




 下载:
下载:
-
闽楠Phoebe bournei被列为国家二级保护濒危树种[1−2],是“金丝楠木”的原植物种之一,天然分布于福建、浙江、江西、湖南、云南和广西等省[3]。闽楠木材纹理细致、结构紧密、香气浓郁、防虫耐腐、坚硬不易开裂,是制作高档家具、建筑和雕塑的优质木材。闽楠树体高大挺拔、冠型优美,现已成为中国南方地区珍贵造林与园林绿化树种。然而,干旱、低温等非生物胁迫显著影响闽楠的生长发育,极大限制了林分生产力。
转录因子作为一类与启动子顺式作用元件特异结合、调控功能基因表达的DNA结合蛋白,对植物多种生理生化过程起重要调控作用[4−5]。其中,碱性亮氨酸拉链(bZIP)转录因子是目前已知普遍存在、成员较多、功能复杂且高度保守的基因家族之一[6−7]。bZIP转录因子是按照其高度保守的结构域来命名的,保守结构域包括富含亮氨酸残基的拉链区域和N-X7-R/K碱性区域,由60~80个碱基组成,C端N-X7-R/K碱性区域由20个氨基酸构成,富含精氨酸和赖氨酸,可与DNA特异性结合;N端亮氨酸拉链区域富含亮氨酸,相邻7个氨基酸构成一段连续片段,且第7位碱基是亮氨酸,但亮氨酸拉链区域不完全保守,个别位点的亮氨酸可被甲硫氨酸、缬氨酸及异亮氨酸等疏水性氨基酸所取代[6]。
bZIP转录因子功能多样性,广泛参与调控脱落酸(ABA)、干旱、高盐及高温等多种非生物胁迫。例如,ABA、高盐和干旱胁迫下,ZmbZIP72在玉米Zea mays幼苗各器官上调表达,其异源过表达使拟南芥Arabidopsis thaliana显著改善叶片生理状态,通过减少失水和电解质渗漏来提高转基因株系的抗旱性和耐盐性[8]。高盐、干旱、高温、低温和ABA胁迫下,ZmbZIP4在玉米幼苗各器官被诱导表达,其异源过表达不仅增加拟南芥主根长和侧根数,还显著提高ABA合成,增强植株抵御非生物胁迫能力[9]。水稻Oryza sativa的OsbZIP42可正向调控ABA介导的信号通路,增强水稻过表达植株对干旱胁迫应答的敏感性[10]。bZIP还和ANAC096等转录因子互作,协同提高转基因植株抗旱性[11]。茶树Camellia sinensis的CsbZIP18是ABA信号和冷胁迫的负调控因子,异源过表达导致拟南芥对ABA信号敏感性下降、电解质渗漏升高以及光合效率下降,降低植株耐冻性[12]。bZIP还参与调控其他生物学过程,如组织器官发育[13−14]、盐胁迫[15]、激素和糖信号传递[16]、病虫害防御[17]、光反应和次生代谢物合成调控等[18−19]。
bZIP基因家族成员有进化多样性,成员数量在物种间存在显著差异,例如,拟南芥75个[20]、番茄Solanum lycopersicum 70个[21]、水稻89个[22]、玉米125个[23]、毛果杨Populus trichocarpa 214个[24],而闽楠bZIP成员数量不详,极大限制了闽楠bZIP基因功能研究。因此,本研究对闽楠bZIP转录因子家族成员进行全基因组鉴定,分析其蛋白理化性质,比较进化关系以及ABA处理下的表达水平,为解析bZIP家族成员的基因功能及其在逆境胁迫的调控机制提供理论基础。
-
从PFAM数据库获取包含保守结构域(PF00170、PF07716)的隐马尔科夫蛋白模型。运用HMMER软件在闽楠基因组[25]筛选bZIP同源序列。将初步获得的基因序列提交SMART数据库和美国国家生物技术信息中心(NCBI)网站(
https://www.ncbi.nlm.nih.gov/ )确认结构域,手动去除冗余,获得PbbZIP基因成员。利用Protparam数据库(https://www.expasy.org/ )进一步分析PbbZIP蛋白理化性质,通过植物亚细胞定位(Plant-mPLoc)数据库(http://www.csbio.sjtu.edu.cn )进行亚细胞定位分析,通过Prabi数据库(https://npsa-prabi.ibcp.fr/ )分析PbbZIP蛋白的二级结构。 -
运用Jaview软件对ClustalW输出的多序列比对结果进行美化,基因保守结构域用Tbtools[26]进行可视化,MEME suite数据库(
https://meme-suite.org/ )分析蛋白序列的保守基序,利用GSDS工具(http://gsds.gao-lab.org/ )在线绘制PbbZIP基因结构。 -
从Ensembl数据库(
http://plants.ensembl.org/species.html )获取拟南芥、番茄、毛果杨基因组数据,通过HMMER软件挑选出bZIP同源基因,构建最大似然进化树(Bootstrap: 1 000次)。通过Evolview网站(http://www.evolgenius.info/evolview )美化系统进化树。用TBtools的MCScanX工具分析PbbZIP基因的共线性关系。 -
在浙江农林大学苗圃地挑选长势健康的2年生闽楠半同胞家系WY8苗,选用2 mmol·L−1ABA均匀喷洒整株苗,使土壤表面湿润,吐温80 (Tween 80)为对照组(ck)。采用取样时间倒推法,分别在处理1、3、12、24、48和72 h取叶片和根,均在11:00取样,每个时间段包含15株闽楠苗,每5株混合作为1个生物学重复,3次重复,液氮速冻后储存于−80 ℃冰箱。
-
从闽楠基因组[25]提取PbbZIP基因编码区(CDS)上游2 000 bp序列,提交至PlantCARE数据库分析启动子顺式作用元件,统计与逆境胁迫相关的作用元件,对ABA响应元件较多的PbbZIP基因进行实时荧光定量,并分析其响应表达模式。
-
用CTAB法提取高质量总RNA,经质量分数为1%的琼脂糖凝胶电泳和NanoDrop检测合格后用于后续研究。每份RNA取1 μg,按照Vazyme公司提供的HiScript® II Reverse Transcriptase试剂盒反转录合成高质量cDNA,用ddH2O稀释5倍待用。使用NCBI 设计PbbZIP基因特异性引物用于实时荧光定量PCR (RT-qPCR)反应。使用RT-qPCR反应酶配成10.00 μL标准反应体系,包含1.00 μL cDNA、0.25 μL上下游引物、5.00 μL反应液和3.50 μL ddH2O。两步法RT-qPCR反应程序:95 ℃ 30 s,95 ℃ 10 s,60 ℃ 30 s,40个循环。闽楠ef1α基因(登录号:KX682032)作为内参基因,用2−ΔΔCt法计算各基因相对表达量[27],并通过SPSS进行方差分析及多重比较。
-
利用HMMER软件从闽楠基因组初步获得78条蛋白序列,经SMART和NCBI数据库验证结构域完整性,最终鉴定出63个PbbZIP成员,按染色体位置命名为PbbZIP01~PbbZIP63,并分析其蛋白特征和理化性质。PbbZIP蛋白长度介于110 (PbbZIP60)~835 (PbbZIP12)个氨基酸,相对分子量为13.05~88.65 kDa,平均为39.76 kDa。蛋白等电点为4.48 (PbbZIP02)~11.95 (PbbZIP32),疏水性为−1.19 (PbbZIP58)~−0.19 (PbbZIP16)。亚细胞定位分析结果表明:59个成员定位于细胞核,而PbbZIP02定位于细胞质,PbbZIP05和PbbZIP12定位于内质网,PbbZIP16定位于线粒体。
-
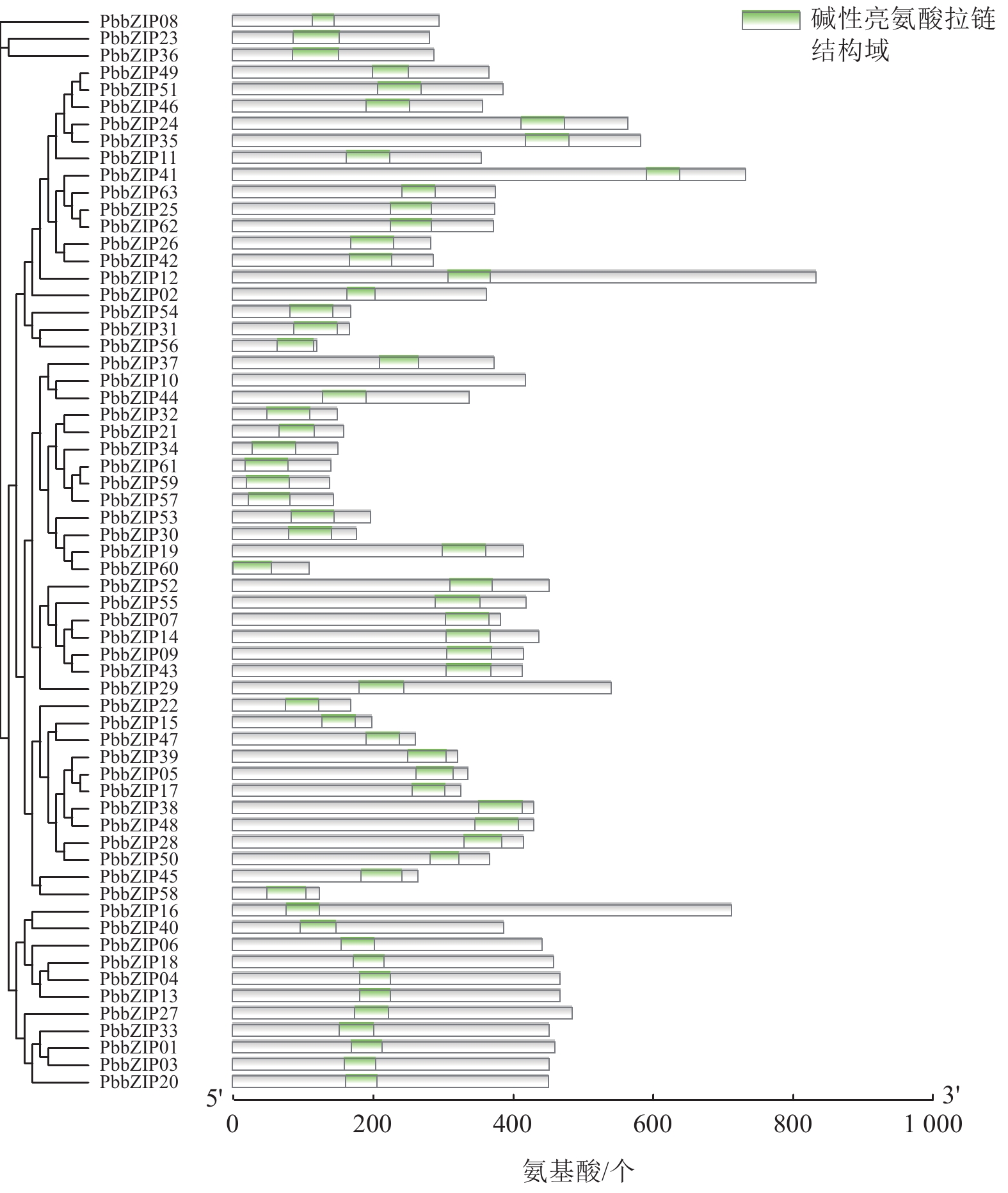
Motif分析结果表明:PbbZIP基因保守基序较丰富,且不同亚族差异明显。Motif 5和Motif 1在PbbZIP基因家族广泛存在(图1),分别为碱性结构区域和亮氨酸拉链结构区域,碱性区域可以和DNA特异性结合,发挥调控活性(图2)。A亚族特有Motif 6、Motif 10和Motif 13;C亚族和S亚族特有Motif 11;D亚族的保守基序较丰富,特有基序包括Motif 2、Motif 3、Motif 4、Motif 7、Motif 9和Motif 12;F亚族特有Motif 15;I亚族特有Motif 6;G亚家族特有Motif 8和Motif 14。其中,A、E、F、G、J亚族含有重复基序。
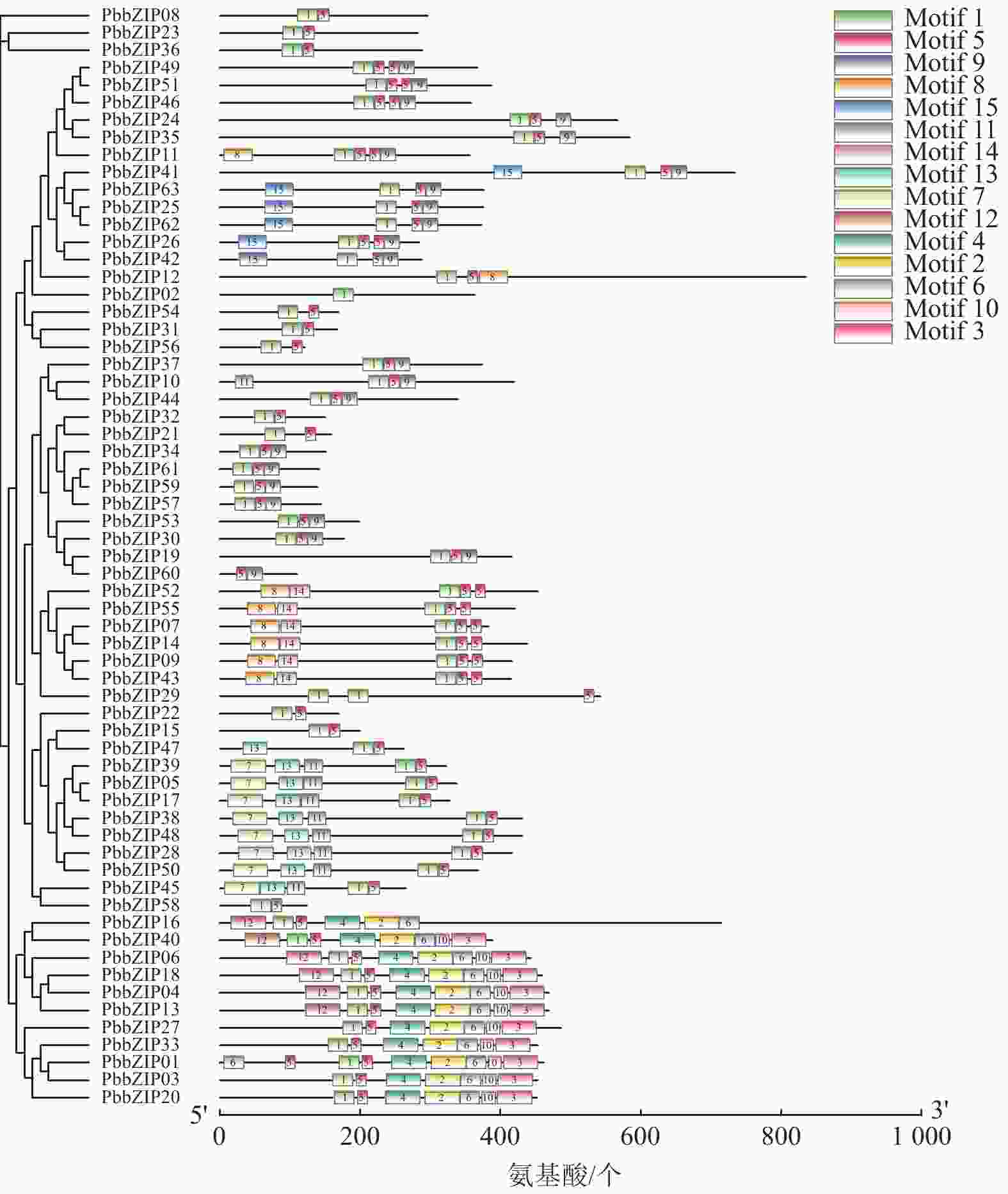
图 1 闽楠bZIP家族成员的保守基序
Figure 1. Conserved motifs of PbbZIP family members
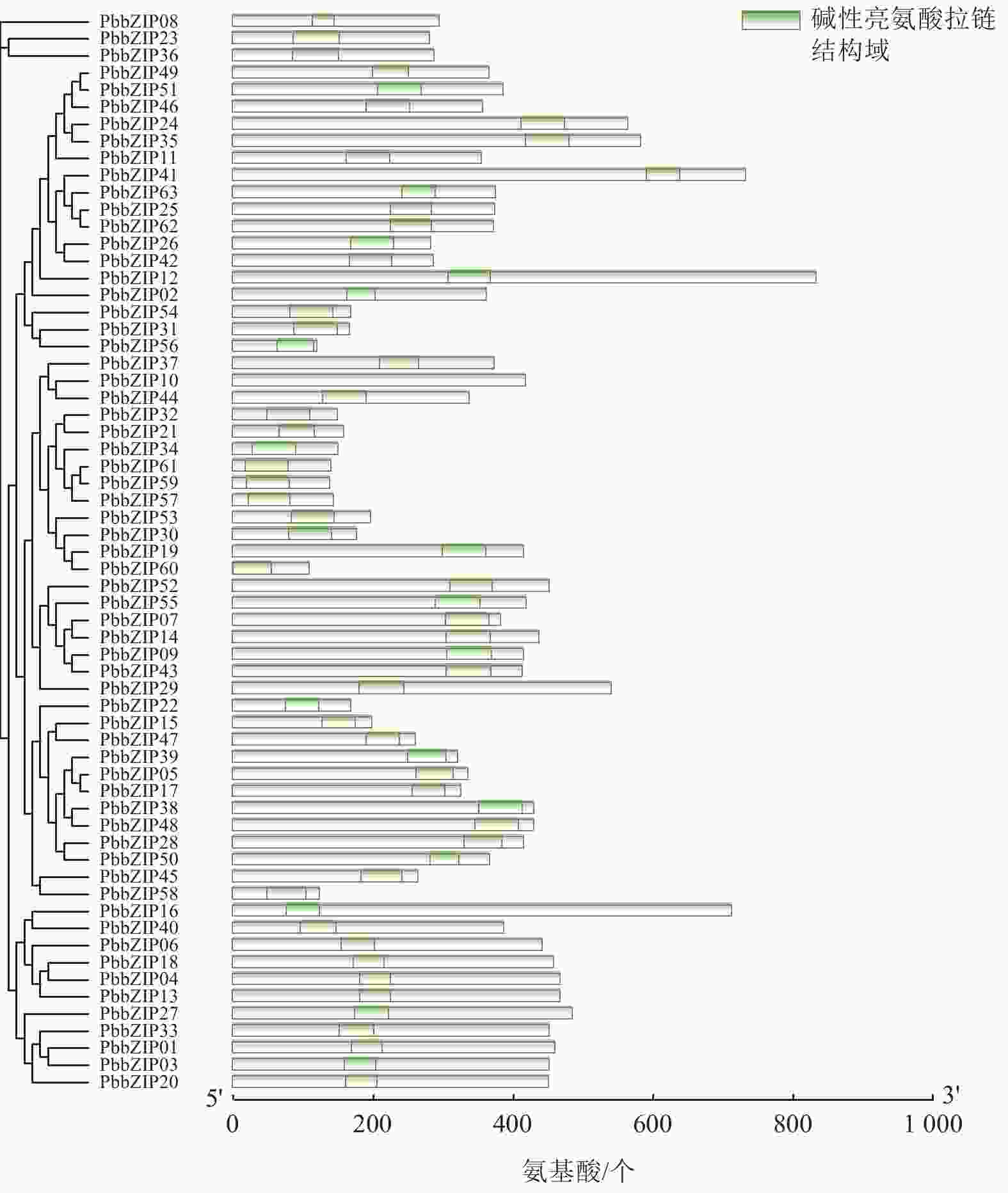
图 2 闽楠bZIP家族成员的结构域
Figure 2. Domains of PbbZIP family members
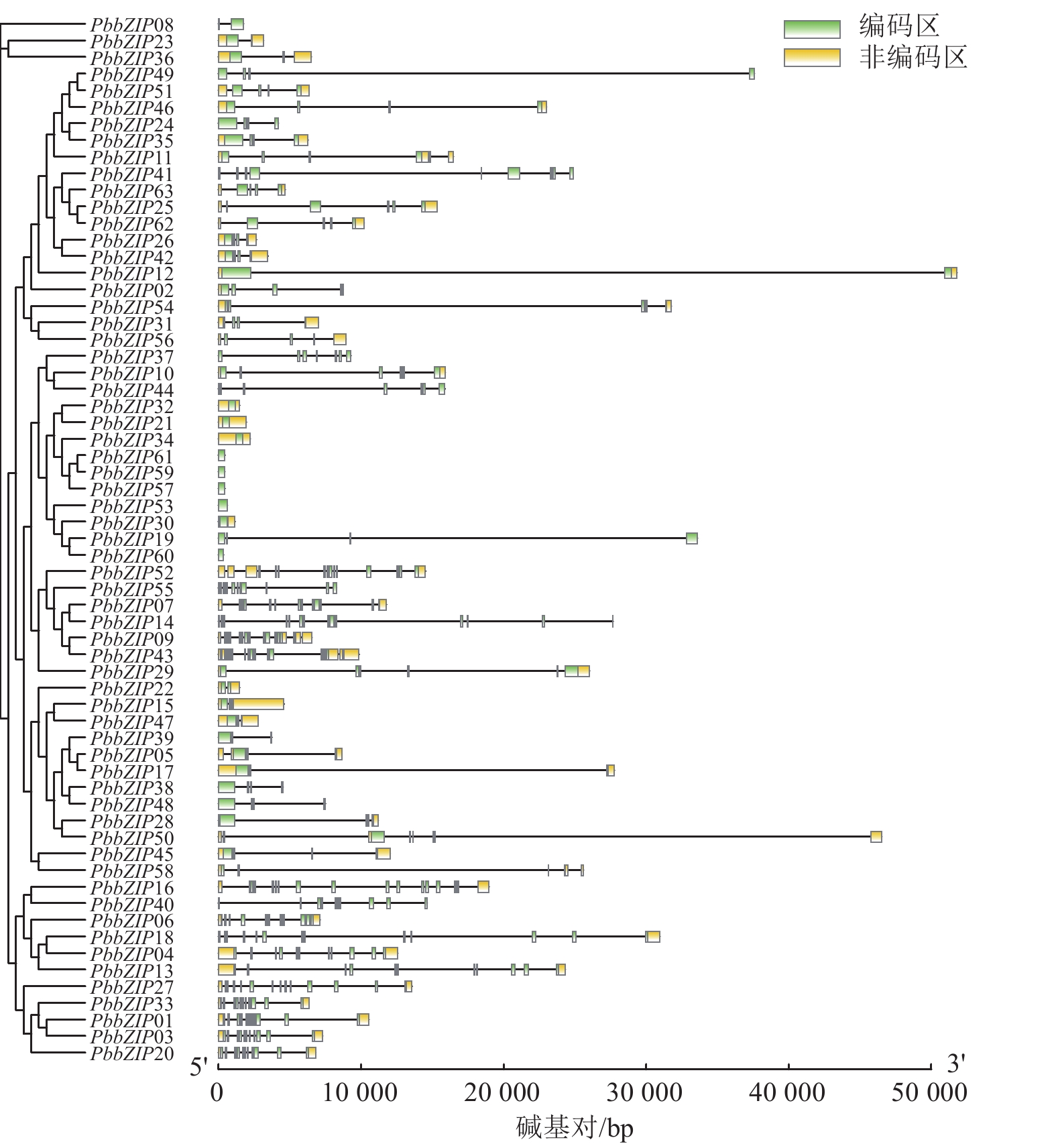
PbbZIP家族的基因结构在内含子、外显子的数量和位置存在显著多样性和差异性(图3)。例如,S亚族外显子数量最少(1~4个),且多数无内含子,这与拟南芥bZIP-S亚族结构相似;A亚族含有3~4个外显子,C亚族含有6~7个外显子,D亚族含有11~12个外显子;E亚族含有5~6个外显子,F亚族含有2~3个外显子,而G亚族内含子和外显子最多,为13~14个。
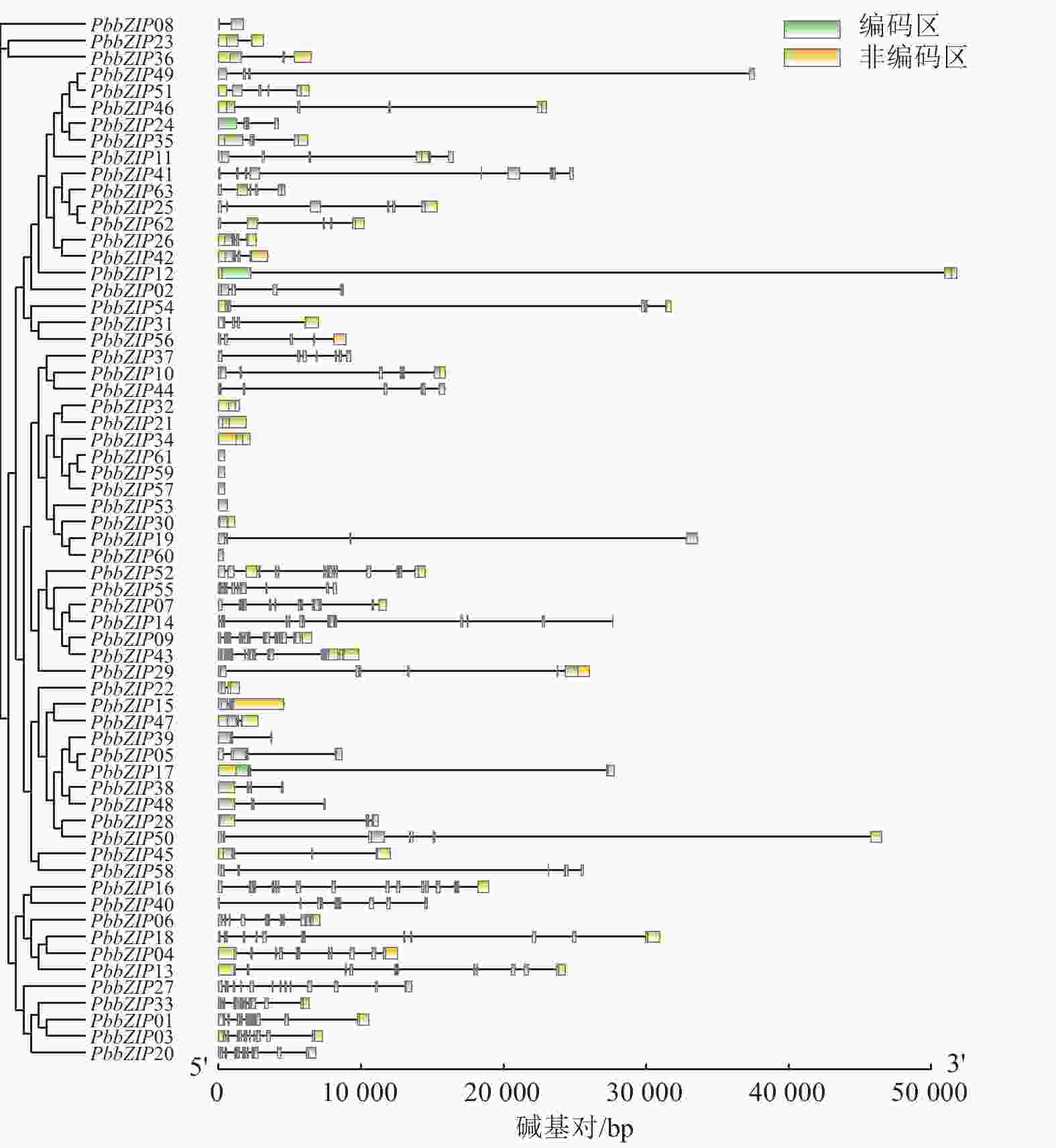
图 3 闽楠bZIP家族成员的基因结构
Figure 3. Gene structures of PbbZIP family members
-
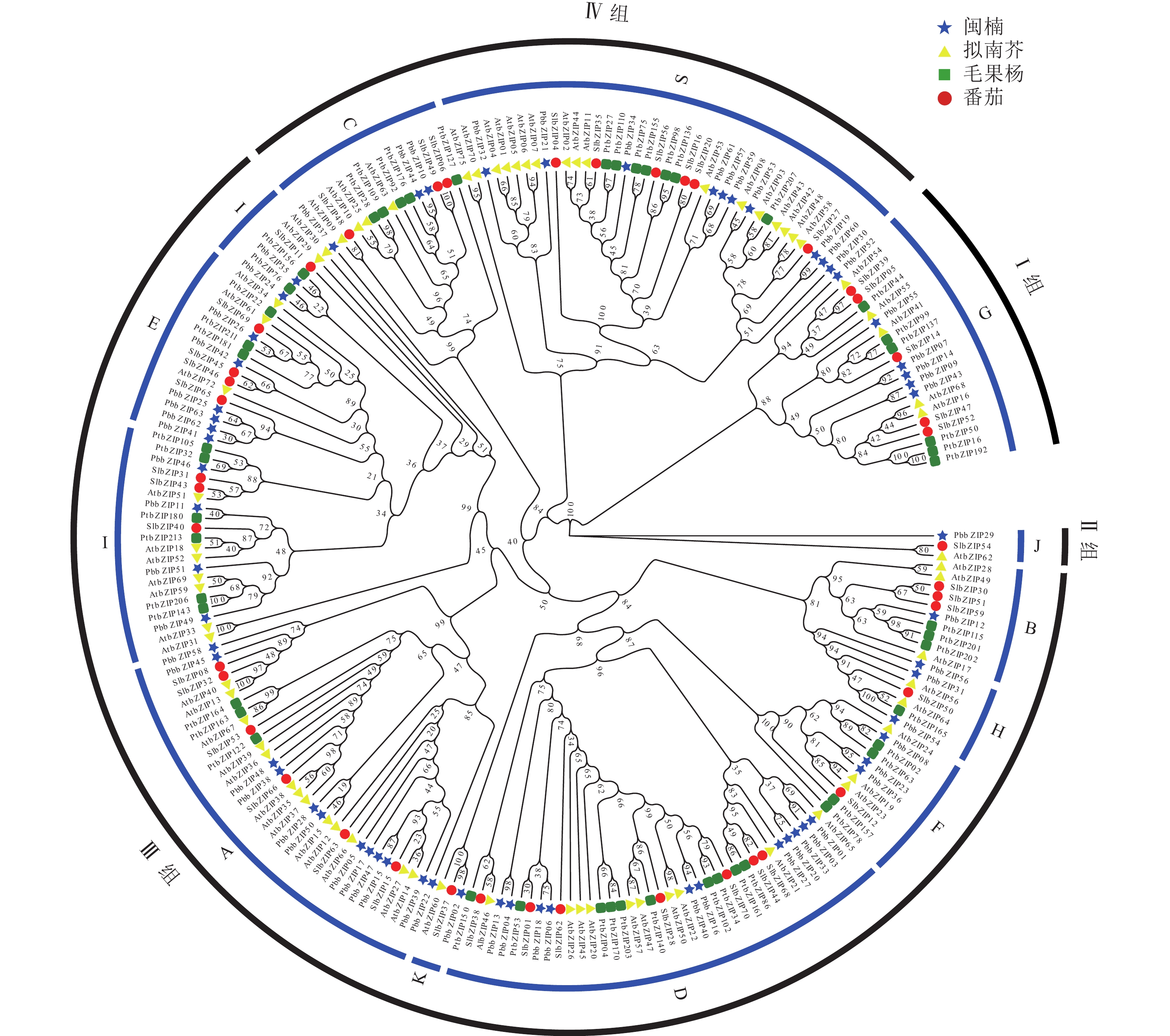
为研究bZIP基因的进化关系,选取拟南芥、番茄、毛果杨bZIP氨基酸序列与PbbZIP基因构建最大似然进化树(图4)。系统进化分析表明:63个PbbZIPs基因按序列同源性划分为4组12亚族,Ⅰ 组(G)、Ⅱ 组(J)、Ⅲ 组(B、H、E、I、F、K、A、D)和Ⅳ 组(C、S),其中A亚族12个、B亚族1个、C亚族3个、D亚族11个、E亚族5个、F亚族3个、G亚族6个、H亚族4个、I亚族6个、J亚族1个、K亚族1个和S亚族10个。
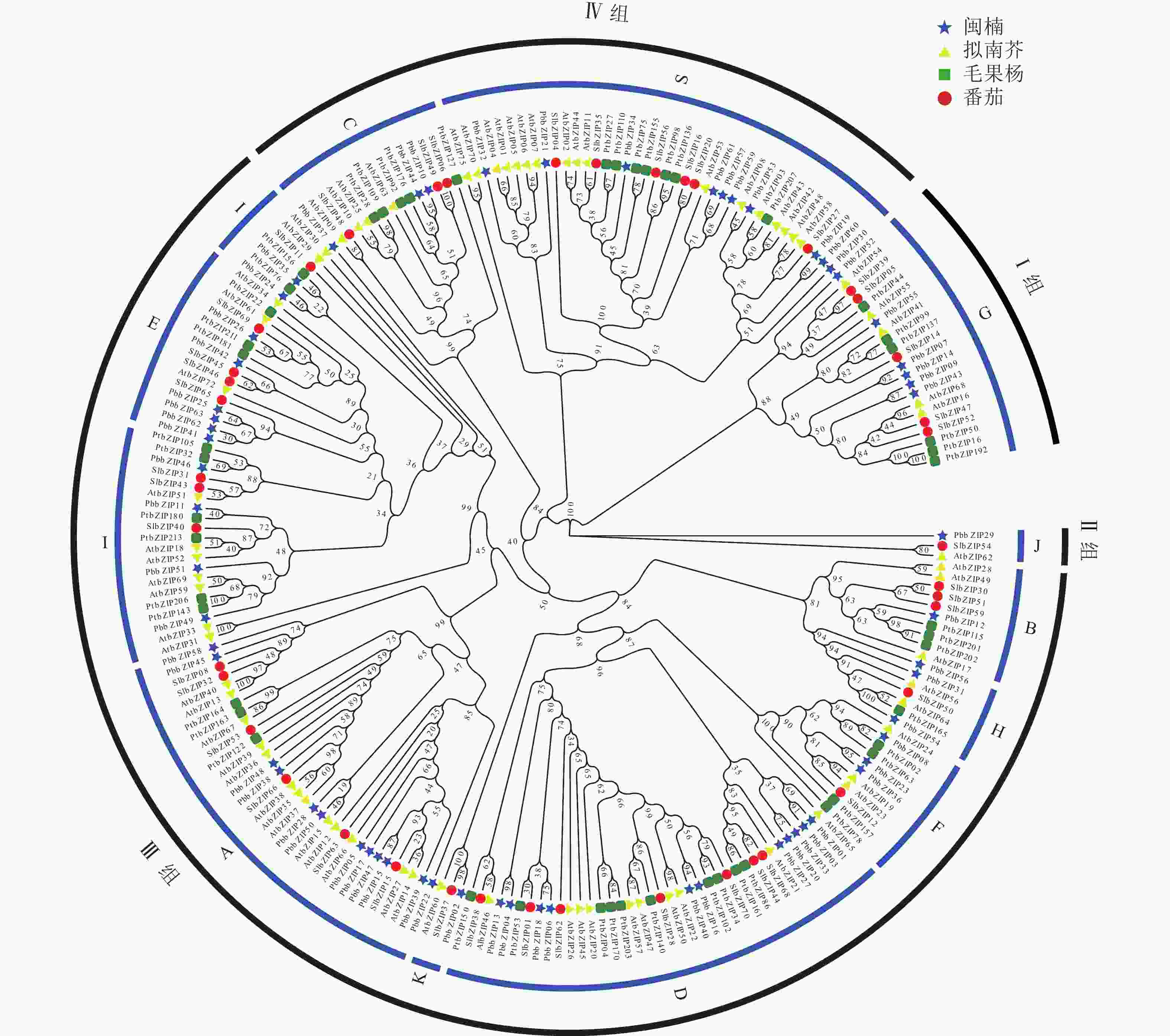
图 4 闽楠、拟南芥、毛果杨和番茄bZIP家族系统进化分析
Figure 4. Phylogenetic analysis of bZIP family from P. bournei, P. trichocarpa, A. thaliana, and S. lycopersicum
-
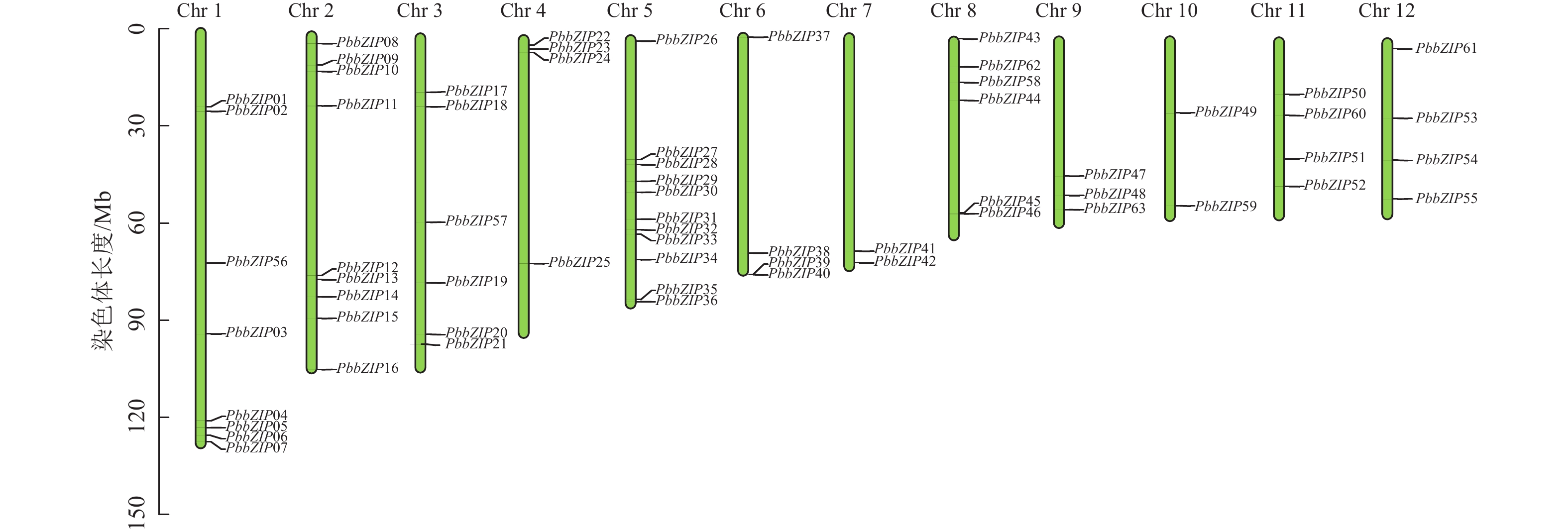
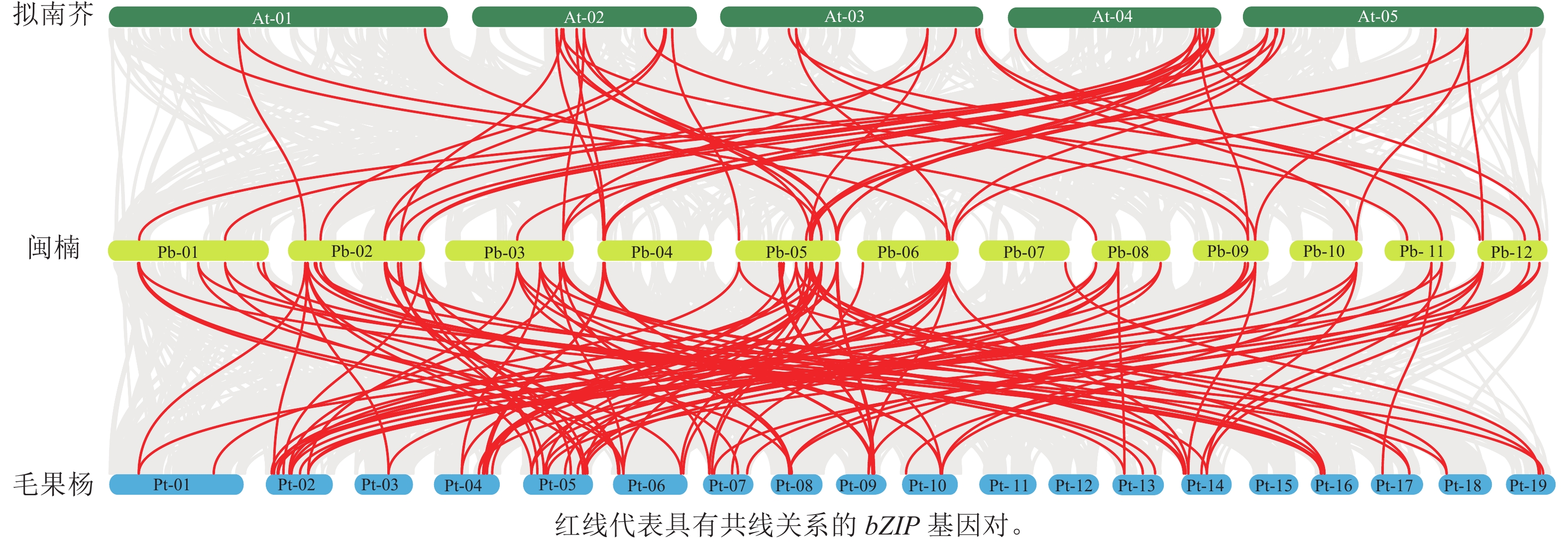
染色体定位结果表明:63个PbbZIPs基因不均匀分布于12条染色体(图5),其中Chr 5基因数量最多,含11个PbbZIPs成员,占17.46%,而Chr 7和Chr 10最少,仅有2个PbbZIPs成员。为深入了解PbbZIP基因家族的扩张模式,对63个PbbZIPs基因进行共线性分析,结果表明:分布于12条染色体上的27对PbbZIPs基因存在片段复制,其中D亚族共线基因对最多(10对),未检测到串联复制,表明片段复制是PbbZIP基因家族扩张的主要模式。不同物种间共线性结果表明(图6):闽楠与拟南芥47个、毛果杨110个bZIPs基因对存在共线性关系,闽楠和毛果杨bZIP基因更保守,亲缘关系较近。
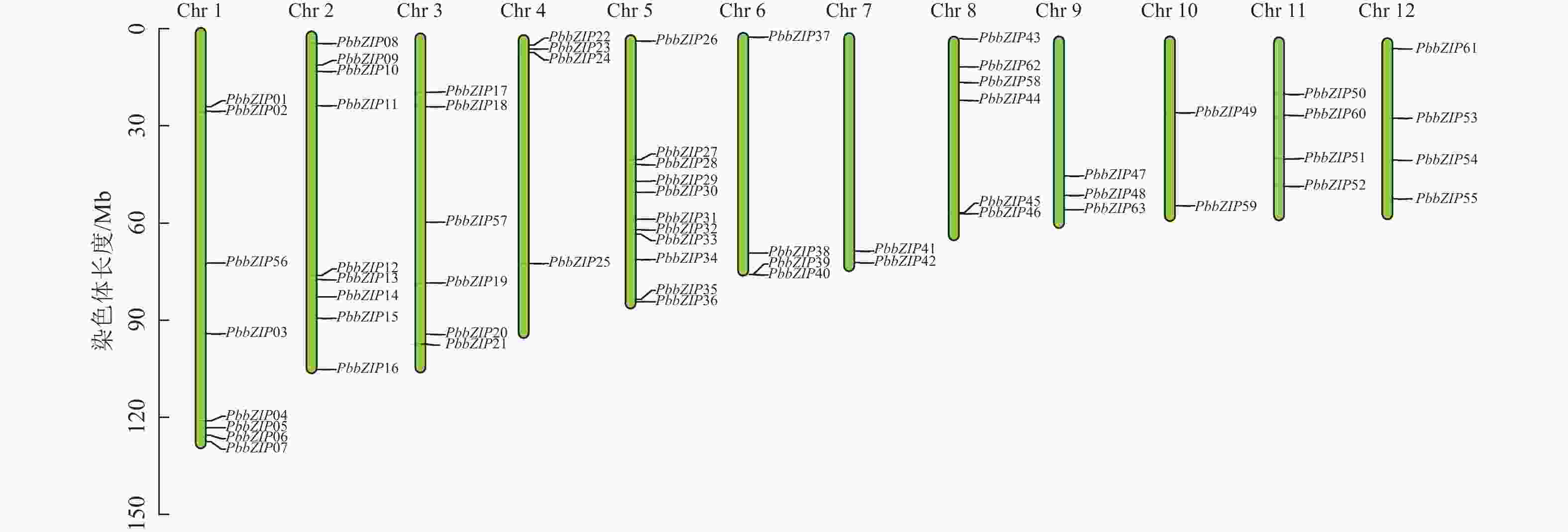
图 5 PbbZIP家族成员染色体定位
Figure 5. Chromosomal localization of PbbZIP family members

图 6 闽楠bZIP家族成员种间共线性分析
Figure 6. Interspecific collinear relationships of PbbZIP family members
-
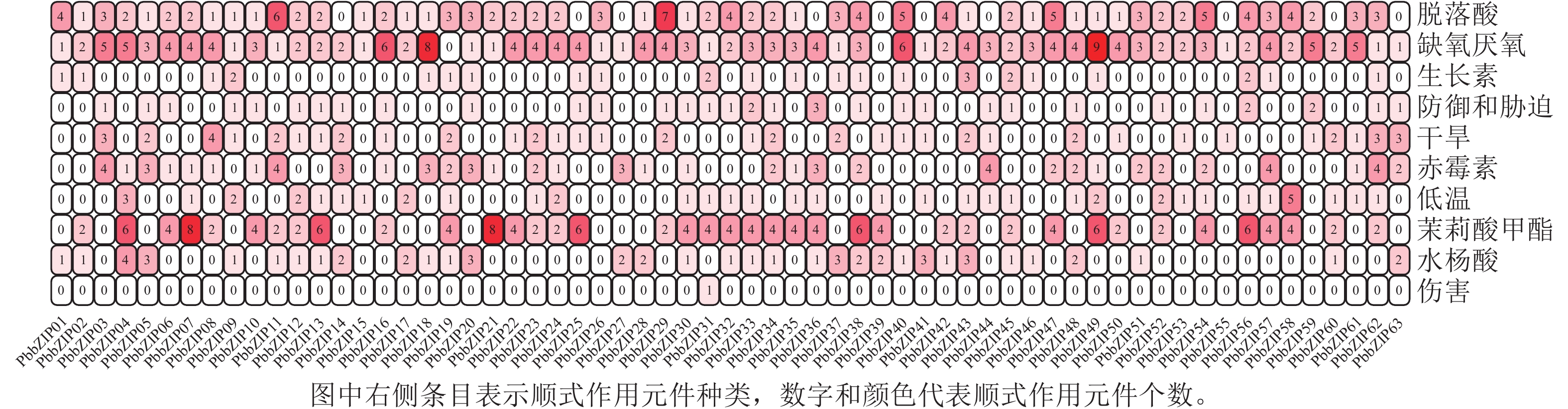
PbbZIP基因CDS区上游启动子顺式作用元件分析结果表明(图7):共发现18种(745个)与胁迫相关的顺式作用元件,其中激素响应元件428个,包括ABA响应元件(ABRE) 129个,茉莉酸甲酯响应元件(TGACG-motif、CGTCA-motif) 146个,赤霉素响应元件(P-box、GARE-motif、TATC-box) 71个,水杨酸响应元件(SARE、TCA-element) 54个,生长素响应元件(TGA-element、TGA-box、AuxRR-core) 28个。此外,还发现多种非生物胁迫响应元件,如低温响应元件(LTR、DRE) 45个,干旱响应元件(MBS) 53个,缺氧厌氧元件(ARE、GC-motif) 182个,防御和胁迫响应元件(TC-rich repeats) 36个,伤害响应元件(WUN-motif) 1个。
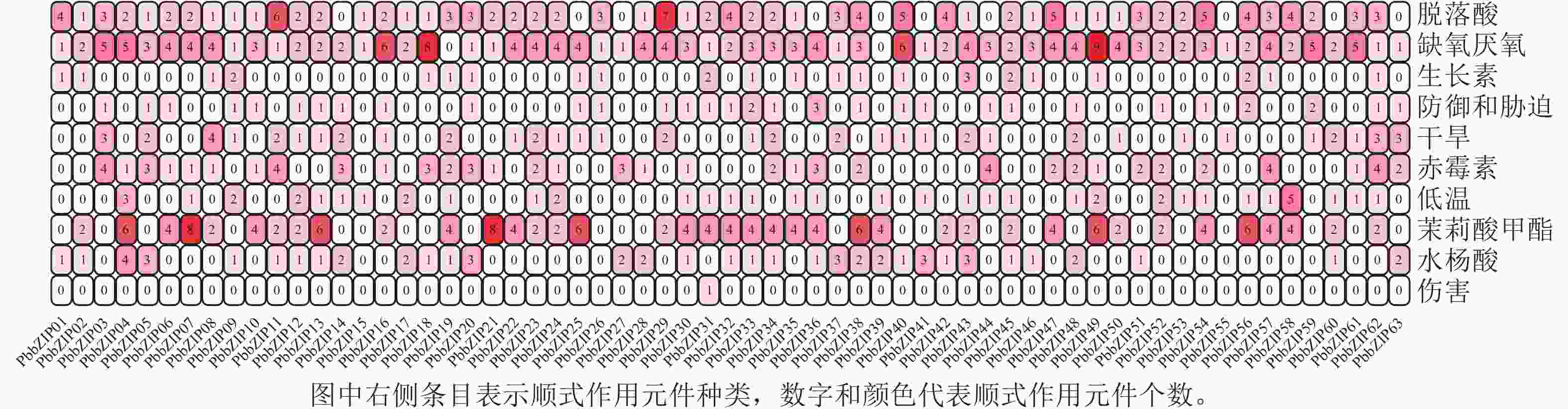
图 7 PbbZIPs与逆境胁迫相关的顺式作用元件
Figure 7. Cis-acting elements associated with adversity stress in promoters of PbbZIPs
-
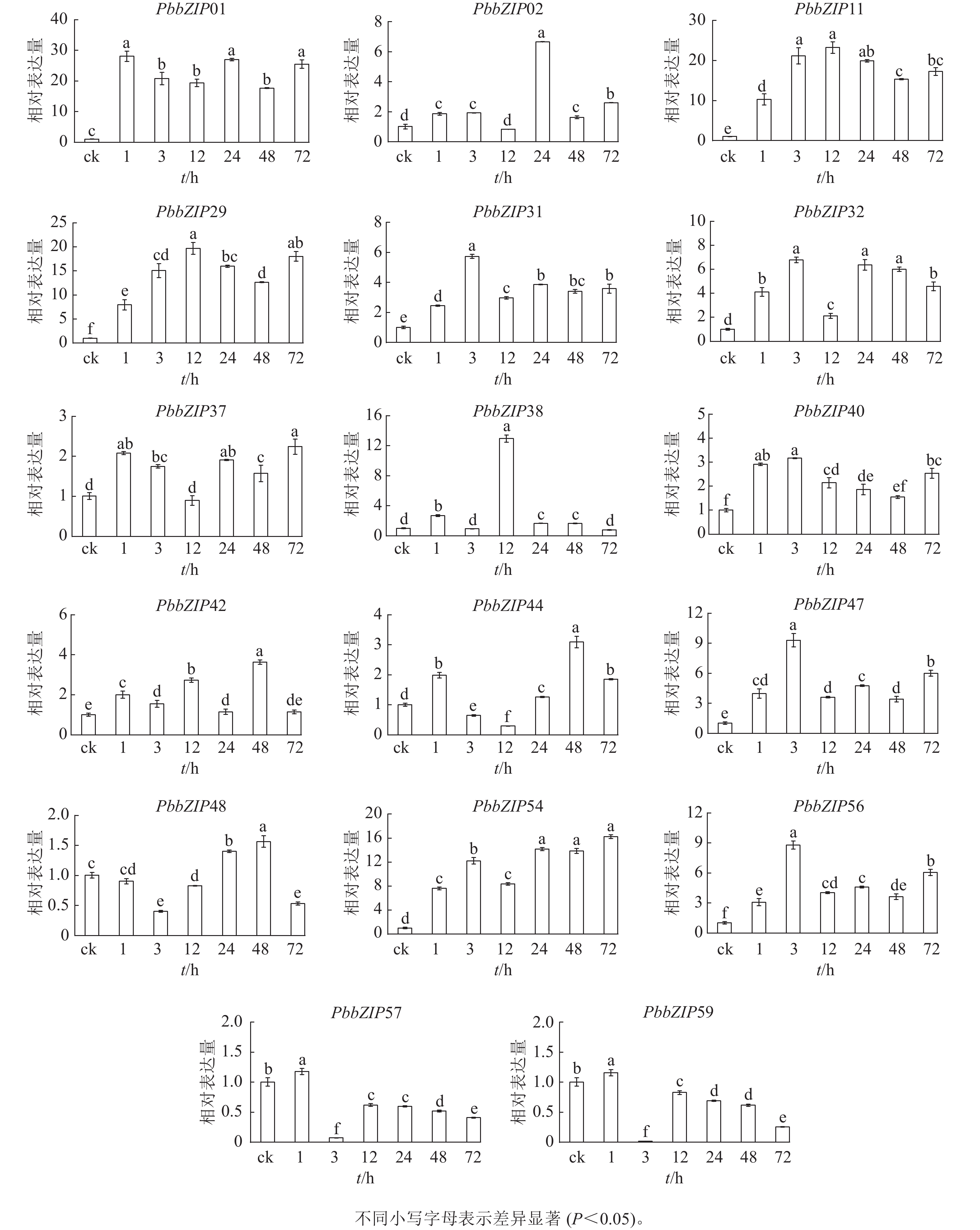
PbbZIP基因启动子区域富含ABA响应元件,挑选含较多ABA响应元件的17个PbbZIPs成员进行ABA处理下的表达模式分析。首先,应用RT-qPCR分析ABA处理后叶中17个PbbZIPs的响应程度,表明叶中17个PbbZIPs对ABA信号有着不同程度的诱导响应(图8)。例如,PbbZIP01、PbbZIP11、PbbZIP29和PbbZIP54等4个基因在叶中不同处理时间均被显著诱导(P<0.05),相对表达倍数高于8倍;PbbZIP31、PbbZIP32、PbbZIP37、PbbZIP40、PbbZIP42、PbbZIP47和PbbZIP56等7个成员均被ABA诱导表达,但表达倍数增加幅度不高;PbbZIP44、PbbZIP48、PbbZIP57和PbbZIP59在ABA处理3 h或12 h时被显著抑制表达(P<0.05);PbbZIP02和PbbZIP38分别仅在ABA处理24 h和12 h强烈诱导表达,而其他处理时间段基本不被诱导。
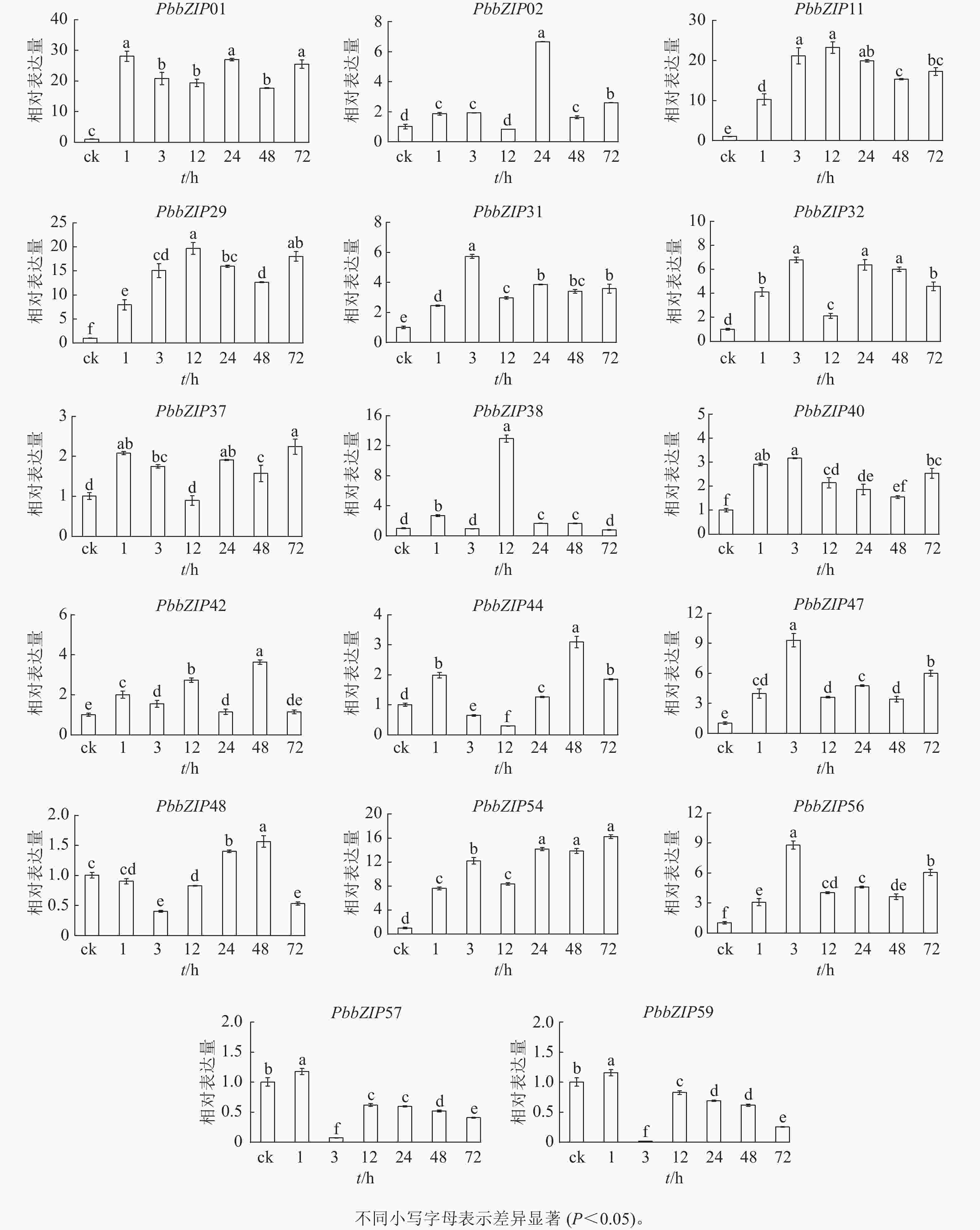
图 8 叶中PbbZIP基因对脱落酸处理的表达响应
Figure 8. Response of PbbZIP genes to ABA treatment in leaves
-
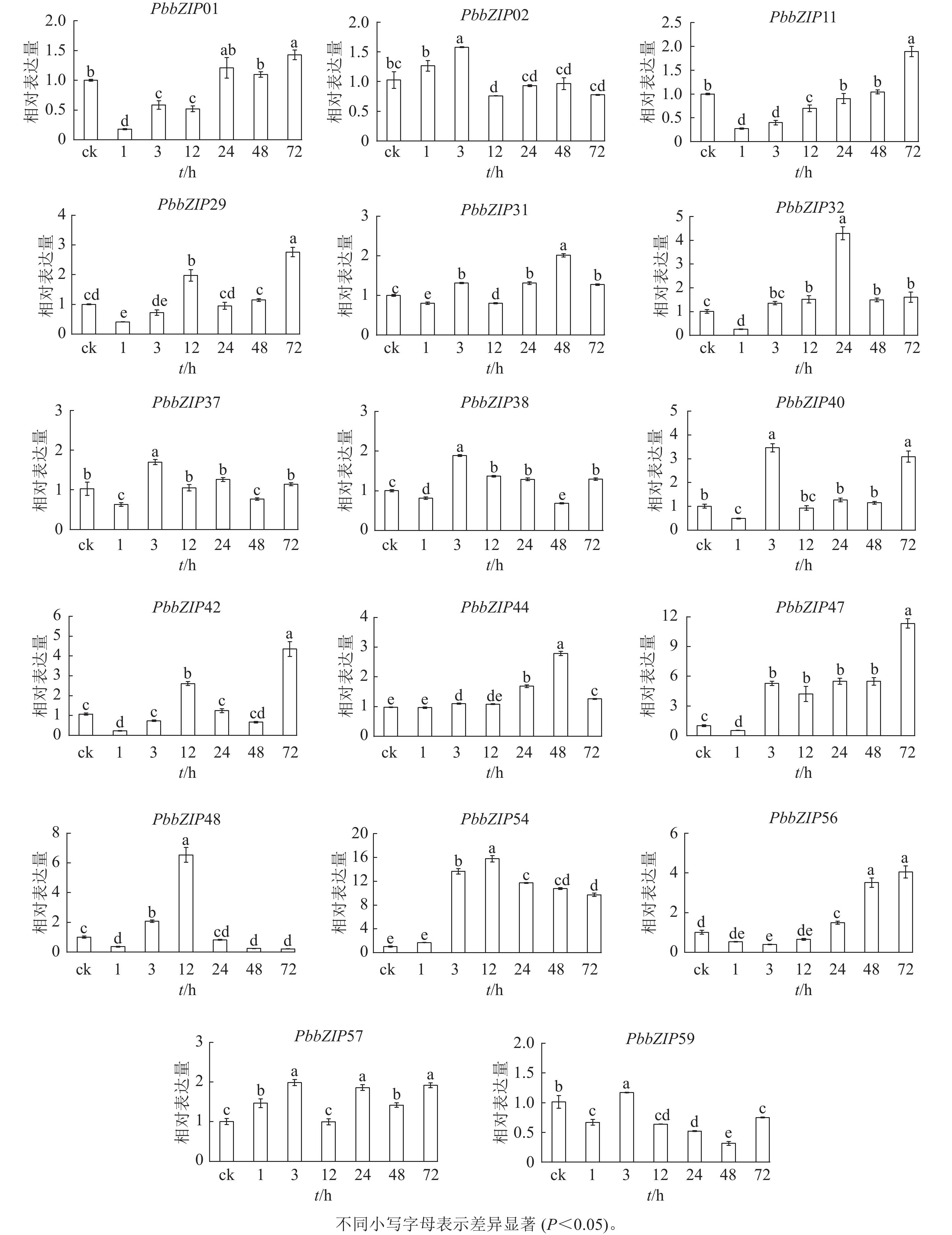
ABA处理1~72 h,17个PbbZIP基因在根中表现出不同趋势,且表达倍数较叶片低(图9)。PbbZIP01、PbbZIP11、PbbZIP29、PbbZIP31、PbbZIP32、PbbZIP37、PbbZIP38、PbbZIP40、PbbZIP42、PbbZIP47、PbbZIP48、PbbZIP56和PbbZIP59等表达水平先降低后升高,其中PbbZIP32、PbbZIP37、PbbZIP38、PbbZIP40、PbbZIP48和PbbZIP59等基因的表达量在ABA处理48 h后降低,而PbbZIP37、PbbZIP38、PbbZIP40和PbbZIP59表达水平在ABA处理72 h后显著升高(P<0.05)。PbbZIP02、PbbZIP44和PbbZIP57表达水平受ABA影响较小;PbbZIP54在ABA处理3 h后显著诱导表达(P<0.05),且保持较高水平,可能参与ABA信号响应发挥重要调控功能。
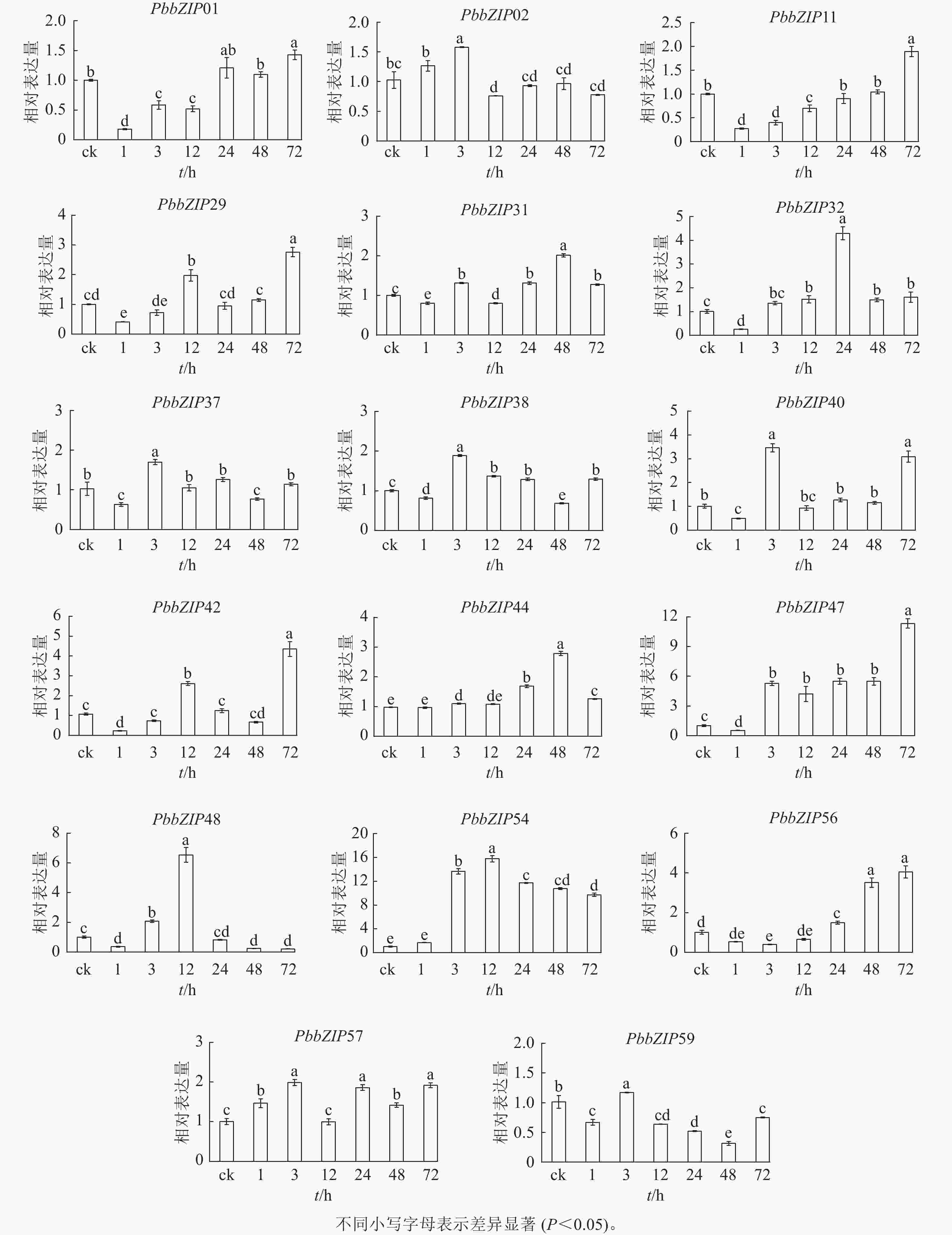
图 9 根中PbbZIP基因对ABA处理的表达响应
Figure 9. Response of PbbZIP genes to ABA treatment in roots
-
bZIP基因家族是成员和功能最复杂的转录因子家族之一,参与多种代谢过程,目前功能研究集中于拟南芥、毛果杨、水稻和番茄等物种。本研究从闽楠基因组鉴定出63个PbbZIP转录因子,该家族成员在蛋白理化性质、基因结构和保守结构域等方面与已报道物种(拟南芥[20]、毛果杨[24]、番茄[21]等)较相似,说明bZIP基因结构较保守。进一步将PbbZIP与拟南芥、番茄和毛果杨构建最大似然进化树,发现63个PbbZIP基因可进一步划分为12亚族,可能由4个以上不同功能的原始bZIP基因进化扩张产生[28],而在其他物种中,bZIP转录因子被划分为不同亚族,如拟南芥10个[20]、毛果杨12个[29]、烟草Nicotiana tabacum 11个[30]。其中PbbZIP29基因结构高度分化,单独聚类在一个进化分支上,其功能是否分化有待后续深入研究。
通过启动子顺式作用元件分析发现:PbbZIP基因CDS上游启动子区域存在多种响应元件,推测PbbZIP基因可能受生物及非生物胁迫诱导表达。例如ABA、茉莉酸甲酯和生长素等激素响应元件在启动子区域数量较多,推测PbbZIP基因可能受到激素诱导。低温、干旱和缺氧厌氧诱导等非生物胁迫响应元件数目也较多,推测PbbZIP表达可能受非生物胁迫诱导,正向或负向调控次生代谢途径而参与植物生长发育进程。例如,猕猴桃Actinidia deliciosa bZIP12正向调控ABA介导的AchnKCS酶基因,促进果实创伤愈合,免受病原体侵害[31]。在ABA、干旱、盐胁迫下,辣椒Capsicum annuum叶片bZIP-D亚族中的CaDILZ1显著上调,它通过调节ABA含量、气孔开合度及抗旱基因表达水平来增强转基因植株的耐旱性[32]。
17个PbbZIP基因在叶和根有不同程度的诱导表达,随ABA诱导表现出不同表达趋势。处理1~72 h,叶中PbbZIP11等基因和根中PbbZIP44等基因表达量先升高后降低,而叶中PbbZIP37等基因以及根中PbbZIP56基因表达趋势与之相反,先下降后上升,烟草bZIP-A亚族基因经1 μmol·L−1ABA处理24 h后也有相同趋势[33];武立伟等[34]研究发现:25和50 mg·L−1 ABA处理甘草Glycyrrhiza uralensis根,GubZIP1~GubZIP56响应趋势与本研究结果相似,表明PbbZIP基因可能参与ABA等非生物胁迫的响应;孙晓丽等[35]研究发现:拟南芥AtbZIP1与ABRE元件结合负向调节植株对ABA敏感性,AtbZIP1缺失提高了种子萌发率,增加了主根长和ABA响应基因的表达,推测PbbZIP基因可能有相似功能。叶中PbbZIP01、PbbZIP29、PbbZIP32、PbbZIP38和PbbZIP54等被ABA显著诱导,根中PbbZIP47和PbbZIP54被显著诱导,表达量较高,后续需进一步开展功能验证,阐明其生物学功能与调控机制。
-
本研究通过对闽楠bZIP转录因子家族鉴定和表达分析,发现63个基因家族成员高度保守,不同亚家族在蛋白理化性质、基因结构、染色体定位、保守基序等方面表现出进化多样性和差异性。PbbZIP基因的表达随ABA诱导时间增加呈非同步变化的趋势,具有一定组织特异性,叶响应程度普遍高于根,这种表达特征与闽楠胁迫耐受性高度关联,存在复杂的调控网络,需进一步探索PbbZIP基因功能。
Identification and expression analysis of bZIP gene family under ABA treatment in Phoebe bournei
-
摘要:
目的 对闽楠Phoebe bournei bZIP (PbbZIP)转录因子家族成员鉴定,分析其对脱落酸(ABA)信号的响应水平。 方法 基于生物信息学方法,对PbbZIP基因家族进行了全基因组鉴定,分析其蛋白理化特性、基因结构、进化关系、启动子顺式作用元件和ABA处理下的表达分析。 结果 从闽楠12条染色体共鉴定出63个PbbZIPs基因,分为12亚族,不同亚族基因结构和基序差异显著,但同亚族高度保守。PbbZIP基因多数定位在细胞核,其编码蛋白长度为110~835个氨基酸,等电点为4.48~11.95,疏水性为−1.19~−0.19。分布于12条染色体的27对PbbZIPs基因存在共线性关系,是PbbZIP基因家族扩张的主要模式。PbbZIP基因上游启动子区域存在多种与非生物胁迫相关的作用元件,其中ABA、水杨酸、茉莉酸甲酯的响应元件较多。基因表达分析结果表明:2 mmol·L−1ABA处理闽楠1~72 h,17个PbbZIPs基因在叶和根中被ABA信号不同程度地诱导表达,普遍上调,且根基因表达水平普遍低于叶片。 结论 63个PbbZIPs基因序列高度保守,不同亚族间基因结构、染色体定位和保守基序有进化多样性和差异性;叶和根中PbbZIP基因不同程度地响应ABA信号,参与调控其他非生物过程。图9参35 Abstract:Objective This study is aimed to identify the bZIP transcription factor family members from Phoebe bournei and investigate the response levels of its members to abscisic acid (ABA) treatment. Method A bioinformatic method was employed to identify the PbbZIPs family throughout the whole genome and to analyze its physicochemical properties, gene structure, evolutionary relationships, cis-acting elements in promoter, and the expression patterns under ABA treatment by RT-qPCR. Result A total of 63 PbbZIP genes were identified from 12 chromosomes in P. bournei, divided into 12 subfamilies with significantly different in gene structure and motifs, but highly conserved in the same subfamily. Most of the PbbZIPs were localized in the nucleus, and their encoded protein were 110 − 835 amino acid in length, −1.19 − −0.19 in hydrophobicity, and 4.48 − 11.95 in isoelectric point. The 27 pairs of PbbZIPs distributed on 12 chromosomes were featured with collinearity existence, which was the main pattern of PbbZIPs family expansion. A variety of abiotic stress-related action elements were found in the upstream promoter region of PbbZIPs, among which ABA, salicylic acid and methyl jasmonate were more abundant response elements and the genes expression of RT-qPCR revealed that 17 PbbZIPs were induced differentially by ABA signals and generally up-regulated in leaves and roots when P. bournei was treated with 2 mmol·L−1ABA for 1 − 72 h, with the relative expression of PbbZIPs in roots being generally lower than that in leaves. Conclusion The 63 PbbZIPs identified from the P. bournei genome were unevenly distributed across the 12 chromosomes and highly similar in gene sequences, whereas the chromosomal localization genetic structure and conserved motifs were evolutionarily diverse and different between subgroups. The PbbZIPs in leaves and roots responded differently to ABA treatment and got involved in the regulation of other abiotic processes. [Ch, 9 fig. 35 ref.] -
Key words:
- Phoebe bournei /
- bZIP family /
- systematic evolution /
- ABA /
- expression analysis
-
图 4 闽楠、拟南芥、毛果杨和番茄bZIP家族系统进化分析
Figure 4 Phylogenetic analysis of bZIP family from P. bournei, P. trichocarpa, A. thaliana, and S. lycopersicum
图 6 闽楠bZIP家族成员种间共线性分析
Figure 6 Interspecific collinear relationships of PbbZIP family members
图 7 PbbZIPs与逆境胁迫相关的顺式作用元件
Figure 7 Cis-acting elements associated with adversity stress in promoters of PbbZIPs
图 8 叶中PbbZIP基因对脱落酸处理的表达响应
Figure 8 Response of PbbZIP genes to ABA treatment in leaves
-
[1] 丁鑫, 肖建华, 黄建峰, 等. 珍贵木材树种楠木的野生资源调查[J]. 植物分类与资源学报, 2015, 37(5): 629 − 639. DING Xin, XIAO Jianhua, HUANG Jianfeng, et al. Investigation on wild resource of valuable timber tree Phoebe zhennan (Lauraceae) [J]. Plant Diversity and Resources, 2015, 37(5): 629 − 639. [2] 葛永金, 王军峰, 方伟, 等. 闽楠地理分布格局及其气候特征研究[J]. 江西农业大学学报, 2012, 34(4): 749 − 753, 761. GE Yongjin, WANG Junfeng, FANG Wei, et al. Distribution pattern of Phoebe bournei (Hemsl.) Yang and the characteristics of climate [J]. Acta Agriculturae Universitatis Jiangxiensis, 2012, 34(4): 749 − 753, 761. [3] 李娟, 欧汉彪, 林建勇, 等. 我国楠属种质资源分布现状及主要种特征差异[J]. 广西林业科学, 2020, 49(1): 54 − 59. LI Juan, OU Hanbiao, LIN Jianyong, et al. Distribution of Phoebe germplasm resources in China and characteristic differences among main species [J]. Guangxi Forestry Science, 2020, 49(1): 54 − 59. [4] 桑新华, 吴忠义, 黄丛林, 等. 植物逆境抗性相关转录因子的研究进展[J]. 植物学通报, 2004, 21(6): 700 − 708. SANG Xinhua, WU Zhongyi, HUANG Conglin, et al. Progresses on transcription factors related to plant stress-tolerance [J]. Chinese Bulletin of Botany, 2004, 21(6): 700 − 708. [5] 王金英, 丁峰, 潘介春, 等. 植物bZIP转录因子家族的研究进展[J]. 热带农业科学, 2019, 39(6): 39 − 45. WANG Jinying, DING Feng, PAN Jiechun, et al. Research progress of bZIP lineage transcription factors in plant [J]. Chinese Journal of Tropical Agriculture, 2019, 39(6): 39 − 45. [6] 崔荣秀, 张议文, 陈晓倩, 等. 植物bZIP参与胁迫应答调控的最新研究进展[J]. 生物技术通报, 2019, 35(2): 143 − 155. CUI Rongxiu, ZHANG Yiwen, CHEN Xiaoqian, et al. The latest research progress on the stress responses of bZIP involved in plants [J]. Biotechnology Bulletin, 2019, 35(2): 143 − 155. [7] 冉静, 邹杰, 刘爱玲, 等. 植物bZIP转录因子在非生物胁迫中的作用[J]. 湖南农业科学, 2012(7): 11 − 13. RAN Jing, ZOU Jie, LIU Ailing, et al. The role of plant bZIP transcription factors in abiotic stress [J]. Hunan Agricultural Sciences, 2012(7): 11 − 13. [8] YING Shen, ZHANG Dengfeng, FU Jing, et al. Cloning and characterization of a maize bZIP transcription factor, ZmbZIP72, confers drought and salt tolerance in transgenic Arabidopsis [J]. Planta, 2012, 235(2): 253 − 266. [9] MA Haizhen, LIU Can, LI Zhaoxia, et al. ZmbZIP4 contributes to stress resistance in maize by regulating aba synthesis and root development [J]. Plant Physiology, 2018, 178(2): 753 − 770. [10] JOO J, LEE Y H, SONG S I. OsbZIP42 is a positive regulator of ABA signaling and confers drought tolerance to rice [J]. Planta, 2019, 249(5): 1521 − 1533. [11] XU Zhengyi, KIM S Y, HYEON D Y, et al. The Arabidopsis NAC transcription factor ANAC096 cooperates with bZIP-type transcription factors in dehydration and osmotic stress responses [J]. The Plant Cell, 2013, 25(11): 4708 − 4724. [12] YAO Lina, HAO Xinyuan, CAO Hongli, et al. ABA-dependent bZIP transcription factor, CsbZIP18, from Camellia sinensis negatively regulates freezing tolerance in Arabidopsis [J]. Plant Cell Reports, 2020, 39(4): 553 − 565. [13] SHIN J, BUI D C, KIM S, et al. The novel bZIP transcription factor Fpo1 negatively regulates perithecial development by modulating carbon metabolism in the ascomycete fungus Fusarium graminearum [J]. Environmental Microbiology, 2020, 22(7): 2596 − 2612. [14] STRATHMANN A, KUHLMANN M, HEINEKAMP T, et al. BZI-1 specifically heterodimerises with the tobacco bZIP transcription factors BZI-2, BZI-3/TBZF and BZI-4, and is functionally involved in flower development [J]. The Plant Journal, 2001, 28(4): 397 − 408. [15] GAI Wenxian, MA Xiao, Qiao Yiming, et al. Characterization of the bZIP transcription factor family in pepper (Capsicum annuum L.): CabZIP25 positively modulates the salt tolerance [J/OL]. Frontiers in Plant Science, 2020, 11: 139[2023-05-20]. doi: 10.3389/fpls.2020.00139. [16] THALOR S K, BERBERICH T, LEE S S, et al. Deregulation of sucrose-controlled translation of a bZIP-type transcription factor results in sucrose accumulation in leaves [J/OL]. PLoS One, 2012, 7(3): e33111[2023-05-20]. doi: 10.1371/journal.pone.0033111. [17] NIGAL M C, ANTHONY D M G. Molecular and physiological aspects of nitrate uptake in plants [J]. Trends in Plant Science, 1998, 3(10): 389 − 395. [18] MOHANTY B, LAKSHMANAN M, LIM S, et al. Light-specific transcriptional regulation of the accumulation of carotenoids and phenolic compounds in rice leaves [J/OL]. Plant Signaling & Behavior, 2016, 11(6): e1184808[2023-05-20]. doi: 10.1080/15592324.2016.1184808. [19] PONTES L C G, CARDOSO C M Y, CALLEGARI D M, et al. A cassava CPRF-2-like bZIP transcription factor showed increased transcript levels during light treatment [J]. Protein &Peptide Letters, 2020, 27(9): 904 − 914. [20] JAKOBY M, WEISSHAAR B, DRÖGE-LASER W, et al. bZIP transcription factors in Arabidopsis [J]. Trends in Plant Science, 2002, 7(3): 106 − 111. [21] 朱芸晔, 薛冰, 王安全, 等. 番茄bZIP转录因子家族的生物信息学分析[J]. 应用与环境生物学报, 2014, 20(5): 767 − 774. ZHU Yunye, XUE Bing, WANG Anquan, et al. Comprehensive bioinformatic analysis of bZIP transcription factors in Solanum lycopersicum [J]. Chinese Journal of Applied &Environmental Biology, 2014, 20(5): 767 − 774. [22] NIJHAWAN A, JAIN M, TYAGI A K, et al. Genomic survey and gene expression analysis of the basic leucine zipper transcription factor family in rice [J]. Plant Physiology, 2008, 146(2): 323 − 324. [23] 于滔, 王成波, 曹士亮, 等. 玉米bZIP转录因子的生物信息学分析[J]. 黑龙江农业科学, 2016(4): 1 − 5. YU Tao, WANG Chengbo, CAO Shiliang, et al. Bioinformatics analysis of the bZIP transcription factor in maize [J]. Heilongjiang Agricultural Sciences, 2016(4): 1 − 5. [24] 王升级, 孙赫, 党慧. 盐胁迫条件下杨树bZIP转录因子全基因组分析[J]. 山西农业大学学报, 2018, 38(8): 1 − 7, 14. WANG Shengji, SUN He, DANG Hui. Genome-wide analysis of the bZIP transcription factors in Populus in response to salt stress [J]. Journal of Shanxi Agricultural University, 2018, 38(8): 1 − 7, 14. [25] HAN Xiao, ZHANG Junhong, HAN Shuang, et al. The chromosome-scale genome of Phoebe bournei reveals contrasting fates of terpene synthase (TPS)-a and TPS-b subfamilies [J/OL]. Plant Communications, 2022, 3(6): 100410[2023-05-20]. doi: 10.1016/j.xplc.2022.100410. [26] CHEN Chengjie, CHEN Hao, ZHANG Yi, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data [J]. Molecular Plant, 2020, 13(8): 1194 − 1202. [27] BALOGLU M C, ELDEM V, HAJYZADEH M, et al. Genome-wide analysis of the bZIP transcription factors in cucumber [J/OL]. PLoS One, 2014, 9(4): e96014[2023-05-20]. doi: 10.1371/journal.pone.0096014. [28] CORRÊA L G G, RIAÑO-PACHON D M, SCHRAGO C G, et al. The role of bZIP transcription factors in green plant evolution: adaptive features emerging from four founder genes [J/OL]. PLoS One, 2008, 3(8): e2944[2023-05-20]. doi: 10.1371/journal.pone.0002944. [29] ZHAO Kai, CHEN Song, YAO Wenjing, et al. Genome-wide analysis and expression profile of the bZIP gene family in poplar [J/OL]. BMC Plant Biology, 2021, 21(1): 122[2023-05-20]. doi: 10.1186/s12870-021-02879-w. [30] DUAN Lili, MO Zejun, FAN Yue, et al. Genome-wide identification and expression analysis of the bZIP transcription factor family genes in response to abiotic stress in Nicotiana tabacum L. [J/OL]. BMC Genomics, 2022, 23(1): 318[2023-05-20]. doi: 10.1186/s12864-022-08547-z. [31] HAN Xueyuan, MAO Linchun, LU Wenjing, et al. Positive regulation of the transcription of AchnKCS by a bZIP transcription factor in response to ABA-Stimulated suberization of kiwifruit [J]. Journal of Agricultural and Food Chemistry, 2019, 67(26): 7390 − 7398. [32] CHEA W L, WOONHEE B, SUNG C L. Roles of pepper bZIP protein CaDILZ1 and its interacting partner RING-type E3 ligase CaDSR1 in modulation of drought tolerance [J]. The Plant Journal, 2018, 96(2): 452 − 467. [33] 薛翀, 邱诗蕊, 李虹, 等. 烟草bZIP基因家族的鉴定及其A亚族在ABA处理下的表达分析 [J]. 分子植物育种, 2020, 18(17): 5607 − 5621. XUE Chong, QIU Shirui, LI Hong, et al. Identification of bZIP gene family and gene expression analysis of its subgroup A gene under ABA treatment in Nicotiana tabacum [J]. Molecular Plant Breeding, 2020, 18(17): 5607 − 5621. [34] 武立伟, 徐志超, 王清, 等. 响应ABA胁迫的甘草bZIP转录因子的系统筛选与分析 [J]. 药学学报, 2022, 57(3): 818 − 830. WU Liwei, XU Zhichao, WANG Qing, et al. Systematic screening and analysis of bZIP transcription factors in Glycyrrhiza uralensis and their response to ABA stress [J]. Acta Pharmaceutica Sinica, 2022, 57(3): 818 − 830. [35] 孙晓丽, 李勇, 才华, 等. 拟南芥bZIP1转录因子通过与ABRE元件结合调节ABA信号传导[J]. 作物学报, 2011, 37(4): 612 − 619. SUN Xiaoli, LI Yong, CAI Hua, et al. Arabidopsis bZIP1 transcription factor binding to the ABRE cis-element regulates abscisic acid signal transduction [J]. Acta Agronomica Sinica, 2011, 37(4): 612 − 619. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230342
 点击查看大图
点击查看大图

计量
- 文章访问数: 2099
- HTML全文浏览量: 670
- PDF下载量: 45
- 被引次数: 0
