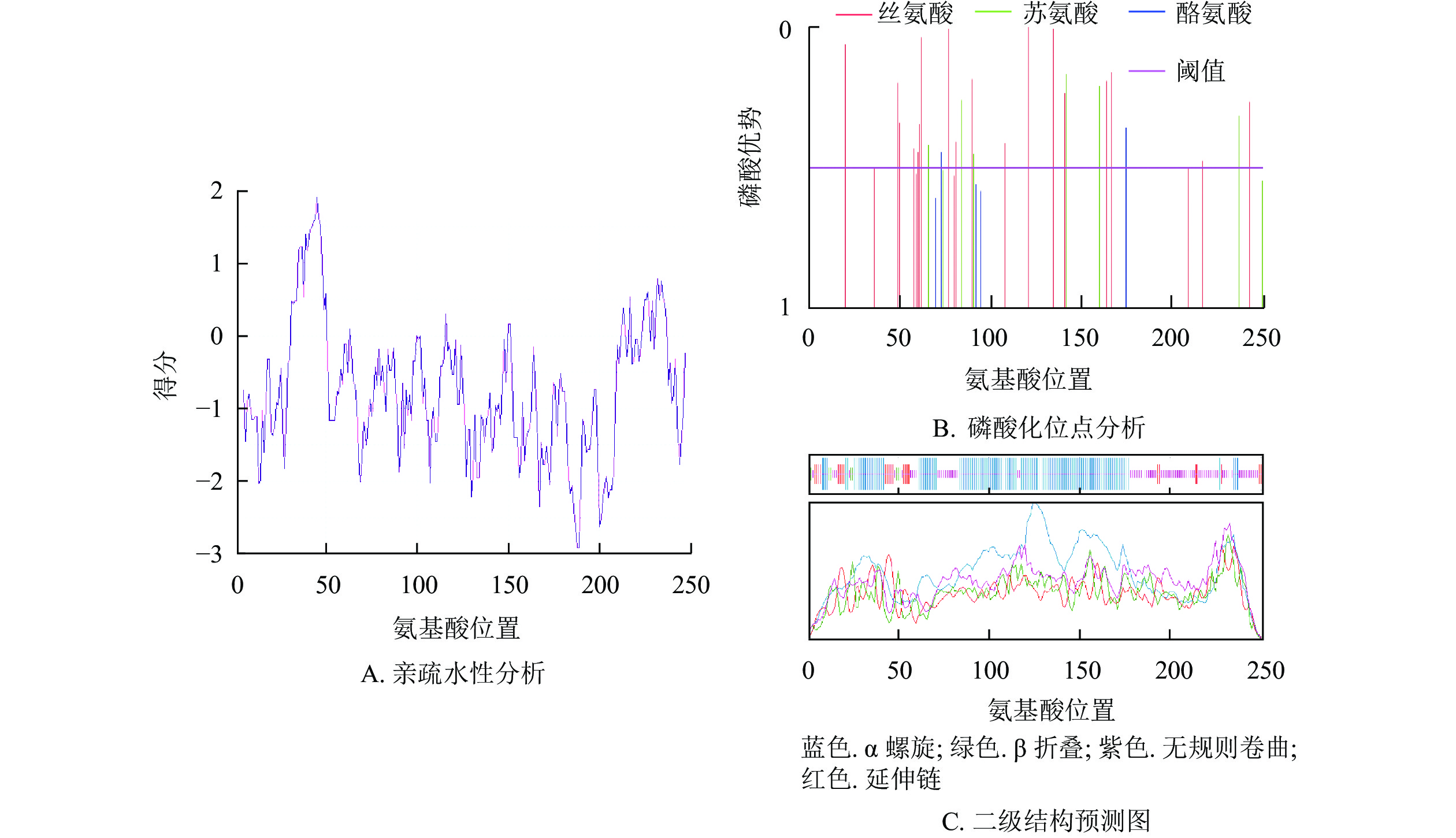
预测紫薇LiCMB1蛋白分子式为C1250H2027N383O387S10,相对分子量为28 933.78,等电点为8.93,亲水性平均系数为−0.757,为亲水性蛋白(图4A),脂肪系数为79.92。最富氨基酸占比为谷氨酸(Glu)和赖氨酸(Leu),氨基酸组成成分分析表明:LiCMB1蛋白肽链正电荷残基[精氨酸(Arg)+赖氨酸(Lys)]总数为39,负电荷残基[天冬氨酸(Asp)+谷氨酸(Glu)]为35,推测其为碱性蛋白。NetPhos分析显示:LiCMB1存在27个磷酸化修饰位点,其中丝氨酸磷酸化位点18个,苏氨酸磷酸化位点6个, 酪氨酸磷酸化位点3个(图4B)。亚细胞定位预测结果表明LiCMB1位于细胞核中。采用NCBI-CDD保守结构域分析发现:LiCMB1具有典型的MADS_MEF2_like和K-box结构域。SOPMA在线网站预测蛋白的二级结构显示:该蛋白质含有51.79%的α-螺旋,3.59%的β-折叠,9.56%的延伸和33.06%的无规则卷曲(图4C)。
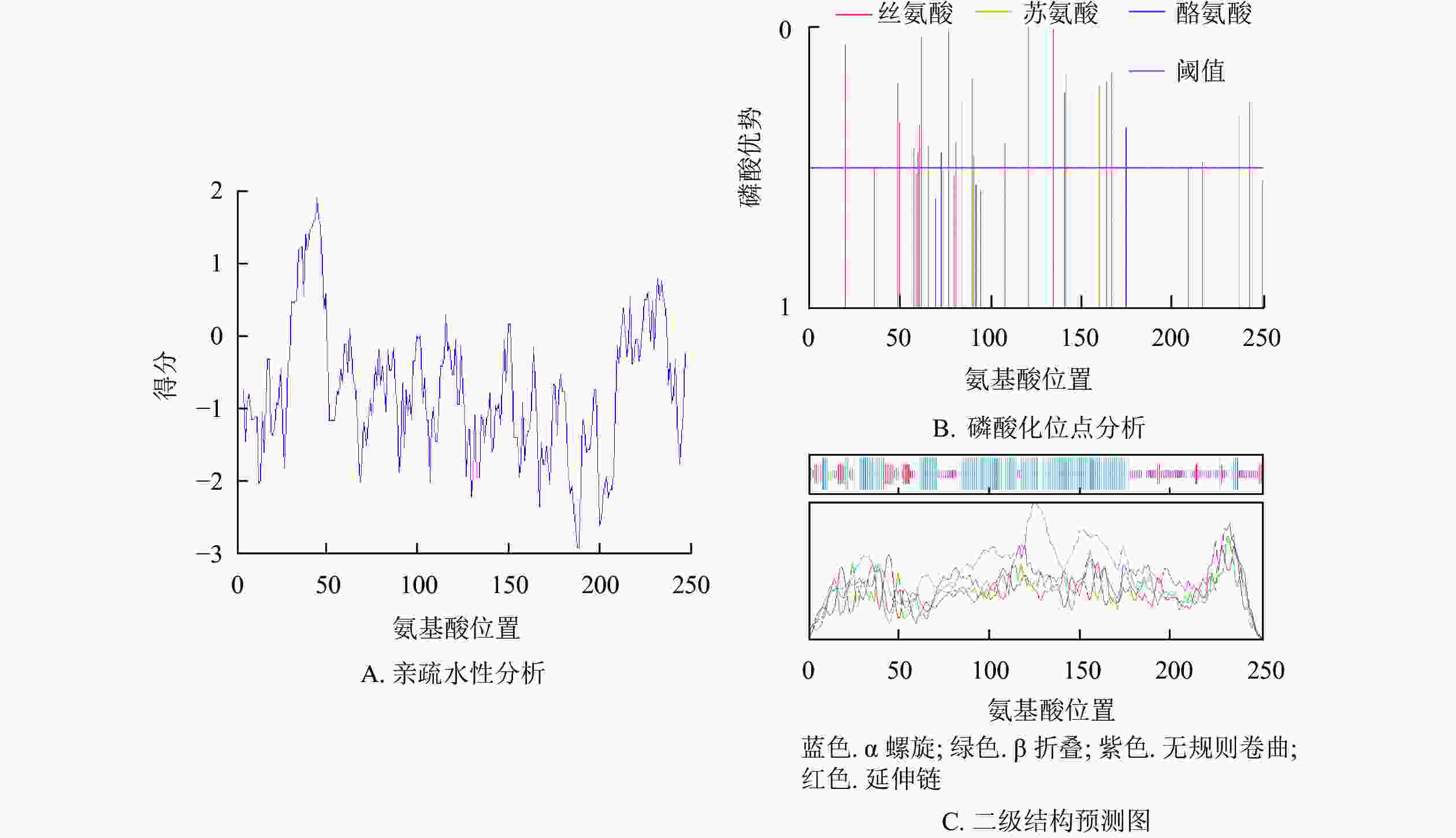
图 4 紫薇LiCMB1蛋白理化性质和二级结构预测图
Figure 4. Physicochemical properties and secondary prediction diagram of protein of LiCMB1
-
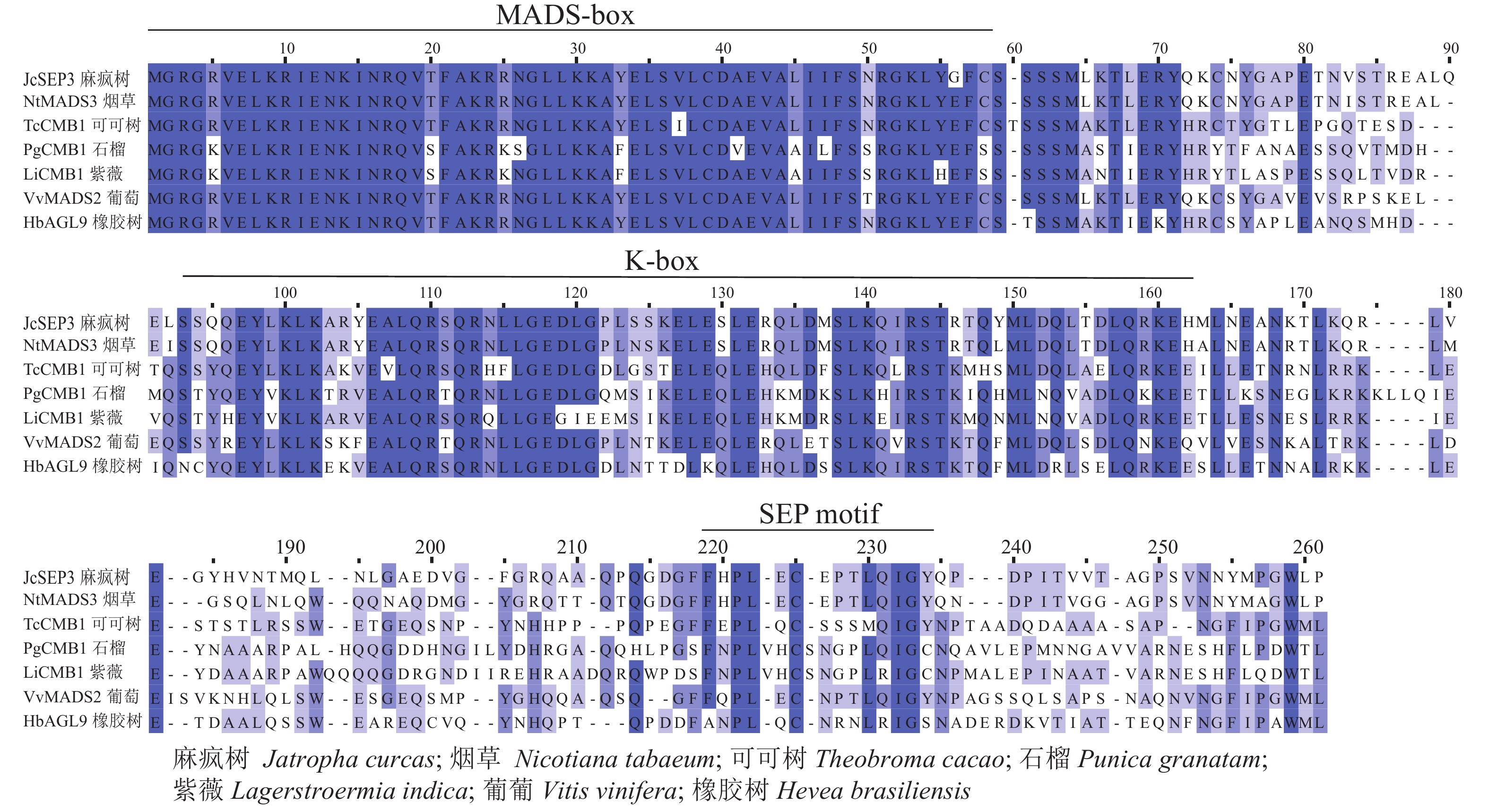
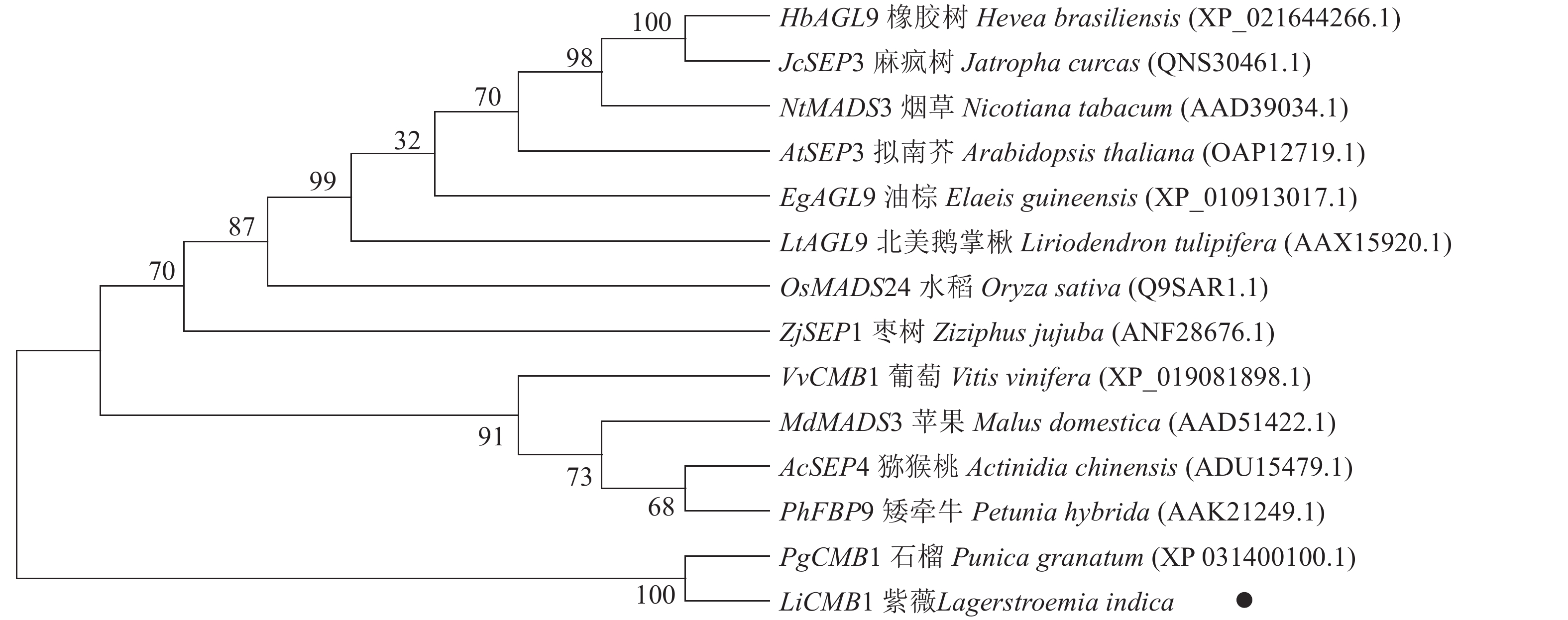
采用Jalview软件将其与其他物种的SEP类蛋白进行比对发现:其氨基酸序列基本相似,除了含有MADS-box典型的结构域外,靠近C端处还含有一个典型的SEP motif保守基序(图5)。采用MEGA 6.0将紫薇LiCMB1与其他物种的SEP类蛋白构建系统发育树(图6):LiCMB1与葡萄Vitis vinifera的VvCMB1以及其他物种的SEP类蛋白亲缘关系相对较远,而与石榴的PgCMB1具有较近的亲缘关系,蛋白质相似度高达78.97%。
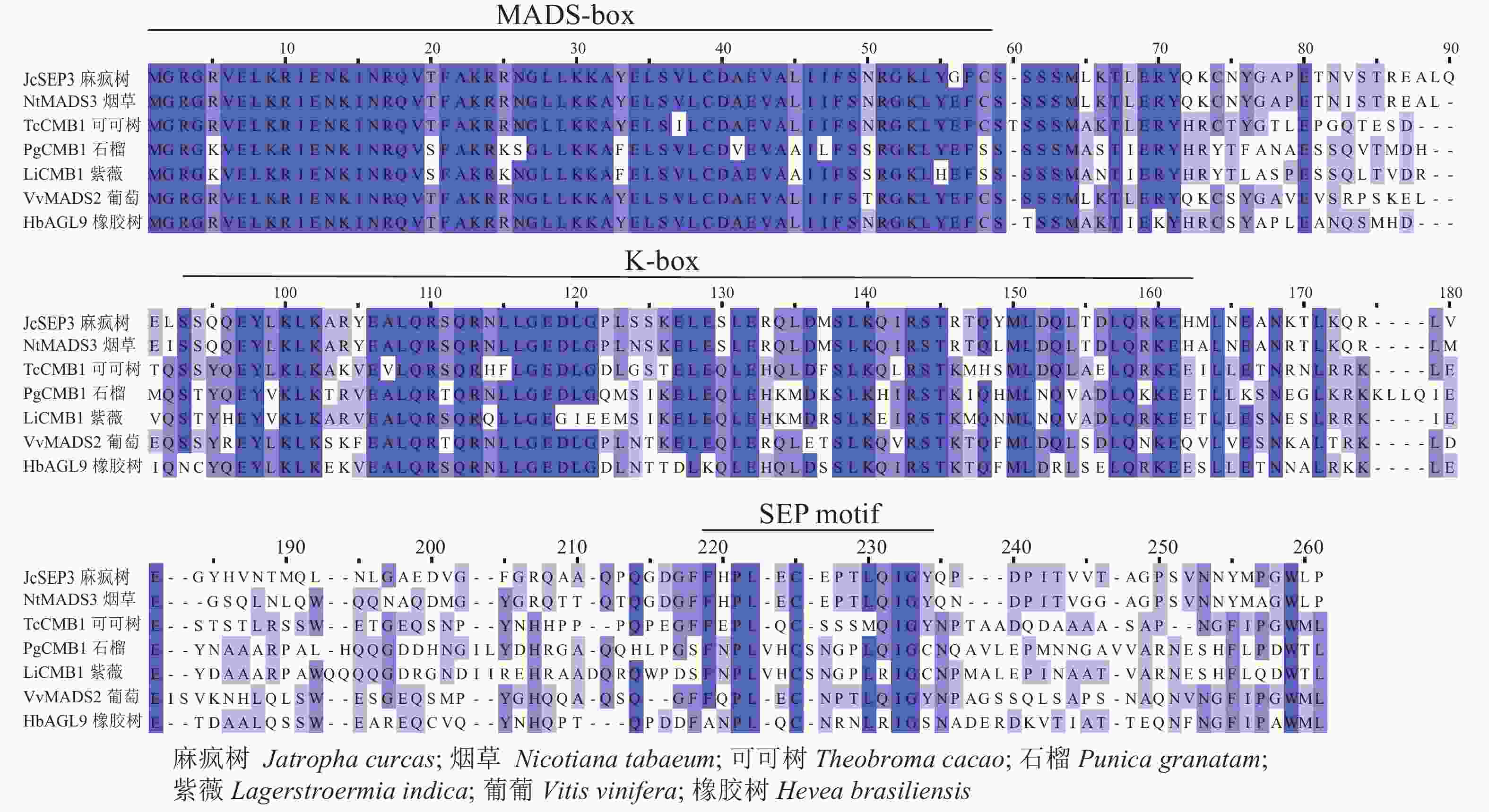
图 5 紫薇LiCMB1与其他物种的SEP类蛋白序列比对
Figure 5. Comparative analysis of LiCMB1 protein sequence
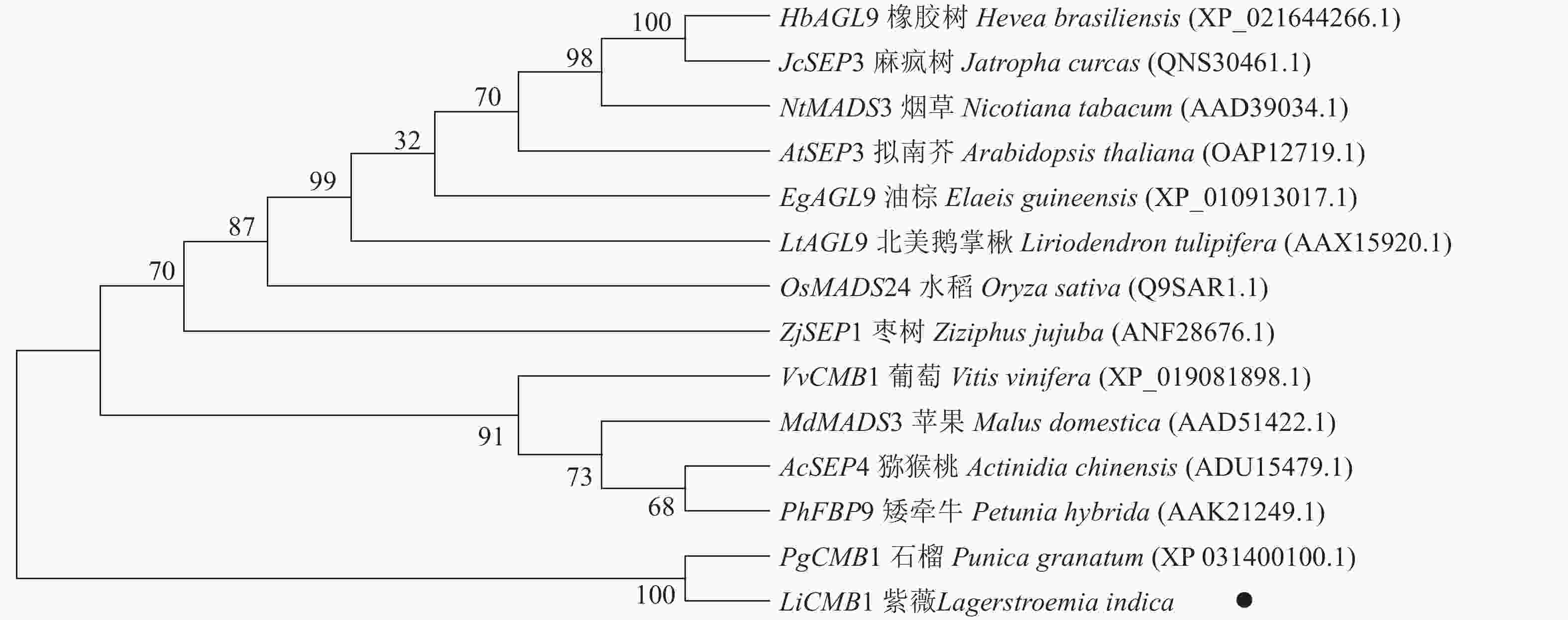
图 6 紫薇LiCMB1与其他物种SEP类蛋白系统进化树
Figure 6. Phylogenetic tree of LiCMB1 and SEP like proteins of other species
-
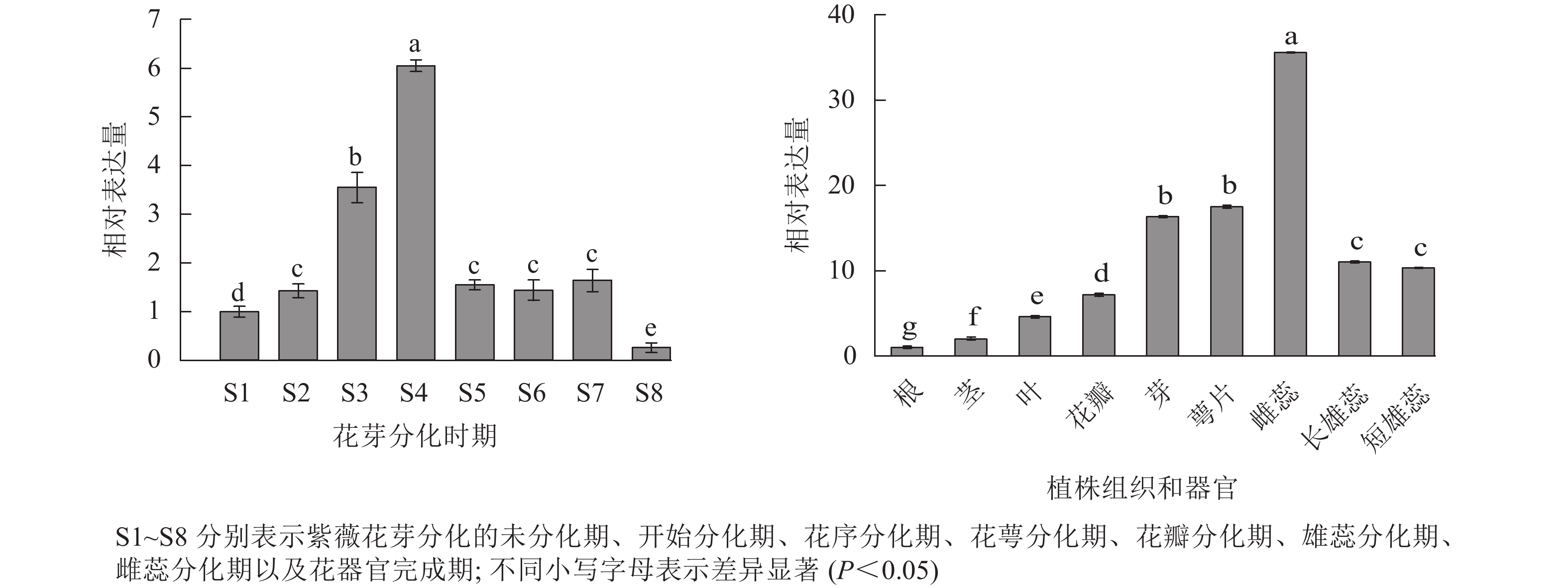
采用RT-qPCR技术,对紫薇花芽分化的7个时期、花器官完成期以及9个不同组织和器官中LiCMB1基因的表达情况进行分析(图7)。结果表明:LiCMB1在紫薇花芽分化的过程中呈现先上升后下降,然后趋于稳定的趋势,在花萼分化期时表达量最高,约为未分化时期表达量的6倍;此外,LiCMB1在不同的组织和器官中都有表达,在根中的表达量最低,其次是茎,在雌蕊中的表达量最高,约根中表达的35倍,萼片和花芽的表达量次之。而LiCMB1在长雄蕊和短雄蕊中的表达量没有显著差异,在花瓣中的表达相对于芽、萼片、雌蕊、长雄蕊和短雄蕊而言较弱,但对于根、茎和叶表达量较高。

图 7 紫薇LiCMB1基因的相对表达量
Figure 7. Relative expression of LiCMB1 gene
-
植物从营养生长到生殖生长的转变是开花植物发育的关键过程[18]。在这一发育过程中,紫薇花芽分化显著的变化是花序和花的形成。许多MADS-box家族基因,特别是SEP MADS-box转录因子在调控花芽分化过程中发挥着重要作用[11, 19]。本研究发现:紫薇处于未分化期时花芽较小,当开始分化直至雌、雄蕊出现时芽体不断增长膨大。然后根据各部分原基的出现进行时期的划分,将紫薇的花芽分化过程划分为未分化期、开始分化期、花序分化期、花萼分化期、花瓣分化期、雄蕊分化期以及雌蕊分化期等7个时期,并从紫薇中克隆到756 bp的LiCMB1基因,共编码251个氨基酸,LiCMB1的氨基酸序列除了含有典型的MADS_MEF2_like和K-box结构域,在靠近C端的位置还含有相对保守的 SEP基序。该基序与SEP3蛋白复合物形成有关[20],推测SEP基序可能与LiCMB1蛋白的功能有重要关系。系统进化树分析显示:LiCMB1与石榴的PgCMB1蛋白亲缘关系最近,蛋白质相似度高达78.97%,这可能因为紫薇和石榴都属于桃金娘目Myrtales。而与葡萄、苹果Malus domestica、拟南芥、烟草Nicotiana tabacum等植物的SEP类蛋白[21]关系相对较远。
SlCMB1
在番茄Solanum lycopersicum植株组织中的表达谱分析表明:SlCMB1可能参与调控花序和萼片的发育[22]。从桃Prunus persica中分离出的PpCMB1,可能通过抑制PpDAM5蛋白的功能促进花发育,且PpCMB1主要在萼片和雌蕊中高表达[13]。本研究RT-qPCR结果表明:LiCMB1整体呈先上升后下降的表达趋势,这说明其在紫薇花芽分化过程的前期可能起到促进分化的作用。特别是在花序分化期至花萼分化期时表达量较高,这表明LiCMB1参与了紫薇花芽分化的过程并在花萼分化期起到重要作用。在形成完整的花器官后,LiCMB1表达量相对较少,可能是其在该时期不发挥作用。此外,通过组织特异性分析发现:LiCMB1在各组织和器官中均有表达,在花瓣、芽、萼片和雌雄蕊等组织中显著提高,且在雌蕊和萼片中的表达量要高于长雄蕊、短雄蕊和花瓣中的表达量。这说明LiCMB1可能在调控花器官发育过程中发挥了作用,并且可能参与了雌蕊和萼片的发育过程。 在经典的花发育ABCDE模型中,SEP亚组属于E类基因, 通常起到调控花瓣、萼片、雄蕊等的发育[23-24]。SEP类基因VvMADS6能够促进葡萄花芽萌发,表达量相对也较高[25]。重瓣百合Lilium brownii‘Belonica’中LiSEP3的表达模式与双子叶植物的有所不同,其主要在花中表达量较高,并且在最内侧的花瓣中表达量最高[26]。番茄的SEP 类基因SlCMB1在花序中显著表达,其转录水平在花萼中也较高[22],可能正向调控番茄的花发育。PsMADS4基因在牡丹Paeonia suffruticosa花发育过程中起着重要作用,且与牡丹花的衰老具有密切关系[27]。PheMADS15基因在毛竹Phyllostachy edulis花发育的初期表达量最高,主要在花芽中表达,可能参与毛竹成花转变过程[28]。
本研究通过对LiCMB1蛋白的基本性质以及其在花芽分化各个时期、不同组织和器官的表达特征做了初步探索,发现LiCMB1在紫薇花芽分化过程中的花萼分化期起关键作用,并且可能参与调控花器官的发育过程。但LiCMB1是否会与其他基因互作共同促进紫薇的花芽分化以及花发育,具体的调控机制还有待进一步的验证。
Cloning and expression characteristics of LiCMB1 gene in Lagerstroemia indica
-
摘要:
目的 克隆紫薇Lagerstroemia indica LiCMB1基因并分析其在紫薇花芽分化的不同时期及不同组织和器官中的表达,探讨LiCMB1基因的表达特性。 方法 利用简单克隆技术从紫薇中克隆得到LiCMB1的基因序列,通过ExPasy等在线工具对其进行蛋白质理化性质分析,使用MEGA 6.0构建系统进化树,结合紫薇花芽分化的表型观察和石蜡切片,采用实时荧光定量PCR分析花芽分化的不同时期及不同组织和器官中LiCMB1基因的表达。 结果 LiCMB1基因属于MADS-box家族SEP类基因,除了具有典型的MADS_MEF2_like和K-box结构域外,靠近C端处还含有一个SEP motif保守基序;LiCMB1在紫薇花芽分化过程中呈现先上升后下降的表达趋势,在各组织和器官中均有表达,表达量从高到低依次为雌蕊、萼片、芽、长雄蕊、短雄蕊、花瓣、叶、茎、根,说明LiCMB1可能对紫薇的花芽分化起到重要作用,且参与调控花器官发育。 结论 LiCMB1基因属于MADS-box家族的SEP基因,在紫薇花芽分化的前期发挥重要作用,尤其是在花萼分化期表达量最高,组织特异性分析表明该基因很可能参与了调控花器官发育。图7参28 Abstract:Objective The LiCMB1 gene of Lagerstroemia indica was cloned, and its expressions in different stages of flower bud differentiation and different tissues and organs were analyzed, so as to explore the expression characteristics of LiCMB1 gene. Method The gene sequence of LiCMB1 was cloned from L. indica by simple cloning technology. Physical and chemical properties of the protein were analyzed by online tools including ExPasy, and phylogenetic tree was constructed by MEGA 6.0 software. Combined with the phenotypic observation and paraffin section of L.indica flower bud differentiation, the expressions of LiCMB1 gene in different stages of flower bud differentiation and different tissues and organs were analyzed by real-time quantitative PCR (RT-qPCR). Result LiCMB1 gene belongs to SEP gene of MADS-box family, except for typical MADS_ MEF2_ like and K-box structure domain, there is also a SEP motif conserved motif near the C-end. The results of RT-qPCR showed that the expression trend of LiCMB1 increased first and then decreased in the process of flower bud differentiation of L. indica. It is expressed in different tissues and organs, and the expression levels of LiCMB1 from high to low were in the order of pistil, sepal, bud, long stamen, short stamen, petal, leaf, stem, root, indicating that LiCMB1 may play an important role in flower bud differentiation and participate in the regulation of flower organ development. Conclusion LiCMB1 gene belongs to the SEP gene of MADS-box family. It plays an important role in the early stage of flower bud differentiation of L. indica, especially in the calyx differentiation period. Tissue specificity analysis indicated that it was likely involved in the regulation of floral organ development. [Ch, 7 fig. 28 ref.] -
图 4 紫薇LiCMB1蛋白理化性质和二级结构预测图
Figure 4 Physicochemical properties and secondary prediction diagram of protein of LiCMB1
图 6 紫薇LiCMB1与其他物种SEP类蛋白系统进化树
Figure 6 Phylogenetic tree of LiCMB1 and SEP like proteins of other species
-
[1] 董晓晓, 别沛婷, 袁涛. 3个牡丹品种花芽分化过程形态及叶片碳水化合物质量分数变化[J]. 东北林业大学学报, 2020, 48(7): 34 − 39. DONG Xiaoxiao, BIE Peiting, YUAN Tao. Changes of morphology and carbohydrate content in leaves of three tree peony during flower bud differentiation [J]. Journal of Northeast Forestry University, 2020, 48(7): 34 − 39. [2] 郜爱玲, 李建安, 刘儒, 等. 高等植物花芽分化机理研究进展[J]. 经济林研究, 2010, 28(2): 131 − 136. GAO Ailing, LI Jian’an, LIU Ru, et al. Advance in research on flower bud differentiation mechanism in higher plants[J] Nonwood Forest Research, 2010, 28(2): 131 − 136. [3] 曲波, 张微, 陈旭辉, 等. 植物花芽分化研究进展[J]. 中国农学通报, 2010, 26(24): 109 − 114. QU Bo, ZHANG Wei, CHEN Xuhui, et al. Research progress of flower bud differentiation mechanism of plant [J]. Chinese Agricultural Science, 2010, 26(24): 109 − 114. [4] HUANG Tao, BÖHLENIUS H, ERIKSSON S, et al. The mRNA of the Arabidopsis gene FT moves from leaf to shoot apex and induces flowering [J]. Science, 2005, 309(5741): 1694 − 1696. [5] MANDEL M A, GUSTAFSON-BROWN C, SAVIDGE B, et al. Molecular characterization of the Arabidopsis floral homeotic gene APETALA1 [J]. Nature, 1992, 360(6401): 273 − 277. [6] LIU Chang, XI Wanyan, SHEN Lisha, et al. Regulation of floral patterning by flowering time genes [J]. Developmental Cell, 2009, 16(5): 711 − 722. [7] IMMINK R G H, TONACO I A N, de FOLTER S, et al. SEPALLATA3: the ‘glue’ for MADS box transcription factor complex formation[J/OL]. Genome Biology, 2009, 10(2): R24[2022-04-21]. doi: 10.1186/gb-2009-10-2-r24. [8] SEYMOUR G B, RYDER C D, CEVIK V, et al. A SEPALLATA gene is involved in the development and ripening of strawberry (Fragaria x ananassa Duch. ) fruit, a non-climacteric tissue [J]. Journal of Experimental Botany, 2011, 62(3): 1179 − 1188. [9] GREGIS V, SESSA A, COLOMBO L, et al. AGAMOUS-LIKE24 and SHORT VEGETATIVE PHASE determine floral meristem identity in Arabidopsis [J]. The Plant Journal, 2008, 56(6): 891 − 902. [10] TEPER-BAMNOLKER P, SAMACH A. The flowering integrator FT regulates SEPALLATA3 and FRUITFULL accumulation in Arabidopsis leaves [J]. The Plant Cell, 2005, 17(10): 2661 − 2675. [11] PELAZ S, GUSTAFSON-BROWN C, KOHALMI S E, et al. APETALA1 and SEPALLATA3 interact to promote flower development [J]. The Plant Journal, 2001, 26(4): 385 − 394. [12] TZENG T Y, HSIAO C C, CHI P J, et al. Two lily SEPALLATA-like genes cause different effects on floral formation and floral transition in Arabidopsis [J]. Plant Physiology, 2003, 133(3): 1091 − 1101. [13] 张新昊, 沈红艳, 文滨滨, 等. 桃MADS-box家族PpCMB1基因调控花发育分子机制[J]. 植物生理学报, 2021, 57(6): 1211 − 1217. ZHANG Xinhao, SHEN Hongyan, WEN Binbin, et al. Molecular mechanism of PpCMB1 gene in peach MADS-box family regulating flower development [J]. Plant Physiology Journal, 2021, 57(6): 1211 − 1217. [14] ZHANG Qixiang. Studies on cultivars of crape myrtle (Lagerstroemia indica) and their uses in urban greening [J]. Journal of Beijing Forestry University, 1991, 13(4): 57 − 66. [15] SUN Chunqing, CHEN Fadi, TENG Nianjun, et al. Factors affecting seed set in the crosses between Dendranthema grandiflorum (Ramat. ) Kitamura and its wild species [J]. Euphytica, 2010, 171(2): 181 − 192. [16] ZHENG Tangchun, CHEN Zhilin, JU Yiqian, et al. Reference gene selection for qRT-PCR analysis of flower development in Lagerstroemia indica and L. speciosa [J/OL]. PLoS One, 2018, 13(3): e0195004[2022-04-21]. doi: 10.1371/journal.pone.0195004. [17] LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔC t method [J]. Methods, 2001, 25(4): 402 − 408. [18] 陈晨, 喻方圆. 林木花芽分化研究进展[J]. 林业科学, 2020, 56(9): 119 − 129. CHEN Chen, YU Fangyuan. Research progress on flower bud differentiation of trees [J]. Scientia Silvae Sinicae, 2020, 56(9): 119 − 129. [19] PELAZ S, DITTA G S, BAUMANN E, et al. B and C floral organ identity functions require SEPALLATTA MADS-box genes [J]. Nature, 2000, 405(6783): 200 − 203. [20] HONMA T, GOTO K. Complexes of MADS-box proteins are sufficient to convert leaves into floral organs[J]. Nature, 2001, 409(6819): 525 − 529. [21] ZAHN L M, KONG H, LEEBENS-MACK J H, et al. The evolution of the SEPALLATA subfamily of MADS-box genes: a preangiosperm origin with multiple duplications throughout angiosperm history [J]. Genetics, 2005, 169(4): 2209 − 2223. [22] ZHANG Jianling, HU Zongli, WANG Yunshu, et al. Suppression of a tomato SEPALLATA MADS-box gene, SlCMB1, generates altered inflorescence architecture and enlarged sepals [J]. Plant Science, 2018, 272: 75 − 87. [23] KAUFMANN K, MELZER R, THEISSEN G. MIKC-type MADS-domain proteins: structural modularity, protein interactions and network evolution in land plants [J]. Gene, 2005, 347(2): 183 − 198. [24] WEIGEL D, MEYEROEITZ E M. The ABCs of floral homeotic genes [J]. Cell, 1994, 78(2): 203 − 209. [25] 黄晓婧, 张珺, 夏惠, 等. 葡萄MADS-box转录因子家族全基因组鉴定及表达分析[J]. 园艺学报, 2019, 46(10): 1882 − 1896. HUANG Xiaojing, ZHANG Jun, XIA Hui, et al. Genome-wide identification and expression analysis of the MADS-box gene family in Vitis vinifera [J]. Acta Horticulturae Sinica, 2019, 46(10): 1882 − 1896. [26] 隋娟娟, 李晓昕, 杨秋燕, 等. 重瓣百合LiSEP3基因克隆与表达分析[J]. 南京林业大学学报(自然科学版), 2017, 41(1): 42 − 48. SUI Juanjuan, LI Xiaoxin, YANG Qiuyan, et al. Cloning and expression analysis of gene LiSEP3 in double lily [J]. Journal of Nanjing Forestry University (Natural Science Edition), 2017, 41(1): 42 − 48. [27] 姜纯, 周陶敏, 盖树鹏, 等. 牡丹PsMADS4基因的克隆、表达分析及载体构建[J]. 植物生理学报, 2021, 57(1): 94 − 100. JIANG Chun, ZHOU Taomin, GAI Shupeng, et al. Cloning, expression analysis and vector construction of tree peony PsMADS4 gene [J]. Plant Physiology Journal, 2021, 57(1): 94 − 100. [28] 程占超, 马艳军, 侯丹, 等. 毛竹PheMADS15基因的克隆及功能分析[J]. 浙江农林大学学报, 2017, 34(3): 421 − 426. CHENG Zhanchao, MA Yanjun, HOU Dan, et al. Isolation and functional analysis of the PheMADS15 gene in Phyllostachys edulis [J]. Journal of Zhejiang A&F University, 2017, 34(3): 421 − 426. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220333
 点击查看大图
点击查看大图
图(7)
计量
- 文章访问数: 1813
- HTML全文浏览量: 417
- PDF下载量: 128
- 被引次数: 0

